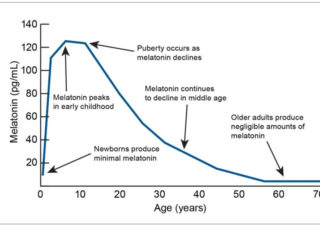
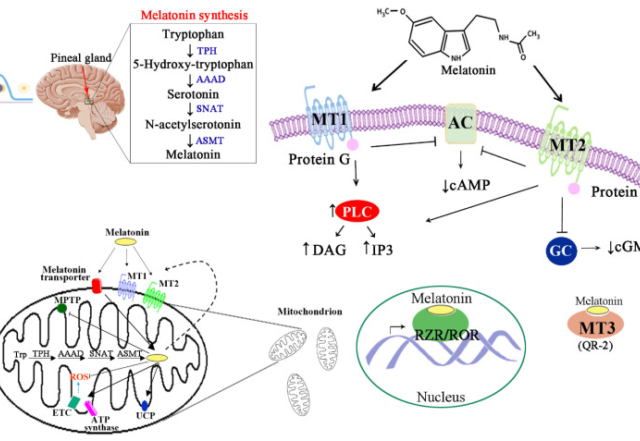
メラトニン 成人におけるメラトニンの高用量の安全性:系統的レビューとメタアナリシス Safety of higher doses of melatonin in adults: A systematic review and meta-analysis 初公開: 2021年12月19日 資金調達情報 CMHとALDはNHMRC-ARC Dementia Rese 2024/05/23 メラトニン
メラトニン 書籍:『メラトニン:アルツハイマー病などに対する守護神!』 第1版 2017Melatonin MELATONIN ACARIST ALZHEIMER'S AND MORE! 文:クリストファー・ヘルツォーク博士 序文 体内の重要なホルモンであるメラトニンは、過小評価され、誤解されている。 ほとんどの人がメラトニンのことを耳にしたことがあると思うが、一般的には睡眠パターンの 2024/05/23 メラトニン
ステロイドホルモン 書籍:超高用量メラトニン 奇跡のアンチエイジングホルモン抗アルツハイマー病ホルモン・抗加齢ホルモン・更年期障害逆転ホルモン Extreme Dose! Melatonin The Miracle Anti-Aging Hormone Anti-Alzheimer’s Hormone Anti-Baldness Hormone Menopause Reversal Hormone ジェフ・T・ボウルズ 2024/05/22 ステロイドホルモンメラトニン認知症 予防と治療
メラトニン メラトニンで治療されたアルツハイマー病の一卵性双生児:症例報告 Monozygotic twins with Alzheimer's disease treated with melatonin: Case report 松果体研究 1998: 25:260-263 ブルスコLI、マルケスM、カルディナリDP。メラトニンで治療されたアルツハイ 2024/05/21 メラトニン
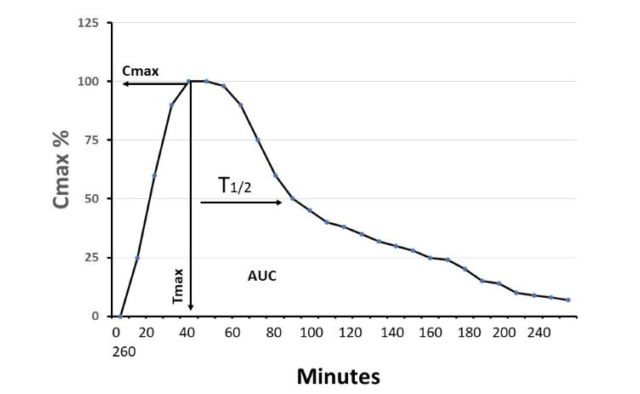
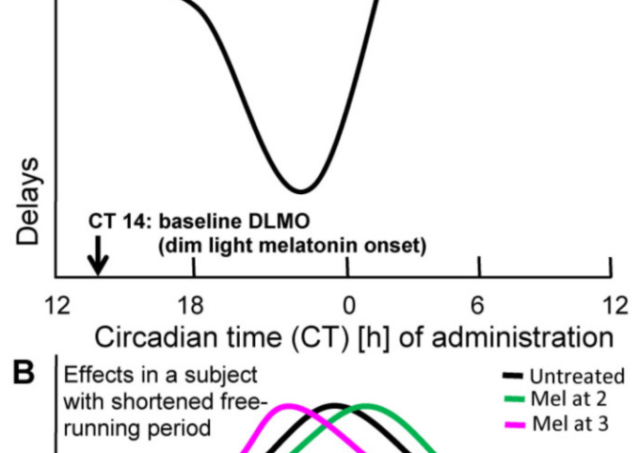
メラトニン 高用量メラトニン療法と低用量メラトニン療法における時間生物学的考察の重要性は異なる Divergent Importance of Chronobiological Considerations in Highand Low-dose Melatonin Therapies リュディガー・ハードランド Johann Friedrich Blumenbach In 2024/05/21 メラトニン
メラトニン メラトニンは放射線腫瘍治療に役立つか? Can Melatonin Help Us in Radiation Oncology Treatments? 総説|オープンアクセス 2014年|論文ID 578137 受領 2014年2月24日 受理 2014年4月15日 掲載 2014年5月11日 要旨 今日、放射線治療は 2024/05/20 メラトニン癌・ガン・がん
メラトニン 書籍:『メラトニン奇跡の分子』(2022) Melatonin Miracle Molecule: Transform your life with Melatonin. Why higher doses are safe and benefits beyond sleep as the bodies master str 2024/04/20 メラトニンワクチン後遺症治療生物毒素・カビ毒・3型睡眠神経発達障害(自閉症・ADHD)電磁波・5G・6G
