Where Does the Body’s Vitality Come From?
www.midwesterndoctor.com/p/where-does-the-bodys-vitality-come-1cc
2023/10/21
一目でわかるストーリー
- 水は固体、液体、気体であることに加え、液晶のような4つの相を持ち、全身にこのゲル状の状態で存在する。
- 液晶水は固体の強さと液体の流動性を併せ持ち、生命維持に必要なバイオテンスグリティーのような機能の多くを促進するために、身体が利用している。
- この水が存在することで、細胞の完全性、組織の圧縮に対する抵抗力、組織間の潤滑性がどこから来るのかが説明でき、一方、この水が存在しないことで、多くの疾病状態がどこから来るのかが説明できる。
- 液晶水は、身体の謎の多くを理解する方法を提供し、身体とつながる多くのヒーラー(マッサージセラピストなど)が身体の中で観察していることを言葉にする方法を提供する。
100年以上前から、科学者たちは水が単なる固体、液体、気体という古典的なモデルには当てはまらない、さまざまな変わった性質を持っていることに気づいていた。ジェラルド・ポラックはこれらの観察を基に、負に帯電した表面が存在し、適切なエネルギー源(特に赤外線-どこにでもある)が存在する場合、水はそれ自体が液晶のように振る舞うH3O2という式のオフセットした六角形のシートの層になることを最終的に突き止めた。
注:光に加えて音もH3O2を生成することができる。
私の目から見ると、液晶水は科学的に極めて過小評価されている分野である。なぜなら、説明のつかない身体の謎の多く(例えば、液体を動かすポンプが知られていないにもかかわらず、なぜこれほど多くの液体が体内を流れているのか)を説明するメカニズムであり、私たちが体内で頻繁に観察することの多くを言葉にする方法を提供してくれるからだ。
注:この記事の以前のバージョンは7カ月前に掲載されたものである。これは非常に重要なテーマだと感じているので、その記事は改訂され、本日再公開された。
液晶水とは何か?
液晶水はかなりの固さを持っており、(H2OではなくH1.5Oであるため)変位した水素原子を含め、そこに入るほとんどのもの(例えば、ポリスチレンの微小球)を追い出す。変位した正電荷の水素原子(以後プロトンと呼ぶ)は、この格子のすぐ外側に集まり、pHと電荷の勾配を作り出し、それを測定することができる。
多くの場合、このH3O2格子は巨大になり、ポラックらは0.1ミリメートルから0.5ミリメートル(10万から50万ナノメートル、nm)の大きさを測定している。細胞はこの水に依存しているため、水が形成される表面を多数含んでいる。例えば、細胞の20%は細胞骨格(細胞の構造を維持するタンパク質の格子)で占められており、高電圧電子顕微鏡写真の分析から、細胞骨格内に存在する水の半分以上が、H3O2を形成する可能性のある表面から5nm以内にあることが示されている(参考までに、H2O1分子の大きさは0.27nmである)。
細胞内では、水が構造化できる表面部位が非常に密接に集まっているため、顕微鏡で生きた細胞を観察していると、液晶性水(一般にEZ水とも呼ばれる)が非常によく見えるのは理解できる(実際、H3O2に関する初期の研究の多くはここから始まった)。つまり、なぜ細胞はこれほど多くの液晶水を作り出すように設計されているのだろうか?
