
Did human hairlessness allow natural photobiomodulation 2 million years ago and enable photobiomodulation therapy today? This can explain the rapid expansion of our genus’s brain
pubmed.ncbi.nlm.nih.gov/25703782/

要旨
人間の無毛を説明する現在の仮説は、無毛に即効性が伴わないため、不十分であるように思われる。
低レベル光療法としても知られる光生物調節の研究に基づいて、人間の無毛を説明する新しい、検証可能な仮説が提唱されている。これは、赤色および近赤外放射が、脳を含む表在組織に非常に有益な効果をもたらすことを示している。完全な無毛をもたらすランダム突然変異によって、初期の人類は日没時に赤色と近赤外放射線を毎日浴びることができた。
光生物調節の研究によると、これには2つの効果がある。 脳を含むすべての表在組織において、ミトコンドリア呼吸鎖活性が高まり、その結果ATPの「外部合成」が促進される。 また、転写因子NF-κBの活性化を通じて100以上の遺伝子の発現に有利な影響を与え、その結果、脳の代謝と血行動態が促進される。 また、メラニンが呼吸鎖に電子を供給し、ATPの外部合成をもたらす可能性もある。
このような作用は、無毛になると同時に自動的に始まり、その結果、関係する変異が選択的に掃討されることになる。これに続いて、過去200万年の間に非常に急速な脳の進化が起こったが、これは、新しく無毛になった個体のエネルギーと賢さの増加に基づく、知性主導の進化によるものだと考えられる。
ヒトの無毛は、200万年前に自然な光生物調節(特定の波長の生物組織への影響)を可能にし、今日の光生物調節治療を可能にしたのだろうか?このことは、ヒト属の脳が急速に拡大したことを説明することができる。t.co/xxDfBIs0ps
— Alzhacker (@Alzhacker) September 20, 2023
AI解説
この論文では、人間の無毛化が約200万年前に起こり、それによって自然な光生物調節(PBM)が可能になったことが、ホモ属の脳の急速な拡大を説明できるという仮説が提唱されている。
主な点は以下の通り:
- 1. 無毛化により、赤色と近赤外放射(R&NIRR)が体表に届くようになり、細胞のミトコンドリアでATPの外部合成が促進された。
- 2. R&NIRRは頭蓋骨を透過して脳に達し、多数の遺伝子発現を変化させ、大脳の代謝と血流動態を向上させた。
- 3. これらの効果は、無毛化とほぼ同時に生じたため、無毛の突然変異が急速に選択的に固定された。
- 4. その結果、過去200万年間にホモ属の脳が急速に拡大した。これは、無毛化によってもたらされたエネルギーと知性の向上に基づく、知性主導の進化によるものだと考えられる。
- 5. 日光とメラニンの相互作用により、表皮細胞でもATP合成が起こる可能性がある。
- 6. 脳の拡大は、食事の質とエネルギー密度の向上によって支えられた。
- 7. 北方の寒冷地では、無毛のデメリットが大きくなり、頭髪が再び発達し、衣服の着用が必要になった。
この仮説は、ヒトの脳の進化における無毛化の重要性を示唆するとともに、特に神経疾患に対する非侵襲的で効果的な治療法としてのPBMの可能性を示唆している。
この仮説を支持する証拠
- 1. PBMの研究により、赤色と近赤外放射(R&NIRR)が、試験管内および生体内でATPの外部合成を促進することが示されている。
- 2. PBMの研究により、R&NIRRが多数の遺伝子の発現を変化させ、大脳の代謝と血行動態を促進することが示されている。
- 3. 過去650万年間の脳容積の増加率は一定ではなく、200万年前頃から大きな継続的増加が始まっている(図1)。
- 4. この時期には、食事の質とエネルギー密度の向上が伴っている。
- 5. 真菌の一種であるクリプトコッカス・ネオフォルマンスのメラニンは、ガンマ線照射後に電子伝達と代謝を増加させることが示されており、ヒトのメラニンでも同様の効果がある可能性がある。
- 6. MCPH1、ASPM、FOXP2など、脳の大きさや認知機能に関連する遺伝子の適応的進化が、過去200万年の間に起こったことが示唆されている。
- 7. 動物実験において、経頭蓋PBMがATPレベルの回復、シトクロムcオキシダーゼ活性の上昇、脳血流の増加などの効果を示している。
- 8. ヒトを対象とした研究では、経頭蓋PBMが認知機能の改善や、うつ病などの神経疾患の治療に有効であることが示されている。
はじめに
人間の無毛を説明しようとする現在の仮説[1]、[2]は不十分であるように思われる。無毛の人に直接的な利益を与えるわけでもなく、過去2世紀の間に爆発的に増加した人間の脳の容積を説明するものでもない。無毛の問題は、チャールズ・ダーウィンの時代から問題になっていた[1]。
ワクチンロンスキー [2], [3]によって提唱された現在の有力な説は、ヒトの系統は1.6mya(160万年前)までに無毛になり、無毛によって熱ストレス時の蒸発冷却を可能にする汗腺がさらに発達したことを示唆している。ワクチンロンスキーは、体毛が減少するにつれて汗腺の密度が増加するような、漸進的なプロセスとして無毛化が起こったと想定している。[4]。これはおそらく、無毛化が進むにつれて暑熱ストレスを緩和する漸進的な適応と見ることができるだろう。
