Contents
アルツハイマー病と鉄 用語
関連記事
参考までに正確に知りたい方は他の専門サイトをあたってください。
鉄代謝関連用語(覚書)
二価鉄
不安定で毒性が強い。
細胞膜の通過、細胞内での酸化、還元反応などの触媒で一時的に出現する。
そのため危険といっても、鉄代謝には必須
二価鉄と三価鉄は平衡状態にある。(三価鉄が増えると二価鉄も増える)
二価鉄濃度の上昇によって神経変性に関与するタンパク質(αシヌクレイン、過剰リン酸化tau)などの凝集を引き起こすことがin vitro研究で示されている。
www.ncbi.nlm.nih.gov/pubmed/20502498/
www.ncbi.nlm.nih.gov/pubmed/12358761/
三価鉄
生体内の鉄は、ほとんどが安定した三価鉄として存在している。
体内で移動する時には運び屋が必要。
トランスフェリンと強力に結合する。
ヘム鉄
二価の鉄原子とポルフィリン
ヘムタンパクに囲われた鉄のためヘム鉄という。
赤肉、魚など含まれる、非ヘム鉄と比べ吸収力5倍、
非ヘム鉄
野菜に含まれる三価鉄、吸収には三価鉄から二価鉄へ還元する必要がある。ビタミンC、酢、胃液など。
トランスフェリン (Tf)
鉄の運び屋
血中で3価の鉄と結合して輸送を担うタンパク質。
体内の鉄のほとんどはトランスフェリンと結合しており、いわゆる血清鉄はこのトランスフェリンと鉄の複合体をさす。
脳への輸送
脳へも、トランスフェリン受容体を介したエンドサイトーシスによって血液脳関門を透過して輸送される。
その後脳脊髄液に吸収され循環系に戻るが、脳の鉄必要量は血液中の鉄よりも高いため、脳内で鉄リサイクルされていると考えられている。
そのため、体内鉄の過剰または欠乏が脳内鉄に影響を及ぼすことは少なく、鉄過剰症であっても脳への異常鉄沈着は認められない。
トランスフェリン受容体1(TfRA1)
鉄の受け取り
トランスフェリンを細胞内へ取り込む受容体
![]()
セルロプラスミン(Cp)/フェロオキシダーゼ
銅の運び屋
肝臓で産生されるタンパク質で銅の運搬と代謝、鉄の代謝(二価鉄→三価)に関与している。血清銅はセルロプラスミンと結合している。
セルロプラスミン(Cp)は、血中に分泌される「分泌型Cp」と細胞膜に結合する「GPI結合型Cp」の2種類のアイソフォームが存在する。
二価鉄を三価鉄に
二価の生体不安定鉄はフェロオキシダーゼによって酸化活性され、トランスフェリンに移行する。
「GPI結合型Cp」(GPI-Cp)
「GPI結合型Cp」がないと、鉄の動員に必要なフェロオキシダーゼ活性がなくなり、細胞膜のフェルロポルチンが分解され、鉄の細胞外への排出が阻害されるため細胞内に鉄が蓄積する。
セルロプラスミンがないと脳の鉄蓄積にも寄与し神経変性をもたらす。
www.ncbi.nlm.nih.gov/pubmed/12743117/
ヘファスチン (Hephaestin)
セルロプラスミンの仲間
セルロプラスミン同様三価鉄に酸化させる。フェロポーチン1と協力して鉄排出を媒介する。
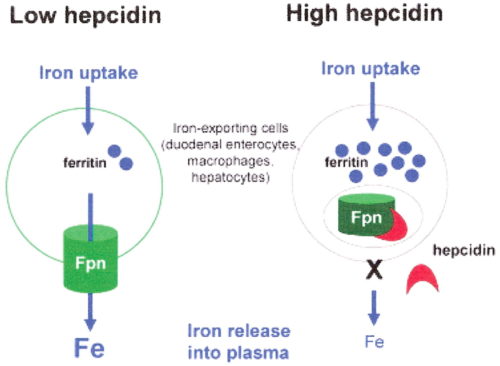
ヘプシジン
鉄の分布を調節する因子
ヘプシジンは、全身の鉄の恒常性を調節するタンパク質。
IL-6によって肝臓で合成されるホルモン、マクロファージからの鉄排出を抑制、腸管からの鉄吸収を抑制、炎症 → IL-6 → ヘプシジン産生 → 赤血球減少 → 貧血
ヘプシジンの分泌が亢進すると、ヘプシジンがフェロポーチンを分解に導く(フェロポーチンはヘプシジン受容体でもある)。
ヘプシジンは体内の鉄量を需要に応じて調整する因子であり、複数の要因によって増加する。