注:液晶水(H3O2)とその物理的性質に関する研究の長い歴史については、こちらの記事を参照されたい。
細胞の謎
前回の記事で、私は科学の世界でよく見られる問題について述べた。自然のプロセスを説明するために誤ったモデルが利用されると、モデルの予測と現実との間に食い違いが生じるのは避けられない。このようなことが起これば、その誤ったモデルを支持する人たちは自分たちのモデルを再検討するようになるだろうと期待するだろうが、そうではなく、そのモデルには多くのことが投資されているため、彼らはそのモデルに対するあらゆる挑戦を非難し、既存のモデルの失敗を説明する無数の創造的な方法を考え出すだろう。
例えば、ワクチンの最初の約束(2回接種すれば完全に免疫ができ、伝播は止まり、COVID-19は急速に記憶の彼方に消えていく)を考えてみよう。ワクチンの臨床試験が不正であったため、ワクチンの約束は何一つ実現せず、COVIDの状況はかえって悪化した。しかし、医療当局(および医学界)は自らの過ちを認め、COVID-19を管理するための別のアプローチに切り替える代わりに、ワクチン接種の義務化を倍加させ、集団接種(およびブースト)が実際に達成するはずだったことに関して何度もゴールポストを動かした。
同様に、細胞生物学に関しても、細胞に関する私たちの知識は驚くほど原始的であり、既存のモデルでは細胞環境で何が起こっているのか説明できないことが多い。しかし、代替モデルが科学界に一般的に受け入れられていないため、私たちは生命の無数の謎を多少なりとも説明できるように、既存のモデルに絶えずパッチを当て続けることを余儀なくされている。
現時点では、このような状況の主な原因のひとつは、科学研究が歪められ、産業界が利益を得られる発見を優先するようになったことだと私は考えている。例えば、免疫学は、ワクチン接種や独自の薬剤で標的とすることができる免疫系の側面に焦点を絞っており、その結果、免疫反応の他の多くの構成要素が無視されたままになっている(免疫系は一般に、体内で最も理解されていないシステムのひとつと呼ばれている)。同様に、医薬品は細胞内のレセプターやチャネルに作用することが多いため、細胞生物学は細胞のそのような側面だけに焦点を絞った研究を採用してきた。
その点、水の液晶相は、既存のモデルでは十分に対応できない多くの生物学的現象に、様々な説明を与えてくれる。この記事では、そのうちのいくつかに焦点を当てる。
注:この記事で論じることの多くは、これら3冊の本でより詳しく述べられている(同様に、この記事の参考文献の大半もこれらの本から出典しているので、記事中では繰り返し引用しない)。もし自分でこのテーマをさらに研究したいのであれば、これらの本(すべてジェラルド・H・ポラック著)をこの順番で読むことをお勧めする:
- 『水の第四相: 固体、液体、蒸気を超えて』(2013)
- 細胞、ゲル、生命のエンジン: 細胞機能への新しい統一的アプローチ (2001)
- 「細胞生物学における相転移」(2008)-この3冊の中で最も専門的である。
細胞の完全性:
生物学の大きな謎の一つは、細胞が持つ巨大な耐久性である。古典的なモデル-細胞は液体の袋であり、流体モザイク膜の中に収められている-を考えれば、外力によって細胞が「弾け」、その内容物がすべて流出することは容易なはずだ。しかしほとんどの場合、細胞は大きなストレスを受けてもその完全性を維持する。
細胞は、ギロチンで真っ二つにされたり、(体外受精を行うときなど、特定の成分を分離して作業できるように)引き抜かれて四つ割りにされたり、電気弾で穴だらけにされたりといった外傷に耐えることができる。しかし、いずれの場合も、残った成分の細胞の完全性は維持される。
同様に、細胞の膜が剥がされても、内部の内容物は急速に拡散するのではなく、その場に留まる。また、筋繊維が膜を失っても、その機能的能力(収縮力を生み出す)は無傷のままであることも、50年以上前から知られている。
これらの現象を説明する手がかりが3つある。1つ目は、ポラックが示したように、水滴は驚くべき完全性を持っており、細胞のように、マイクロブレードで真っ二つに切断されても、その完全性を維持する(そしてその後、再び融合する)ということである。第二に、ポラックは水滴がかなりの程度の液晶水を含んでいることも明らかにしている。3つ目は、細胞内の水分子は主にゲル内に存在し、そのゲルは同じ液晶水で構成されているということである。別の言い方をすれば、細胞の安定性は、それを包む外的構造よりも、むしろそれを支えている水の性質が大きいということだ。