JablonskiとChapman [3], [4]は、紫外線(UVR)によるビタミン葉酸の光分解の証拠も指摘している。UVRは熱帯地方や高地でより高くなる。南緯27°に位置するオーストラリアのブリスベンで最近行われた調査では、出産適齢期の女性において、UVR光分解による葉酸レベルの有意な、最大20%までの減少が確認された。[5]。これは、オスロ(北緯60°)で得られた逆の所見[6]にもかかわらず、おそらくUVRの緯度差が一役買っている。妊娠中の葉酸の欠乏は、出生時の神経管欠損症や無脳症、その後の人生における巨赤芽球性貧血に関連しており、先進国では食品の葉酸強化につながっている。
アフリカで無毛が発生したとき、ワクチンロンスキーは素肌がピンク色であったと仮定している。[2]。もしそうであれば、すぐに色素が濃くなることで保護されることになるが、これは紫外線Aを介した皮膚のメラノサイトにおける視覚光色素ロドプシンの発現によるもので、その結果メラニンが急速に生成されることが示されている。[7]。その後、遅発性日焼け反応が起こる。[4]。アフリカ人の象徴的な黒褐色の肌は、MC1R遺伝子座の突然変異によって定められ、今日に至っている。しかし、アフリカ人と比較してユーラシア人のMC1R遺伝子座における非同義変異の多さは、十分なビタミンD合成を維持するために、ユーラシア人の肌がより淡くなるように適応進化した証拠と考えられている。[8]。

ヒトの無毛を説明する別の解決策を探すと、ビタミンDが関係しているのではないかと考えるかもしれない。ビタミンDの合成は、297nmのUVBが皮膚の7-デヒドロコレステロールと相互作用するときに始まる。ビタミンD受容体(VDR)はリガンド活性化転写因子で、脳、皮膚、その他の組織に広く存在する。ビタミンD合成の最終産物であるカルシトリオールはVDRに結合し、それによって特に脳における何百もの他の遺伝子の発現に影響を与える。[9], [10]。ビタミンD欠乏症はくる病の原因として古くから知られているが、欠乏症は、近位筋の痛みを伴う筋力低下 [11]や、出産時に帝王切開を必要とする確率が4倍近く高くなる。[12]など、古典的な症状以外の影響も引き起こす。ビタミンDは自然免疫を抑制する傾向があり、早期の欠乏は後の自己免疫疾患と関連する可能性がある。[13]。
カルシトリオールは、発達中のラットの脳において神経新生を刺激し、細胞内カルシウムの制御を通じて髄鞘形成、シナプス形成および神経伝達物質放出を促進する。[14] ;VDRは、ミエリン形成に関与するオリゴデンドロサイトおよびシュワン細胞において同定されている。ニューロトロフィンの分泌促進および細胞内カルシウムの制御を通して、カルシトリオールは、脳の原始的な領域だけでなく、大脳新皮質の発達および機能にも影響を及ぼしている。[14].
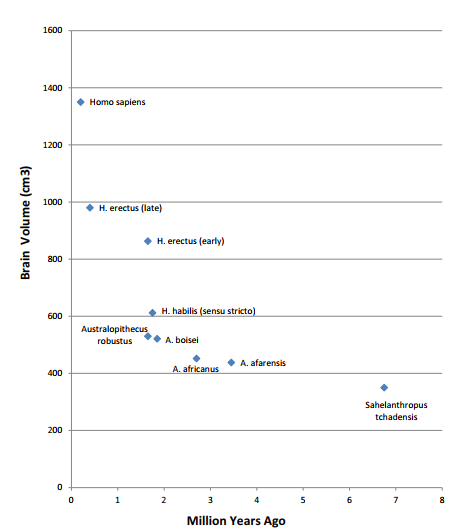
図1. Leonardら[61]の表2のデータを、様々な種の地質年代の中点を用い、サヘラントロプスを加えてグラフ化したもの。W. Leonard教授に謝意を表する。
2つの研究 [15], [16] では、ビタミンDの低値と高齢者の認知機能低下との間に相関関係があることが明らかにされているが、最近の大規模な研究では、その相関関係があらゆる形態の認知症にまで拡大されている。[17]。統合失調症は、肌の色が黒く高緯度への移住者や都市環境において有意に多い。冬と春に生まれた乳児は、後に統合失調症になる傾向が有意に高いが、これはおそらく妊娠後期のビタミンDレベルの低さ、あるいはインフルエンザなどの母親の冬期のウイルス感染に関連している。ラットでは、妊娠期のビタミンD欠乏は統合失調症を示唆する神経学的異常を引き起こす可能性がある。[18]。McCannとAmes [14]は、詳細なレビューを終えて次のように述べている:「ビタミンDが脳の発達と機能に関与しているという証拠は強力である。ビタミンDは脳の進化、特に現代人の大脳新皮質を形成するのに必要な膨大な数の有髄ニューロンの発達と結合に関与している。
脳の温度を最適な範囲に保つために、発汗などのメカニズムを追加することは確かに有益である。しかし、エクリン腺の増設は無毛に対する直接的な反応ではなく、おそらく拡大するサバンナでの過熱に対する適応的な反応だったのだろう。無毛自体にも欠点があるため(下記参照)、このような重大な出来事には直ちに有益な結果がもたらされる必要がある。ワクチンロンスキーは[2]、「裸の皮膚そのものが、私たちの大きな脳を含む、他の特徴的なヒトの形質の進化に重要な役割を果たした」と示唆しているが、これがどのようにして起こったのか正確には説明していない。エクリン腺の増加だけで、過去2世紀の間にヒト属で起こった脳の大きさの大きな増加を説明できるとは思えない。同様に、ビタミンDレベルの増加は大きな脳を作るのに有利ではあるが、その恩恵が明らかになるまでには何世代もかかる。つまり、ビタミンDの増加は僥倖であり、無毛の始まりにビタミンDの遺伝が関与したとは考えられない。蒸発冷却説もビタミンD説も、すぐに利益をもたらすものではない。無毛と同時に何らかの直接的な利点が生じるような理論が必要である。このような理論は、本稿の残りの部分で詳しく説明する。この理論によって、過去2世紀の間に私たちの属の脳容積が大幅に増加したのは、無毛によって始まったと説明することができる[図1]。
哺乳類では、体温維持に必要なエネルギーはエネルギー貯蔵の異化によって生み出され、おそらく哺乳類の毛は内熱を保存するための適応として発達した。そのため、保温性の高い毛が失われた後、体温を維持するためには、周囲温度が37℃を大きく下回るような場合、エネルギー不足に陥ることになる。例えば、アフリカの2ミヤでは、ますます開けたサバンナの寒い夜、あるいは木の上や洞窟の中にいるような場合である。このような明らかに有害な遺伝子の突然変異が、なぜこれほど成功したのだろうか?無毛がどのような利点をもたらすのだろうか?