炎症性サイトカイン、鉄濃度の飽和、造血、低酸素等
テストステロンはヘプシジンの転写を阻害し、赤血球への鉄の取り込みを増加させる。
www.ncbi.nlm.nih.gov/pubmed/23399021/
エストロゲンは、肝臓ヘプシジンを制御することにより、鉄恒常性を調節する。エストロゲン欠乏では、過剰な組織鉄を伴う閉経後骨粗鬆症と関連する可能性がある。
www.ncbi.nlm.nih.gov/pubmed/23041085/
ヘプシジンを阻害することで、細胞内の鉄排出を促すことができる。
フェロポーチン(Fp)
鉄の排出を促進(細胞から)
血管上皮に存在し、細胞外へ鉄を排出する役割をもつ。ヘプシジンはこのフェロポーチンに結合することで鉄の放出を抑制する。
鉄を細胞外へ輸送するためのタンパク質、APPはこのフェロポーチンを細胞表面上に固定するのに役立つ。βアミロイドペプチドが細胞表面上のAPPの量を減少させ、フェロポーチンの細胞表面上への固定能力も低下させる。→ 細胞内の鉄が上昇 → フリーラジカルにより細胞損傷が亢進(アルツハイマー病患者に見られるニューロン損傷)
フェロポーチンは鉄に加えて、コバルト 、亜鉛、ニッケル、を排出することが示されている。
www.ncbi.nlm.nih.gov/pubmed/24304836
www.ncbi.nlm.nih.gov/pmc/articles/PMC6079014/
フェロポーチンはマンガンを排出する可能性もある。
www.ncbi.nlm.nih.gov/pmc/articles/PMC5695046/
APPのフェロポーチン1活性
アミロイドβ前駆体タンパク質(APP)はニューロンの細胞膜にある鉄イオンを排出するフェロポーチン1と結合し鉄排出を促進する。
www.ncbi.nlm.nih.gov/pubmed/25464026/
www.ncbi.nlm.nih.gov/pubmed/20817278/
細胞の鉄レベルが枯渇すると、鉄代謝関連の遺伝子発現を調節することによって細胞鉄の恒常性を維持する2つ鉄調節タンパク質1,2(IRP1、IRP2)が活性化される。IRPは鉄欠乏の条件下で鉄の生物学的利用能を高めるように作用する。
(フェロポーチン1と相互作用する)APPの形質膜への輸送は、タウタンパク質の活性に依存する。
www.ncbi.nlm.nih.gov/pubmed/22286308/
鉄はタウタンパク質の過剰リン酸化を誘発し、リン酸化タウの凝集を促進するようであり、
www.ncbi.nlm.nih.gov/pubmed/23262393/
アルツハイマー病では過剰リン酸化タウの蓄積がすでにあることから、神経細胞膜での利用可能なAPPが減少し、ニューロンからの鉄の排出が減少することが予想される。
鉄の蓄積は、初期の発達中の脳では重要であり、初期の脳において不十分な鉄蓄積は、その後鉄補給による鉄の異常な基準値を決定する可能性がある。
www.frontiersin.org/articles/10.3389/fnagi.2013.00034/full
亜鉛による阻害
アミロイドβ前駆体タンパク質(APP)による、フェロキダーゼ(セルロプラスミン)活性は、内因性の亜鉛によって阻害される。この亜鉛はアミロイド凝集体からの亜鉛に由来し、アミロイドβの負荷と相関する。
この結果は、フェロポーチンを亜鉛が阻害することで細胞内の鉄蓄積を促進する可能性がある。
カルシウム
フェロポーチンを介した鉄の排出はカルシウムで活性化される。
www.ncbi.nlm.nih.gov/pubmed/30082682
ヘムオキシゲナーゼ1(HO-1)
遊離ヘム除去係