メラトニン メラトニンの高用量についての衝撃的な真実メラトニンサプリメントは、あなた自身の生産をシャットダウンするか、断食を強化するためにメラトニンを使用する方法 – The Shocking Truth About High-Dose Melatonin, Does Melatonin Supplementation Shut Down Your Own Production, How To Use Melatonin To Enhanc 2024/04/11 メラトニン
メラトニン ライフエクステンション:高用量メラトニンの症例新たなデータによると、メラトニンはがんを予防し、免疫機能を向上させ、炎症を抑える働きがあるという。 A Case for Higher-Dose Melatonin 科学的レビュー 2023年8月、医学博士ゲイリー・ゴンザレス著:ウィリアム・ファルーン ウィリアム・ファルーン 2022年2月1日、米国医師会雑誌(JAMA)は、米国におけるメラトニンの消費量が1999年から201 2024/03/14 メラトニン
メラトニン 書籍:『メラトニンのマジック』 The Magic of Melatonin 目次 序文 はじめに I. メラトニン研究の基礎知識 時空間生物学 メラトニンとその作用機序 メラトニンとその主な応用分野 健康的なメラトニン生成の乱れ II. メラトニンの実際 多才なメラトニン 様々な病態におけるメラトニン 睡眠 2023/06/07 メラトニン癌・ガン・がん
メラトニン がん治療におけるメラトニン:現在の知見と将来の可能性Melatonin in Cancer Treatment: Current Knowledge and Future Opportunities 2021年4月25日オンライン公開 doi:10.3390/molecules26092506 Melatonin in Cancer Treatment: Current Knowledge and Future Opportunities pmcid: pmc8123278 2023/04/25 メラトニン癌・ガン・がん
マシュー・クロフォード 太陽と暗闇のホルモン「メラトニン」が健康を守り、命を救うMelatonin - the Hormone of Both Sun and Darkness - Protects Your Health and Can Save Your Life Melatonin - the Hormone of Both Sun and Darkness - Protects Your Health and Can Save Your Life この記事は、パンデミック時に私の非常に生産的なペンフレンドの一人、M. Langenが書い 2023/03/05 マシュー・クロフォードメラトニン
メラトニン メラトニン 治療効果と神経保護MELATONIN Therapeutic Value and Neuroprotection MELATONIN Therapeutic Value and Neuroprotection 目次 序文 謝辞 編集者 貢献者 第1章 メラトニンの生成とバイオアベイラビリティ 第2章 加齢と疾患におけるメラトニンレベルとシグナリングの逸脱。 治療の選択肢と限界 第3章 松果体 2023/02/13 メラトニン睡眠