毛がないことで、光を媒介としたエネルギー合成が可能になるのだろうか?これはそれほど突飛な考えではない。毛のない人間の組織に作用する低レベルの赤色および近赤外放射(R&NIRR)が、この地球上の生命の普遍的なエネルギー通貨であるATPの産生を刺激することはすでに知られている。低レベルのR&NIRRは、フォト生物調節(PBM)[19]、または低レベル光線療法(LLLT)として知られる非侵襲的医療の基礎となっている。PBMは、光線力学療法や、急成長している光遺伝学の侵襲的科学と混同してはならない。PBMは、創傷治癒、変形性関節症、腱障害[19]、心筋梗塞[20]、特に頚部痛[21]、経頭蓋的には塞栓性脳梗塞[22]、重篤なうつ病[23]、慢性外傷性脳損傷[24]など、さまざまな病態に有効である。しかし、PBMの効果は二相性またはホルミシス性であり、照射量を最適な範囲を超えて増加させると、効果が減少することがある。PBMは、経頭蓋磁気刺激や関連技術である経頭蓋直流電流刺激といった他の非侵襲的療法と類似点がある。3つとも似たような症状の治療に効果的であるが、これらはすべて、最終的には神経細胞の電子輸送鎖(ETC)に非侵襲的に電子を誘導する形態であると考えられるので、驚くには当たらない。いずれもATPの脱合成が起こり、100以上の遺伝子の発現が有意に変化する。[25], [26]。直流電流刺激は、当初は痙攣や失神が報告されるなど、安全性に問題があったが、[27]、現在では克服されている。ヒトを対象とした最近のブレイクスルー論文 [28] では、左海馬との強い結合が確認された皮質領域に経頭蓋磁気刺激を集中させた。右の海馬は異なる働きをするようだ。
動物では、ATPを合成する主なメカニズムは、細胞のミトコンドリアにおける酸化的リン酸化(oxphos)プロセスである。ヒトの場合、脳神経細胞はエネルギー源として解糖を好むが、これはおそらく、脳に栄養を供給する動脈や細動脈の壁にグルコーストランスポーターが豊富に存在することと関係している。Oxphosは4つのタンパク質酵素複合体からなる(最終酵素ATP合成酵素を除く)。一連の酸化還元反応において、電子はある複合体から次の複合体へと極めて高速で受け渡され、ETCを下ってギブスの自由エネルギーを解放し、ミトコンドリア内膜を横切ってプロトンを送り出すことによって電気化学的勾配として蓄えられる。これらのプロトンは、ATP合成酵素によってATPを生成するために使われる。酸化的リン酸化のETCは、光合成の極めて効率的な第一段階と類似している。
PBMがその効果を発揮する正確なメカニズムは、まだ不完全に理解されている。[29], [30]。しかし、R&NIRRは、細胞のミトコンドリアにあるオキシホス複合体IV、シトクロムcオキシダーゼ(COX)によって吸収されることが知られている。一酸化窒素(NO)と活性酸素種(ROS)は断続的に発生する。NOと活性酸素はどちらもシグナル伝達分子として認識されている。Lane[31]は、「(R&NIRRの)即時的な効果は、ATPレベルが煽られるエネルギーバズである……数時間後、110の遺伝子の活性が協調してシフトする」と述べている。Pastoreら[32]は、ミトコンドリアの光受容体であることを示す精製COXを調製し、彼が使用した赤色レーザー光は、COXタンパク質に埋め込まれた金属イオン(ヘム鉄、銅、亜鉛、マグネシウム)と直接相互作用することを提案した。COXのスペクトルには少なくとも5つの吸収ピークがあり、それらは405,625,680,760,820nm付近である[33]。PBMは通常、レーザーまたは非コヒーレント発光ダイオード(LED)により、1つまたは複数の長いピーク(浸透深度が深い)の周辺に照射される。
PBMは、オキシホスETCの活性の上昇を伴い、その結果、ATPの量が増加する(ATP外合成)。PBMの遺伝的影響には、酸化還元感受性転写因子NF-κBの活性化が関与しており、その後、10の異なる機能グループにおける111の下流遺伝子の発現に影響を及ぼす。[25]。全体的なNF-κBは、脳の可塑性、学習と記憶にも関与しており、自然免疫遺伝子と適応免疫遺伝子を制御し、感染に対する応答にも役割を果たしている。
ハンブリンはこう書いている[34]:
「分子による光子の吸収は電子励起状態を引き起こし、その結果、電子伝達反応を加速させる。電子伝達が促進されれば、必然的にATPの生産量も増加する」
Yuら[35]も同様の概念を示している。光吸収分子は通常、獲得したエネルギーを熱として放散する。しかし、光合成鎖や呼吸鎖の吸収分子のように、そうでないものもある。これらは、電子伝達のような生物学的に有用な機能に従事する能力のために、2.4億年以上前の進化によって特別に選択された。酸化的リン酸化と光合成の第一段階は、どちらも様々なチトクロムを用いたETCを用い、非常に効率的にATPを生成するという点で類似したプロセスである(ただし、光合成の全体的な効率は他の理由により低下する)。毛のないヒトでは、酸化的リン酸化のETCが、クレブスサイクルのNADHとFADH2からの電子だけでなく、光光子によってエネルギー化された電子によって駆動されない理由はない。実際、これがPBMの基本である。また、光を利用してATPを合成する動物はヒトだけではない。アブラムシは、カロテノイド光受容体を自ら合成し、ETCを通過する光生成電子を生成することが報告されている。[37]。スズメバチ [38], [39]もまた、光を利用して余分なATPを生成する可能性がある。