ヘム分解の律速酵素、ヘムタンパクから遊離した遊離ヘムを除去することで酸化ストレスに対抗する。代謝産物も細胞保護的に機能する。
しかし、グリア細胞でのHO-1のアップレギュレーションは神経変性に寄与する可能性がある。
www.ncbi.nlm.nih.gov/pubmed/19457088/
アルツハイマー病マウスではHO-1の過剰発現が脳内の鉄負荷、タウ凝集につながる。
www.ncbi.nlm.nih.gov/pubmed/21613741/
アルツハイマー病、軽度認知障害患者の海馬、側頭皮質ではHO-1がアップレギュレートされており、アルツハイマー病の病因と関連することが示唆されている。
www.ncbi.nlm.nih.gov/pubmed/7778849/
www.ncbi.nlm.nih.gov/pubmed/16399210/
鉄トランスポーター1(DMT1)
二価鉄の受取屋
非ヘム鉄と、二価イオン(Zn、Mn、Cu、Co)を細胞内へ取り込むトランスポーター、腸管からの鉄吸収をコントロールする。脳にも発現していて、パーキンソン病の黒質鉄過剰とも関係している。
ヘムキャリアープロテイン(HCP-1)
ヘム鉄の受取屋
ヘム鉄を上皮細胞内に取り入れるトランスポーター、葉酸の輸送体でもある!?
不安定鉄プール Labile Iron Pool(LIP)
細胞内で使われるフリーの二価鉄の分画
DNA合成と修復、細胞周期、ミトコンドリア、ヘム合成、鉄貯蔵、などさまざまな代謝プロセルに不可欠であり、キレート化が可能な酸化鉄、還元鉄を貯蔵する鉄代謝の交差点。
しかし、フェントン反応の触媒としても作用し、酸化ストレス、活性酸素種(ROS)の放出を引き起こすこともある。
細胞のLIPの多くはサイトゾルに存在するが、サイトゾルだけでなく、ミトコンドリア、リソソーム、核にも存在する不均一な全体のプールを表す。
細胞内LIP
鉄がLIPとして細胞内で隔離されることは、活性酸素種のフェントン反応の触媒を防ぐことができるため理にかなう。
細胞内LIPの変化は、ガン、心血管疾患、神経変性障害などさまざまな疾患の開発に深く関与することが示されている。
鉄の貯蔵形態
サイトゾル内のLIPは、主にグルタチオンとFe2+の結合によって存在する。
ミトコンドリア内では、鉄はプロトポルフィリンIX(PPIX)に組み込まれている。
リソソーム内では、金属タンパク質および鉄含有オルガネラの分解によりキレート化可能な鉄が含まれている。
鉄依存性およびOH·媒介性のDNAへの酸化的損傷が報告されており、核内の鉄は主にリソソームの破裂に由来している。
フェリチン
フェリチンの過剰発現はサイトゾルの不安定鉄を減らし、細胞を酸化ストレスから保護する。サイトゾルのフェリチンは、不安定な鉄の解毒と反応性ラジカル生成の防止に非常に効果的。
反対に、フェリチンのダウンレギュレーションはLIPと酸化ストレスを増加させる。
フェリチン中の鉄は、酸性リソソームで利用可能な形に動員され、細胞が鉄を必要とする条件下で代謝利用にリサイクルできる。
ミトコンドリア
ミトコンドリアは、ヘム鉄が鉄・硫黄クラスター生合成経路を介して不安定鉄、生物活性型に変換される主要な目的地。細胞内鉄を使用する主要な場所であり、鉄の輸送、貯蔵、鉄調節タンパク質に依存して鉄の恒常性を維持する。
ISCの合成および機能の欠陥によるミトコンドリアLIPの過剰なレベルは、フリードライヒ運動失調、パーキンソン病、アルツハイマー病などの神経変性疾患を引き起こす可能性があるという仮説が立てられている。
赤血球細胞では、ほとんどのミトコンドリア鉄がさらにヘムおよびFe-Sクラスター上に集合し、過剰な鉄はミトコンドリアに特異的に局在する鉄隔離タンパク質であるミトコンドリアフェリチン(FtMt)に保存される。