ここで、細胞構造のもう一つの重要な側面について考えてみよう。液晶水が形成される条件が細胞全体に存在するため(負に帯電した親水性表面と周囲の赤外線エネルギー)、細胞は厚さ数百マイクロメートルの液晶水層で急速に満たされるはずだ。しかし細胞全体では、表面は数分の一マイクロメートルしか離れていないことが多い。このことは、細胞の構造が液晶水の絶え間ない形成に依存していると同時に、その結晶構造が完全な大きさまで成長することを制約していることを意味している。
テンセグリティ
古典的な建築学では、建物は強固な骨格を持ち、その上に他の構造体が乗っている。例えば、昔は石の柱がよく使われていたが、現在では超高層ビルのニーズを満たすために鉄骨の上部構造が必要とされる:

というのも、生命はそのような建築物とは異なり、素早い動きを必要とするからだ。また、生命体は鉄骨の梁のような強度を持つ強固な構造体を作ることができない。しかし、より複雑な別の建築モデルが開発され、複雑な建築モデルを好む人々によって頻繁に利用されている。
テンセグリティ(tension integrityの略)は、バックミンスター・フラーが最初に提唱したモデルである。このモデルは、一連の非圧縮性構造体を弾性接続の格子(伸ばしたときに張力を蓄えることができる)で連結すると、はるかに強い構造体ができると仮定している。というのも、構造物が受ける力は、単一の構成要素(例えば石柱)に集中するのではなく、それらの弾性的な連結部のそれぞれに均等に分散されるため、単一の構造要素の破断点を超える可能性がはるかに低くなるからだ。
フラーの研究はその後、テンセグリティの原理に基づいて建てられた数多くの建築物に影響を与えた。これは、彼が作ったテンセグリティの球体を持っている古典的な写真:

バイオテンセグリティは、これと同じ仕組みが自然界にも存在することを示したものだ。人体の各組織レベルには、連結した引張マトリックスが存在し、それが安定性を与えている。フランスの手外科医、ジャン・クロード・ギンベルトーも同様に、低侵襲手術の際に体内に設置した小型(拡大)カメラを通して、筋膜(多くの手技療法家が扱う、身体全体に存在する結合組織)のレベルにおいて、連結した引張構造の存在を示す驚くべき仕事をした:
注:ここや彼の他のビデオで使用されている拡大スケールでは、これらの構造をコーティングしている液晶水を直接観察することができる。また、構造体が互いにすべり合うことを可能にする潤滑性にも注目してほしい。この潤滑性が失われると、体内でさまざまな問題が生じるようだ。
年月が経つにつれ、ホリスティック医学の分野では、バイオテンセグリティは身体を理解するための有効なモデルであり、緊張の連鎖したネットワークは身体の最大レベルから最小レベルまで存在する(例えば、細胞骨格は各細胞内の弾性連結単位である)というコンセンサスが高まっているようだ。しかし、この理論は頻繁に議論されるものの、この理論にはまだ2つの大きな未解決の問題があり、液晶水で説明できると私は考えている。
一つ目は、ギンベルトーの研究において、体中の小さな液胞(水で満たされた密閉された区画)が、体のテンセグリティの多くが依存する非圧縮性の基本単位を形成していることが観察されたことである。もうひとつは、細胞内をさらに拡大すると、緊張状態にある細胞骨格は存在するものの、その緊張を生み出すものは何も見えないということである(細胞外マトリックスと細胞の結合が緊張を生み出すのに十分かどうかは大いに議論の余地がある)。テンセグリティー・システムの構造的強度は、構造体の構成要素に張力がかかっているときに初めて現れるからだ。
前者の場合、微小空孔は(圧縮できない)液晶性の水で満たされていると私は考えている。水滴の場合、ポラックは、正電荷を帯びたプロトン(H3O2のシートによって水滴の中心に押し出される)の相互反発が、水滴の液晶境界によって抵抗される外向きの力を生み出すと結論づけた。この2つの力のバランスによって、飛沫は球形になる。私は、同じメカニズムが人体中の微小液胞でも働いていると考えている。