R&NIRRは皮膚の数cm下まで浸透することが知られており、[31]、その結果、表在性四肢筋におけるATP外合成が可能になる。最近、810nmの低レベルのNIRRが潜在性トランスフォーミング成長因子β1を活性化し、ヒト歯幹細胞の分化を開始させることが示された。[40]。R&NIRRが頭骨を伝染するのはより困難であるが、ニューロンは容易に励起され、0.1または0.2J/cm2という低いフルエンスがヒトの大脳皮質に対するPBMとして有効であることが報告されている。[24], [41]。無毛が有用な適応と見なされるのであれば、日光に直接さらされる頭皮の面積が広いのに、なぜ光を散乱する毛で覆われているのか不思議に思うかもしれない。ヒゲの生え方や男性型脱毛症は、無毛遺伝子Hrと同様にジヒドロテストステロンによって制御されていることが知られている。

以下の簡単な要約は、動物およびヒトに経頭蓋的に投与されたPBMの効果を示している。ウサギの脳塞栓によって誘発された脳卒中において、LapchakとDe Taboada [22]は、塞栓によって影響を受けた皮質のATPレベルが低下することを示したが、PBMがこれらのレベルを有意に回復させることも示した。Rojasら[42]は、恐怖条件付けラットを用いた実験で、PBMが前頭前皮質におけるCOX活性を上昇させ、酸素消費量も増加させることを見いだした。COX活性はPBMの24時間後にも有意に上昇し、ラットの恐怖記憶を消滅させ、その再発を防ぐ能力が増強された。脳血流の増加は、PBMの結果として必要とされる余分な酸素を供給するために必要な付随物:Linら[43]は、ヒト海馬細胞株を用いたメチレンブルーの研究でこれを発見した。メチレンブルーは合成電子伝達物質であり、脳血流、脳内グルコース摂取量、酸素消費量を有意に増加させ、ミトコンドリア機能を高めることを示した。これらの論文[42]、[43]は、PBMの脳への作用から生じる脳代謝および血行動態の亢進という概念を示している。これはもともと、無毛が出現した後、夕暮れ時の自然なR&NIRRの日常的な結果として起こったものであり、大きな進化の進歩につながったことが示唆されている。
ヒトにおける類似の研究では、Naeserら[24]によって、慢性外傷性脳損傷の2人の被験者の治療に経頭蓋的に繰り返しPBMが使用され、納得のいく成功を収めた-633nmと870nmのLEDライトが、LEDの下で髪を注意深く分けてから使用された。死後カルバリアの研究では、NIR光子の0.2%または0.3%しか白質に伝染しないことが示された[24]。これに基づくと、大脳皮質でのフルエンスは0.24 J/cmであった。Naeserの結果[24]がより多くの数で再現されれば、慢性外傷性脳損傷や外傷後ストレス障害などの治療抵抗性疾患に対する、費用対効果の高い主流治療としてのPBMの有効性が証明されるに違いない。
重度のうつ病に関するこれまでの研究では、電気けいれん療法と経頭蓋磁気刺激の有効性はほぼ同等であったが、PBM [23]を用いた単回治療の小規模試験では、これらの治療法と比較して良好な結果が得られており、さらなる調査の実施が推奨された。興味深いことに、Timonenら [44] は、北緯65°のフィンランドにおける季節性情動うつ病(SAD)に対する明所光治療の非対照研究において、SADの光治療は視覚オプシンを介してのみ作用するという通説に挑戦し、外耳道にLEDを介して明所光を与えて13人の患者を治療した。治療は週5回、4週間行われ、13人の患者のうち10人が完全寛解(ハミルトンうつ病スコア<7)を達成した。別のSAD研究では、Averyら[45]は、「夜明けのシミュレーション」による照明が、明るい光または「プラセボ」(薄暗い赤色の光)よりも効果的であることを発見したが、驚くべきことに、明るい光と薄暗い赤色のプラセボは等しく効果的であった。篩骨板は頭蓋骨よりも薄いため、例えば大脳基底核は鼻からの照射がより容易である。
Chenら[30]は、マウス胚線維芽細胞に810nmの近赤外レーザー光を0.003~3J/cm2のフルエンスで照射した。その結果、フルエンス0.03から0.3 J/cm2の間でATP合成が大きく増加し、数時間後にはベースラインまで減少した。彼らはまた、NF-κBがレーザー誘起活性酸素の発生によって活性化されることも示した。抗酸化剤の添加は、活性酸素と同様にNF-κBの活性化を阻害したが、ATPの外部合成は阻害しなかった。研究者らは、レーザー光が電子輸送を増加させ、活性酸素を発生させるリークが起こることを示唆している。光による代謝効果(ATP外合成)はほぼ即座に起こるが、遺伝的効果は遅れて起こる。遺伝的効果はシグナル伝達メカニズム(活性酸素)を必要とし、NF-κBの細胞核への移行を伴うが、ATPの外部合成はNIRRが酸化的リン酸化の光受容複合体に直接作用するため、完全にミトコンドリアで起こる。NF-κBが制御する下流遺伝子は、「ミトコンドリアのエネルギー産生を長時間上昇させる」その大きさに比例して、脳は膨大な量のATPを使用する。ここでは、このような大量のATPが、古代のアフリカの夕日のR&NIRRによって、数少ない近縁種で無毛の初期メンバーの脳で初めて生成されたことが提案されている。これによって、上述した大脳の代謝と血行動態の亢進も可能になった。