FtMtの過剰発現は、神経芽細胞腫細胞でのH2O2誘発細胞死を防止する。
ミトコンドリアは、主に3つの方法で鉄を獲得する。
サイトゾルLIPからの鉄の直接取り込み
エンドソームはミトコンドリアと直接接触し、それにより鉄がミトコンドリアに移動する
DMT1はミトコンドリア外膜にも見られ、DMT1はミトコンドリアへの鉄の輸入に効果を発揮する。
フラタキシン
フラタキシンは、真核細胞のミトコンドリアで発現しており、鉄・硫黄クラスターの組み立てに関与するミトコンドリアの鉄シャペロン。
フラタキシン増強剤 ALA 5-アミノレブリン酸(黒酢、赤ワイン、タコ)
サイトゾルの鉄代謝は、ヘムと鉄硫黄クラスターを合成するための鉄のミトコンドリア需要によって部分的に調節されている。
ヘム生成の欠如は、トランスフェリン(Tf)結合鉄摂取の代償的増加をもたらすようである。
鉄・硫黄クラスターとヘム合成の欠陥は、ミトコンドリアの鉄負荷状態から相対的な細胞質鉄欠乏への移行をもたらすようである。
pubs.rsc.org/en/content/articlelanding/2018/MT/C8MT00048D#!divAbstract
フェリチン(FPN)
鉄の貯蔵庫
トランスフェリンによって運ばれた鉄を細胞内に貯蔵する鉄の腫瘍な貯蔵庫。
反応性の高い二価鉄イオンを酸化させて三価鉄イオンとして無毒化する。
4500分子を貯蔵庫に入れておくことが可能で、過剰鉄によって組織が障害されないように役割ももつ。
フェリチンのサブユニットにはL鎖とH鎖があり、L鎖に遺伝子異常があると、鉄貯蔵機能が低下し、グリア細胞、アストロサイトに鉄が蓄積しジストニア、不随意運動障害、認知機能障害に寄与する。
重鎖/軽鎖比(H鎖/L鎖)調節不全
アルツハイマー病患者では、H鎖とL鎖の比率が健常高齢者と比べて5倍高く、H/Lの不均衡、調節不全がアルツハイマー病における炎症、活性酸素ラジカルの産生と関与している可能性がある。
www.ncbi.nlm.nih.gov/pubmed/7616228
L鎖とミトコンドリアフェリチン
www.ncbi.nlm.nih.gov/pubmed/20176086
黒質ではフェリチン重鎖/軽鎖濃度は加齢とともに直線的に増加するが青斑核では一定。そのため青斑核よりも黒質での鉄が神経変性に寄与しえる。
炎症
体に炎症があると細胞内のフェリチンは増加する。
www.ncbi.nlm.nih.gov/pubmed/8695634
リソソームでのフェリチン分解障害
酸化ストレスは、リソソーム機能を調節する重要な因子。
例えば、フェリチンなどの金属タンパク質を分解することなどにより、リソソームに高レベルの鉄が含まれると、三価鉄から反応性の高いヒドロキシルラジカルが生成される。
リソソーム膜は、フェントン反応で二次的に形成されるROSによって容易に過酸化および透過化される。
ヒドロキシルラジカルは、タンパク質、脂質、およびそれらの分解を阻害する他の高分子を酸化することで、リソソームでは消化できない廃棄物を生成し蓄積されてしまう。
リソソーム廃棄物は、リソソーム酵素活性を損ない、リソソーム膜透過性の亢進(LMP)を促進することが知られている。
www.ncbi.nlm.nih.gov/pubmed/18764739
フェリチノファジー
フェリチノファジー(ferritinophagy)細胞内のフェリチンに結合した三価鉄を遊離二価鉄に変換するには,フェリチンに特異的なオートファジーによるフェリチンの分解が必要であり,これをフェリチノファジーとよぶ。
NCOA4を介したフェリチノファジーの調節不全は、全身性鉄の恒常性を破壊し、赤血球生成と酸化ストレスの調節に有害な影響を及ぼす。