ビタミンD・紫外線・日光浴(総合) メラトニンは「次のビタミンD」なのか?新たな科学、臨床的使用、安全性、栄養補助食品についてのレビューIs Melatonin the “Next Vitamin D”?: A Review of Emerging Science, Clinical Uses, Safety, and Dietary Supplements ニュートリエント(Nutrients)2022 Oct; 14(19):3934. 2022年9月22日オンライン公開 doi:10.3390/nu14193934 pmcid: pmc9571539 PMID:36235587 Deanna M. Minich,1,*Melan 2022/12/22 ビタミンD・紫外線・日光浴(総合)メラトニン神経発達障害(自閉症・ADHD)
メラトニン メラトニンは21世紀の宝(コルヌコピア)か?Is Melatonin the Cornucopia of the 21st Century? オンラインでは2020年11月5日に公開 概要 メラトニンは、夜間に松果体や松果体外細胞で産生・分泌されるインドールアミン系ホルモンであり、ヒトの概日リズム(24時間体内時計)の計時や睡眠・覚醒サイクルの調整に重要な役割を果たしている。 しかし、近年、メラトニンが注目されているの 2022/01/30 メラトニン癌・ガン・がん睡眠神経発達障害(自閉症・ADHD)
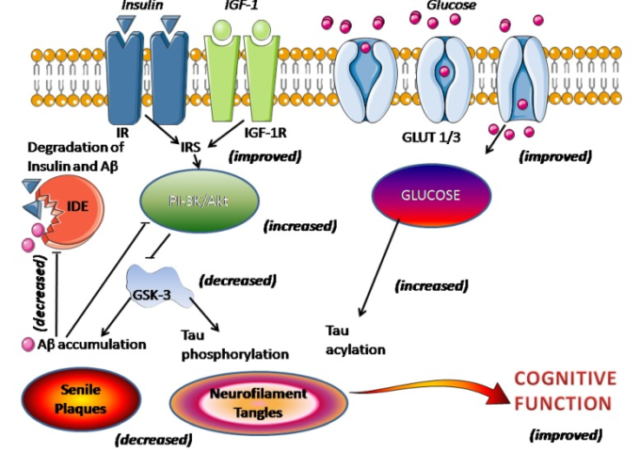
メラトニン アルツハイマー型認知症患者におけるメラトニン療法について Melatonin Therapy in Patients with Alzheimer’s Disease オンライン 2014 Apr 10 概要 アルツハイマー病(AD)は大きな健康問題であり、その予防のための努力を政府機関と非政府機関の両方で行わなければならないという認識 2022/01/30 メラトニン
メラトニン 神経変性疾患と高用量メラトニン(メガドース) 高用量投与に関する文献 メラトニンは、筋萎縮性側索硬化症の疾患進行を遅らせる可能性がある 主要生存分析では、メラトニン使用者は非使用者に比べて年間死亡率が有意に低かった(ハザード比,0.241;95%信頼区間,0.088-0.659;P = 0.0056)(図2A)。研究期間中に 2022/01/19 メラトニン神経変性疾患 精神疾患
メラトニン 致死性ウイルス感染症治療のためのメラトニンの推定投与量:COVID-19に焦点を当てて Estimated doses of melatonin for treating deadly virus infections: focus on COVID-19 Received: 2020年4月19日; Accepted: 受理:2020年4月19日、受理:2020年6 2022/01/10 メラトニン
筋萎縮性側索硬化症(ALS) メラトニンは、筋萎縮性側索硬化症の疾患進行を遅らせる可能性がある Melatonin may slow disease progression in amyotrophic lateral sclerosis: Findings from the Pooled Resource Open-Access ALS Clinic Trials dat 2022/01/10 筋萎縮性側索硬化症(ALS)
メラトニン メラトニン高用量療法と低用量療法における時間生物学的考察の重要性の違いDivergent Importance of Chronobiological Considerations in High- and Low-dose Melatonin Therapies オンラインで2021年3月9日に公開 リューディガー・ハードランド(Rüdiger Hardeland) アレハンドロ・ロメロ・マルティネス(学術編集者 概要 メラトニンは、様々な目的で臨床前および臨床で使用されている。その中には、概日リズムの再調整に関連するものもあれば、メラト 2021/12/03 メラトニン