セクションスニペット
仮説
200万年前頃、初期のホモ属の種でランダムな突然変異、あるいは一連の突然変異が起こり、近縁の数個体が完全に無毛になったと考えられている。通常不利な突然変異がそうであるように負の選択に屈するのではなく、急激な選択的掃討を受けたのである。なぜか?その突然変異は、当時のアフリカでは小さな不利を大きく上回る大きな利点をもたらしたからである。PBMの研究が示すように、それはATPエネルギーの外部合成を可能にし、またR&NIRRが当時の無毛の頭蓋骨を貫通し、多数の遺伝子の発現を有利に変化させ、大脳の代謝と血流動態の向上を可能にした。これが、ホモ属の出現後に起こった急速な知性主導の進化的脳拡大の出発点であったことが示唆されている(図1)。PBMの二相性については、毎晩の夕暮れ時に生理学的に正しい低線量のR&NIRRを浴びることが、本来の自然なPBMを構成することが示唆されている。この放射線の後、12時間の暗闇が続き、その間に放射線の遺伝的作用が定まる。夜明けに同じ放射線を照射しても、その直後にすべての可視波長の過剰な線量が続くため、効果には疑問が残る。私たちの祖先が無毛にならなければ、上記のような影響は起こらなかっただろう。無毛の原因となる突然変異が、遺伝子の急激な掃引によって即座に取り込まれたのは、無毛とその恩恵が同時に起こったからである。
さらなる考察
大量のATPが赤血球(RBC)によって運ばれることを述べておく [47]。毛細血管に到達すると、その一部は血管拡張物質およびシグナル伝達分子として血漿中に放出され、余剰分は血漿中の酵素ectoATPaseによって分解される。ATPは基質のリン酸化によって産生されると考えられる。赤血球は核もミトコンドリアも持たないが、10-19個の遺伝子のmRNA転写産物を持っており、[48]、その半分以上が細胞代謝に関係している。ヘムはポルフィリン分子で、UVB、UVA、バイオレット/ブルー領域に強い吸収帯を持つ。
光子のエネルギー=波の周波数×プランク定数というプランク・アインシュタインの方程式が示すように、青色と紫外線の光子はいずれも赤色よりもエネルギーが高い。皮膚での散乱と吸収は、これらの短い波長の方がはるかに大きいが、組織への浸透は少ない[49]。Liebmannらによるヒト皮膚由来のケラチノサイトと内皮細胞への青色光照射に関する研究 [50]では、より短い青色波長(412-426nm)の非常に高いフルエンス(500J/cm2以上)は細胞毒性を示すが、高いフルエンス(100J/cm2)はT細胞に対して依然として細胞毒性を示すことが示された。これらの研究者らは、453nmの青色光照射後の細胞のエネルギー状態を研究し、ATPレベルの有意な増加を発見した。次にメラニンについて示唆されたように、真皮と表皮下部を循環する血液の静脈流出において、ブルーライトがATPの再生を助ける可能性がある。
さて、チェルノブイリの焼け焦げた原子炉の壁に生えた真菌の奇妙な話である。この真菌、クリプトコッカス・ネオフォルマンスは通常メラニンを合成するが、バックグラウンドの500倍のガンマ株を浴びると、非照射細胞や照射されたアルビノ変異体よりも著しく速く成長することが判明した[51]-「放射線合成」と呼ばれる現象である[52]。メラニンはキノン部分(酸化的リン酸化複合体IIの分子に似ている)を含む生体高分子であり、これがガンマ株被曝後のメラニンの酸化還元挙動とその結果としての電流発生を担っていると考えられている[52]。この電流は、還元剤を加えることで増加した。可視光線を照射するとメラニン生体高分子中のキノン分子の割合が増加し、紫外線を照射するとNADHを酸化するメラニンの能力が劇的に増加すると言われている。[53]。Dadachovaら[51]は、メラニンが代謝エネルギーのために電磁波を利用する能力をメラニン生物に与える可能性を「慎重に示唆」した。Bryanら [53]は、メラニン化したC. neoformansとメラニン化していないC. neoformansの酸化的リン酸化の可能性を調べた。
Bryanら[53]は、メラニン化したC. neoformansとメラニン化していないC. neoformansのガンマ株、紫外線、可視光線照射後のATP生成能を調べた。
この興味深い結果は、おそらくATPパラドックス[54]に関わるものであり、著者らはメラニンがクロロフィルと同様のエネルギーハーベスティング能力を持つ可能性を提起した。ユーメラニンの固体ペレットに関する研究は、ユーメラニンが電子とイオンのハイブリッド伝導体であり、水とドープしてフリーラジカル、電子、プロトンを生成できることを示している。[55]。