後者の場合、ゲルを作るタンパク質(多くのゲルは99.9%以上が水)によってどれだけの膨張が生じるかを覚えておくことが重要である。さらにポラックは、細胞内で常に形成されようとしている液晶性の水が、液晶性のゲルがその大きさまで成長するために起こるはずの伸張にタンパク質が抵抗するために、その大きさに達することができないことを示している。これは細胞レベル(細胞が最大サイズまで膨張すると細胞骨格が伸び、それ以上の膨張ができなくなる)でも、細胞全体のタンパク質内でも起こる。
タンパク質のレベルでは、ゲル形成に反応して起こりうる最大限の膨張を抑制するために、身体はしばしばタンパク質間の化学結合に頼っている。さらに、タンパク質は折り畳まれたコンフォメーションと展開されたコンフォメーション(例えば、らせんとコイル)の間で交互に変化することがあるが、これはタンパク質にかかる張力(ゲル内の液晶水の膨張圧など)によって決まることが多い。このコンフォメーションの変化には様々な重要な結果があり、これについては後述する(例えば、筋肉の生理学)。
さらに、身体の他の多くの構成要素も液晶水に依存しているようだ:
コラーゲンを研究したメラチーニらは、その三重らせん結晶構造を安定化させる水の重要性を指摘した。メラチーニらは、コラーゲンの三重らせん結晶構造の安定化における水の重要性を指摘した。彼らは、コラーゲンらせん内の水が「結晶格子内の三重らせんを取り囲み、相互に連結する半クラスレート様構造」を形成していることを発見した。骨のマトリックス内の水も同様に高度に構造化されており、その構造化はコラーゲンやプロテオグリカンなどの有機高分子だけでなく、鉱物の表面にも関連していることが判明している。実際、水は、骨マトリックス内でミネラルナノ粒子を平行配列に配向させる上で基礎的な役割を果たしているようで、有機分子がない場合でもこの配向を与えている。
日常的なゲルの膨張
私たちがよく知っている多くの技術(例えば紙おむつ)は、水分を保持する半固体構造に膨張できるハイドロゲルに依存している。同様に、体内の多くの大きなプロセスも、より大きな液晶構造に集合する水の傾向に依存していることを直接観察することができる。
タンパク質の通常の架橋構造では、ゲルの膨張が制限されているため(膨張するゲルに対応するために分離する必要があるタンパク質が、タンパク質間の架橋によって妨げられるため)、組織やタンパク質が損傷を受けると、この制限が部分的に取り除かれる。その結果、人間の組織が損傷を受けると膨張することが、ミクロのスケールで確認できる。同様にポラックは、筋骨格系の外傷を経験したときにも、これが起こる可能性が高いと主張している。ポラックの提唱するモデルでは、最初のゲル形成が既存の断裂をさらに拡大させ、この液晶水の漸進的拡大が最終的に目に見える腫脹につながる。
より大きなスケールで言えば、膝のような体重を支える関節が、日常的に経験する継続的な摩擦によって損傷することなく、その可動域を維持できるようにすることが、身体工学上の大きな課題のひとつである。液晶水の驚くべき特徴は、液晶水が形成される表面が無傷のままであれば、液晶水は破壊されても、ほとんど摩擦のない表面とともに何度でも再生することができるということである。
このため、液晶水は健康な関節が経験するストレスを吸収する成分となり、関節が健康であれば、そのストレスから瞬時に再生することができる。このマイナス電荷を帯びた液晶水の再生層とともに、関節の中心部にはプラス電荷を帯びたプロトンのポケットがあり、互いに反発し合って膨張圧(水滴に見られるような)を生み出している。関節包がこの領域を密閉しているため、これらのプロトンは逃げることができず、したがって効果的に反発する磁石のように機能し(たとえば磁気浮上式鉄道を考えてみよう)、身体の重さに抵抗して関節内の中心空間を維持する。
ポラックの関節モデルで私が最も説得力があると思うのは、膝関節内の滑液の特異的な質である。関節鏡手術中に滑液をカメラで見ると、滑液内の拡散は目に見えて遅くなっている。一方、滑液を直接採取すると(例えば膝の吸引)、かなり濃厚でゲル状の質を持っていることがわかるが、これは私が液晶水の存在を連想して学んだことである。ポラックは同様に、彼のモデルが示唆するEZ(液晶性)水の挙動は、放っておくと半固体でありながら、加えられたせん断力に応じて流動することができるゼラチン状の卵白の挙動に似ていると主張している。
細胞の勾配