最近の研究では、NCOA4を介したフェリチノファジーがフェロプトーシスに対する感受性を調節することが示されている。
www.frontiersin.org/articles/10.3389/fnins.2019.00238/full
NCOA4と相互作用する因子
- アンドロゲン受容体
- ペルオキシソーム増殖因子活性化受容体ガンマ
- フェリチン
- ATG8
ヘモジデリン hemosiderin
人体には過剰な鉄を排出する機能がないため、鉄の毒性を最小限に抑えるためにフェリチンとヘモジデリンの形で貯蔵される。ヘモジデリンはフェリチンの異常な代謝経路からも生成される。
ヘモジデリンは血液中を循環しておらず、細胞内でのみで見られる。
ヘモジデリンは、フェリチン、変性フェリチン、その他の物質の代謝産物のフェリチン複合体であると考えられており、食細胞のヘマチン消化によって生成される。
ヘモジデリンに沈着する鉄は、必要な時に十分に利用できないことを特徴とする。
ヘモジデリン鉄当たりのフェリチンの比率は、総鉄貯蔵量の変化に従って変化する。
通常の貯蔵鉄の範囲では、フェリチン鉄の比率がヘモジデリン鉄の比率よりも大きいが、鉄過剰によりフェリチン鉄の増加により飽和に近づくと、比率は逆転する。
www.ncbi.nlm.nih.gov/pmc/articles/PMC4664589/
www.ncbi.nlm.nih.gov/pmc/articles/PMC3770782/
IRP1・IRP2(鉄応答性要素)
フェリチンの発現調整を通して、鉄代謝をコントロールするRNA結合タンパク質
IRP2が細胞の鉄恒常性において重要な働きを示す。
生体不安定鉄(非トランスフェリン結合鉄 NTBI)
主にトランスフェリンとの結合能を超えてしまい飽和したフリー鉄をさす。(本当に危険な鉄はオキソ架橋二核鉄(III)かもしれない。)
フリーラジカルが発生し、細胞小器官の機能障害、最終的には細胞傷害、DNA障害、臓器線維化、発癌などにいたる。

フェロトーシス(フェロプトーシス)
細胞内鉄がもたらす特定の細胞死、アポトーシス、オートファジーとは異なる細胞死経路。
最近の研究の証拠では、このフェロプトーシスが、酸化還元活性鉄、脂質過酸化ストレスによって脳神経細胞で生じアルツハイマー病に寄与する可能が示されている。
www.sciencedirect.com/science/article/pii/S0166432817316960?via%3Dihub
www.ncbi.nlm.nih.gov/pubmed/22632970/
フェロトキシンは、エラスチン、スルファラジン、RSL3などの多様な小分子によって誘発され、トロロックス(水溶性のVEアナログ)やビタミンEなどの親油性抗酸化物質やデフェロキサミンなどの鉄キレート剤によって阻害することができ、アルツハイマー病の治療に期待できる。
フーリンタンパク質(furin)
アミロイド前駆体タンパク質APPの切断に関わるαセクレターゼおよびβセクレターゼはフーリンタンパク質によって調節される。
www.ncbi.nlm.nih.gov/pubmed/23390091/
フーリンの転写は細胞濃度の鉄によって調節される。
細胞内の鉄濃度が高いとフーリン活性は低下し、βセクレターゼ活性が促進され、アミロイドβ形成経路が増強される。
鉄欠乏は逆にフーリン活性を増加させ、αセクレターゼを増強し非アミロイド形成経路刺激する。
www.ncbi.nlm.nih.gov/pubmed/19387120/

www.ncbi.nlm.nih.gov/pubmed/18466351/