真菌から哺乳類への外挿は大きな飛躍であるが、C. neoformansのメラニンの種類であるユーメラニンは、生きているアフリカ人の皮膚で生成されるものと同じである。白人におけるフェオメラニンの吸収スペクトルは、R&NIR領域でより急速に減少するものの、類似している(図2)。酸化的酸化リンの基本形は、好気性真菌とヒトで類似している。[56]。ヒトでは、太陽光線とメラニンとの相互作用により、間接的なATP外合成が行われている可能性がある。メラニンのキノン部位は、光フォトンによって活性化された電子を、酸化的リン酸化のキノンプールに供与し、その後、メチレンブルーと同様の方法でETCを下降すると考えられる[43]。キノンプールには、酵素と緩やかに結合したキノン分子が存在する。キノンは、デヒドロゲナーゼから電子を受け取り、オキシダーゼに電子を供与することで、ETCの酸化還元反応を促進する。ヒトのメラノサイトやケラチノサイトのメラノソームは、ミトコンドリアとは別の小器官であるため、細胞質のNAD+がメラノソームから2個の電子と1個のプロトンを受け取り、還元型NADHになることが示唆されている。その後、NADHはミトコンドリア内膜を通過することができないため、いくつかのミトコンドリアシャトルのうちの1つによって膜に運ばれる。その電子はFADに送られ、FADH2を生成し、その後ETCに入ることができ[57]、ATPの外部合成をもたらす。
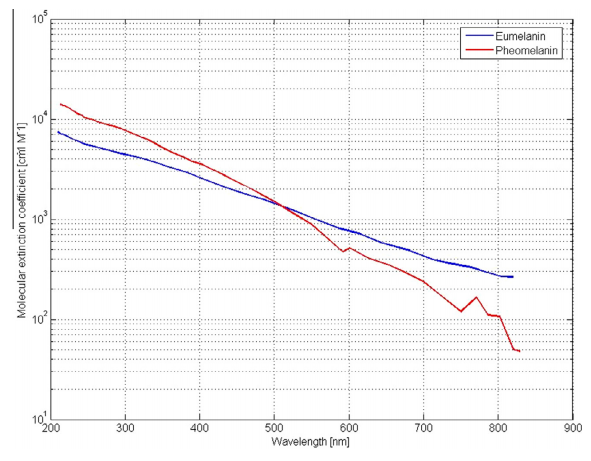
図2. ユーメラニンとフェオメラニンの吸光度とEMR波長 – wild commons.
上記のシナリオは、Dadachovaのグループによって行われた実験によって支持されている[51]。彼らは、ガンマ株、紫外線、可視光線、赤外線のいずれかを照射した場合としなかった場合の、NADHの酸化とフェリシアン化物の還元を含む反応において、電子伝達物質として働くメラニンの能力を測定した。4種類の放射線はすべて、電子伝達の顕著な増加を引き起こし、驚くべきことに、その増加は入射光子のエネルギーに依存しなかった。
これまでのまとめとして、PBMの研究は、毛のない状態において、R&NIRRによるATP外合成が試験管内で行われ、生体内でも行われることを示している。さらに、日光とメラニンから表皮細胞でATP合成外合成を起こすメカニズムが提唱されている。ATPの外部合成は、無毛が最初に200万年前発生したときに生じたエネルギー損失を補って余りあるものである。PBMの研究では、R&NIRRが多数の遺伝子の発現を変化させ、大脳の代謝と血行動態を促進することも示されている。
仮説を検証する
ATPは非常に儚い分子で、すぐに使い切られるか、他の高エネルギー化合物に変換されるか、ホスホクレアチンとして貯蔵される[58]:
ATP=GTP þ クレアチン $ ADP=GDP þ ホスホクレアチン:
ホスホクレアチンは、筋肉が必要とするときに、ATPの迅速な供給源として使用することができる。31リン磁気共鳴スペクトロメトリーやNIRスペクトロスコピーなど、高エネルギー分子を追跡する生体内試験の方法は、急速に進歩している。[58,59]。陽電子放射断層撮影法は、グルコースの取り込みを追跡するために使用することができる。ミトコンドリアの酸素消費量も非侵襲的に評価できる。エネルギー調査の詳細はもちろん研究者に委ねられるが、サハラ以南のアフリカ人と白人という2人の健康な若年成人を4時間絶食させ、暗闇の中で酸化的リン酸化の基本レベルを確立し、その後、日中と夜明けと夕暮れ時の日光浴を含む24時間の間隔で高エネルギー化合物のレベルを測定することが想定されている。脳のエネルギー状態や大脳の酸素・グルコース消費に対する日光の前後効果を評価するために、頭を剃ることに同意した被験者から選ぶこともできる。予備実験として、皮膚と四肢の筋肉に及ぼす影響を前腕だけで調べることもできるだろう。
考察
ATPが大量に合成され、脳のエネルギーと遺伝子発現に予想通りの変化が起きたと仮定すると、このことは、初期のホモ族が12時間の日照時間のほとんどを、サバンナの薄日が差す中で活動していたと想定されることと関係しているはずである。無毛の個体にとっては、余分なATPと大脳の利点が狩猟能力を大きく向上させ、大きな競争上の優位性を与え、動物性タンパク質をより多く食事に加えることを可能にした。いくつかの近縁種が無毛になったとき、狩猟能力が大幅に向上した、より熟練した、よりエネルギッシュな初期人類は、有毛種の配偶者として大きな需要があることに気づくだろう。彼らの無毛の突然変異は、遺伝的掃討作戦によって急速に選択されたのだろう。残念ながら、そのような掃討作戦の証拠は、発生から25万年後までしか発見できない[60]。

無毛化はいつ起こったのだろうか?MC1R遺伝子座に影響を及ぼす突然変異の考察から、Rogersら[8]は、ホモ属は少なくとも120万年前から無毛であったと結論づけた。無毛とそれに伴う自然PBMがヒトの大脳の進化の原動力であるとすれば、ヒトの進化の過程のある時点で、無毛の出現が脳容積の増加速度の識別可能な上昇によって示されることが予想される。無毛の発生はそのような上昇に先行し、エネルギー消費量の多い神経組織のエネルギー需要が増加したことを示す証拠がそれに続くかもしれない。過去650万年間の脳容積の増加率は一定ではないことが判明した。図1に示すように、200万年あたりから、大きな、そして継続的な増加が始まっている。重要な論文[61]によると、この時期には、食事の質とエネルギー密度の向上が伴っている。しかし、初期ホモ・エレクトスの大きな脳のエネルギー需要の増加は、他の組織、特に腸や筋肉のエネルギー需要の減少によって覆い隠され、エネルギーの安い脂肪貯蔵が増加した。Leonardら[61]は、H. erectusは全体的に大きな動物であるが、比較的「筋肉不足で脂肪過多」であると述べている。しかし彼らは、”このことは、食生活の変化が人類進化の過程で脳を大きく拡大させる原動力となったことを意味するものではない”と指摘している。ここでは、原動力は無毛と自然なPBMであり、無毛の出現は上昇の始まりの直前、200万年前頃に起こっただろうと示唆している。図1と、ホモ属の出現後に爆発的に増大した脳の大きさを見ると、進化は200万年前の間に、一般的に有利な遺伝子の選択から、特に神経学的に有利な遺伝子の選択へと、その目的を再優先させたのではないかと思われる。種全体が無毛になった後でも、よりエネルギッシュで器用な無毛の個体に対する最初の交配相手の魅力は常に続いていたことが示唆される。しかし、今世紀になっても知能が進化しているかどうかは議論の余地がある。人類の進化は今や、適者生存というよりも、万人の生存を目指すようになっているのだ。ヒト属の進化が進むにつれて、無力で未熟な幼児が生まれることが多くなった。このことは、アウストラロピテクスで推定されていた一夫多妻制を捨て、一夫一妻制、あるいは少なくとも連続一夫一妻制を採用する一つの正当な理由となるだろう。親が二人いる文化は、母子により良いものを与えることで乳児死亡率を低下させる。
脳の大きさから判断すると(図1)、2ミヤからごく最近まで知能が進化してきたのは200万年前からごく最近までの知能の進化は、現在の進化よりも議論の余地が少ないかもしれない。知能には、環境の変化に適応する能力も含まれる。200万年の気候は現在と似ていたかもしれないが、私たちのように地球温暖化に直面するのではなく、更新世初期の人類はより寒い乾燥した気候に直面していた。アフリカの熱帯雨林は乾燥し、サバンナは拡大していた。広々としたサバンナでは捕食者が危険なため、知能に関連した形質に進化的に焦点を当てる必要があった。知性に焦点を当て続けたことで、過去200万年の間にヒト属の脳が大きく拡大したことを説明することができるが、この進化のきっかけは、自然のPBMを容易にする無毛であった。
遺伝子研究者たちは[62]、前頭前皮質がヒト特有の遺伝子で濃縮されていることを発見した。彼らはまた、霊長類特有の転写因子制御因子を多数同定した。そして、少数の転写因子の変化が、ヒトとチンパンジーとの間の遺伝子発現の大きな違いを決定している可能性が示唆されている。[63]。
[64]のような研究は、知能が遺伝性であることを疑う余地はないが、おそらく多くの遺伝子が関与しているため、どの遺伝子も有意な影響を及ぼしていない。しかし、いくつかは注目に値する。最近同定された遺伝子NPTN [65]の塩基配列における一塩基多型(SNiP)は、シナプス可塑性や皮質の厚さ(それ自体が知能と関連している)と関連している。興味深いことに、NPTNの発現は両半球で不均等であり、左半球の方がSNiPに敏感であることが判明した。脳容積と知能の様々な尺度との間には、不完全ではあるが一般的な相関関係がある[66]。しかし、ホモ・フローレシエンシス、別名ホビット族は、初期のホモ種の島嶼小人型であったと思われ、頭蓋容積はわずか400ccで、約13,000年前まで生存していた。しかし彼らは石器を作り、狩りをし、ステゴドン(小型のゾウ)を絶滅させたかもしれない。神経細胞の数よりも神経回路の方が重要であることを示唆している。

2005年には、同じファミリーに属する2つの遺伝子の対立遺伝子に関連する、よく引用されるが物議を醸す2つの論文が発表された。[69,70]、
MCPH1とASPMである。最近生まれたこれらの遺伝子は、まだ強い正の選択と適応進化を受けていることが示唆された。しかし 2008年の論文[66]では、これらの対立遺伝子はもはや進化しておらず、”とっくに固定化”に達しており、現在では一般的な認知とは関連していないが、これらの対立遺伝子の適応が過去200万年の間に脳の大きさが劇的に増加したことと関連していることがわかった。
もう一つの興味深い遺伝子は、1/200万年前以上前に有利な突然変異を起こした転写因子FOXP2である。この遺伝子は発話の明瞭さと理解力に関連する遺伝子としてよく知られている。マウスとヒトの間では、その配列はわずか2アミノ酸しか違わず、最近、2アミノ酸をヒトのものに置き換えることによって、マウス遺伝子がヒト化された[71]。これにより、皮質-線条体経路に有益な変化が生じ、マウスの学習能力が向上した。ヒトの場合、共著者のアン・グレイビエルによれば、この突然変異によって、意識的な連想を伴わずに、ほとんど自動操縦のような意思決定ができるようになるという。彼女はまた、「脳がどのようにして、意識的に何かを考えることから無意識に行うことへと移行するのかは誰も知らないが、ヒト型のFOXP2を持つマウスはずっとうまくいった」と述べている[72]。
アフリカの祖先は、更新世初期の気候が涼しくなっていることを知らなかった。
特に移住を考えていたのであればなおさらである。しかし、初期のH.エレクタスの一群は、まさにそれを実行した-移住したのである。グルジア共和国の現在の町ドマニシ近郊で、180万年前とされる彼らの頭蓋骨が発見されている[73,74]。H.エレクトスの体格と皮下脂肪の増加は、寒さを和らげる一因になるだろうが、180万年前は、動物の皮を身につけたり火を使ったりするようになったとされる最古の年代よりも前である。しかし、ドマニシで発見された数個の頭骨にもかかわらず、このような初期の人々の多くは、その後も生き延び、繁殖したのかもしれない。それは誰にもわからない。更新世後期の厳しい氷河期はまだ始まっていなかった。
アフリカからの主な脱出は比較的最近、60-70k年頃に起こったと考えられている。その頃には、更新世後期の氷河期はすでに確立しており、73k年のトバ山の大爆発は、さらに一時的な冷却を引き起こしただろう。北方気候の弱い冬の日照は、無毛によるエネルギー不足を補うことはまったくできなかっただろう。しかしその頃までには、洞窟に住み、動物の皮を被り、火を使い、さらに脳を2mほど進化させたおかげで、ホモ・サピエンスはヨーロッパと北アジアで氷河期の厳しさを生き延びることができた。

どのような遺伝子変異が無毛を引き起こしたのだろうか?無毛遺伝子Hrは皮膚と毛包の発達を制御している[75]。Hrは皮膚・毛髪と脳に広く発現している。ビタミンD受容体(VDR)の共抑制因子として働く。転写されたHrタンパク質のレベルは、保存された非コードのオープンリーディングフレームORFU2HRによって制御されている。万能脱毛症を引き起こすHrの劣性突然変異は、血縁関係にある家系で報告されている[76]。ORFU2HRの優性突然変異 [77] は思春期以降に進行性の脱毛を引き起こし、眉毛やまつ毛にも影響を及ぼし、Hrタンパク質の制御不能な高濃度に関連する。Hrの遺伝子配列には、ヒトと現存するチンパンジーとの間で、同義および非同義の変異が多数存在する[60]。200万年前の間にどのような遺伝的事象が起こったかを特定することは難しいが、Hr遺伝子またはVDRとの相互作用が関与している可能性がある。また、X連鎖性の「狼男症候群」[78]の遺伝学的解明も役立つ可能性がある。
無毛の突然変異は、マウス、ネコ、イヌなど他の動物でも時折起こる。しかし、小型の内温性無毛哺乳類は、野生で生存し増殖するには、体重に対する表面積の割合が大きすぎる。小さなハチドリは、起きている時間のほとんどすべてを餌に費やす必要があり、眠っているときは体温が劇的に低下する。哺乳類は緯度が高いほど大きくなる傾向があるため、内熱を保持しやすくなる(バーグマンの法則)。逆に、人間のピグミー種は熱帯地方(コンゴ、アンダマン、フローレス諸島など)にしか生息していない。ホモの系統に無毛が出現すれば、それ自体が熱損失を促進し、アフリカの200万年前でも大きな体の発達を促しただろう。
この仮説には2つの仮定がある。1つ目は、200万年前以前の祖先の毛髪は、散乱や反射によってNIRR(700~1100nm)を遮断していたというものである。PBM/LLLT分野の研究者たちは、これが真実であると仮定しているが、直接的な裏付けとなる証拠は不足しているようだ。2つ目の仮定は、無毛は当初200万年前の完全なもので、頭皮にも及んでいたというものである。
まとめ
PBMとR&NIRRの効果に関する研究の結果、ヒトの脳の進化の最後の200万年の主なきっかけは無毛であり、それによってATPの外部合成と大脳の代謝および血流動態の向上が可能になったという仮説が提唱されている。これが、180万年前頃に始まった進化において、非常に急速な脳の拡大を促した。更新世の気候がまだ安定していた200万年前の間、アフリカで無毛になることにはすぐに利点があった。しかし最近になって寒くなり、頭皮の毛髪が再び発達し、衣服を着用する必要が出てくると、ヨーロッパと北アジアでは利点が負債に変わった。ビタミンDの合成は無毛の恩恵を受けたが、これは僥倖であった。ビタミンDは脳の進化に重要な役割を果たし続けているが、衣服によってその合成が制限され、欠乏症は特に高齢者に多い。
この仮説によって、特に神経疾患に対する非侵襲的で効果的な治療法として、[24]のように繰り返し治療を行うPBMの研究がさらに進むことが期待される。
利益相反
著者に利益相反はなく、資金源もない。