注:「勾配」とは、2つの異なる領域にあるものの濃度の差を表す。これには電荷(つまり電池は勾配に依存する)、電解質、温度などが含まれ、多くの勾配は容易に採取可能なエネルギー源となる。
既存の物理学のパラダイムは、次のように主張している:
- 物事の自然な状態は、無秩序で均一に混ざり合っていることである。
- 例えば、結晶を形成したり、2つの領域の間に勾配を作ったりする)何かをより秩序あるものにするときはいつでも、その秩序を作り出すためにエネルギーを消費しなければならない。
- 秩序だった構造が無秩序になるとき、その過程でエネルギーが放出される。
- エネルギーが何かから放出されるときはいつでも、そのエネルギーを捕獲して貯蔵しようとすれば、必ずいくらかのエネルギーが失われる。
これらの法則は、ポラックが提唱している排除層(EZ)のような、収穫可能な勾配が多数存在するような「フリーエネルギー」システムが存在する可能性を否定するために使われる。既存のパラダイムにとって不運なことに、生物学はしばしばこれらの法則に違反しているように見える。なぜなら、生物学は現在のパラダイムが予測する無秩序な状態ではなく、より秩序だった状態へと絶えず向かっているからだ。
(ノーベル賞を受賞した)このパラドックスに対する現在の解決策は、生物は「散逸的構造体」として機能し、環境から蓄積した大量の秩序成分中の秩序を交換する代わりに、自身の無秩序な内容物に秩序を与えるというものである。これはある程度、既存のパラダイムを維持することを可能にしているが、水はその液晶構造によって、通過する放射エネルギーを蓄積し、そのエネルギーを身体が利用できる秩序に変換する能力を持っているため、完全に正確であるとは思えない。
ナトリウム・カリウム勾配
生きている細胞でよく知られていることのひとつは、カリウムを細胞内に濃縮し、ナトリウムを排出することである。細胞内と細胞外では濃度が異なるため、勾配が存在し、物理学の既存の法則に従えば、勾配は均等化され、急速に消滅するはずだ。
そうならないため、既存のモデルでは、細胞膜がナトリウムとカリウムのすべてではないが大半の通過を阻止し(それによって勾配が均等化するのを阻害している)、細胞膜上にナトリウム・カリウムポンプが存在し、細胞内のナトリウムと細胞外のカリウムを絶えず交換していると主張してきた。この交換は細胞の健康維持に極めて重要であるため、細胞生物学ではナトリウム・カリウム交換ポンプの重要性に大きな焦点が当てられている。
残念なことに、このモデルには3つの根本的な問題がある(それぞれの証拠とそれ以上のことは、ギルバート・リンがここで紹介している):
- 既存のナトリウム・カリウムポンプには、ナトリウムとカリウムの勾配が自然に元に戻るのを打ち消すだけの能力がない。例えば、筋肉細胞では、ポンプを使ってナトリウム・カリウム勾配を維持するには、細胞内で利用可能な総エネルギーの4倍から30倍を必要とする。
- 細胞は、排出する他の様々な望ましくない成分で勾配を維持することができる(例えば、抗生物質を排出するバクテリア)。これらの現象を説明するために、より多くのポンプが同定され、モデルをサポートしようとしている。これは問題である。というのも、これほど多くの異なるポンプを同時に維持する能力を細胞が持っているとは考えにくいからだ。
(Na-カリウムポンプを作動させるのに必要な)エネルギーが完全に欠乏するように環境を変化させたカエルの筋肉細胞を用いた。にもかかわらず、勾配は維持された。
では、何がこの勾配を作り出しているのだろうか?リンは次のような観察を行った:
- (ポンプが存在する)細胞膜を除去しても勾配は維持された。
- 細胞質(細胞の内部)を含まないナトリウム・カリウムポンプのある膜を作ると、勾配は膜とともに急速に消失した。
このことは、勾配が細胞膜ではなく細胞質の特性であることを示唆している。興味深いことに、同じ性質が人工ゲルでも見つかっている。人工ゲルも細胞質と同様に、多量の液晶水を生成する:
私たちは、ゲルバリアが、イオン強度と化学組成の異なるイオン溶液を、ポンプ作用なしに安定に分離維持できることを示した。Na+/K+濃度勾配がバリアを横切って持続する場合、K+が豊富な側では負の電位が発生する。この状況は細胞内を彷彿とさせる。さらに、ゲルの細孔が大きく、浸透圧または静水圧勾配があるにもかかわらず、ゲル障壁を横切る水分子の移流も制限される。
この濃縮現象は、他の著者によっても観察されている:
このような蓄積凝縮の証拠は、1932年にBungenberg de Jongが、ポリマーの希薄溶液であっても振ると合体して飛沫になり、有機物が高濃度になることを示したときに、ほぼ1世紀前に得られている。このような飛沫のポリマー濃度は、周囲の浴の濃度を1万倍も上回ることがあった。ある種の染料の中に入れると、飛沫は次第に着色し、その濃度は周囲の溶液の何倍にもなることがよくあった。このように、ゲル飛沫は、細胞がカリウムを濃縮するように、ある種の溶質を濃縮する能力を持っていた。
ゲルは細胞の不可解な特徴の多くを単独で示すので、生物学全体で観察されるナトリウム・カリウム勾配を作り出すメカニズムを共有している可能性があると結論づけるのは妥当である。
障壁:
全ての細胞にとって大きな課題の一つは、不要なものを排除することである。古典的には、病原性物質が細胞内に侵入する過程は、細胞上のレセプターと相互作用して生じるものと考えられている(この世界観は、この過程を阻害する薬が開発される可能性があるため、医学によって強調されていると私は考えている)。私は、細胞によるこの排除プロセスは、既存の電荷、ゼータ電位、液晶水に大きく影響されると考えている。
液晶水はほとんどの場合、マイナスに帯電した表面でしか形成されないため(水分子がそれぞれ水素原子を半分排出し、それによって失われた水素によって正味のマイナス電荷を保持するのと同じ)、体内(および他の多くの自然システム)のほとんどすべての表面はマイナス電荷でコーティングされている。コロイドの安定性(ほとんどの流体系の挙動を支配している)は、同一の電荷によって生じる相互反発に大きく依存しているため、ほとんどの自然のコロイド系は負の電荷を利用して分散状態を保ち、過剰な正の電荷が存在する場合にのみ凝集する(同様に、ゼータ電位を復元するほとんどの方法もこの原理に基づいている)。
病原性生物は、しばしばプラス電荷を環境に与える(例えば、バクテリアはタンパク質内のアミノ酸のマイナス電荷を脱炭酸する)。これにより、体内のタンパク質や細胞は凝集し始め(マラリア寄生虫は特にこれが得意である)、免疫系が微生物に接近するのを妨げると同時に、微生物が細胞に接近する能力を増大させる。
生理学的ゼータ電位の最初の提唱者によってなされた特に興味深い観察のひとつは、細菌やウイルス感染(これはインフルエンザで最も研究されたと思う)が、生理学的ゼータ電位の一貫した劣化を引き起こすというものであった。これは、インフルエンザのようなウイルスが高齢者にとって非常に危険である理由の説明となる。健康なゼータ電位を持っている人は、コロイドの安定性が多少損なわれても耐えることができるが(少し気分が悪くなるだけである)、すでにゼータ電位が損なわれている人(これは老化の非常に一般的な結果である)にとっては、その低下は重篤な病気が発症する臨界閾値を通過するのに十分である。
COVID-19の特徴的な特徴のひとつは、多くの患者が重度のゼータ電位障害を示唆する臨床症状を示したことである。ワクチンが市場に出回るずっと前から、私がスパイクプロテインを懸念していた主な理由は、このゼータ電位破壊の犯人である可能性が高いと同定していたからだ。これはSARS-CoV-2ウイルスの新規成分(SARS-CoV-1とは異なる)であり、ウイルス表面に存在し、異常に高い正電荷密度を持っていたからだ。その後、スパイクプロテインが赤血球の凝集を直接誘導することが明らかになったが、これはおそらくその電荷分布が生理的ゼータ電位を著しく乱す可能性があるためであろう。
液晶水の体内生産を改善するためのプロトコルを受けた人と、ゼータ電位を改善するために同じことをした人の両方が、「病気になることはほとんどない」と述べているのを、私は繰り返し見てきた。これは、これらの生理学的パラメータがすでに良好な健康状態であれば(例えば、臨界ゼータ電位のしきい値を通過しないので)、感染症の症状がはるかに軽微であることを意味するか、あるいは病原体によって細胞が感染するのを直接的に防ぐためである可能性がある。どちらか一方のパラメータにプラスの影響を与える多くの要因が、もう一方のパラメータにも影響を与えることが多いため(例えば、アーシングはゼータ電位を改善し、体内の液晶水の成長を促進する)、最終的にどちらの効果が優勢かを言うのは少し難しく、このトピックに関する私の研究の大部分は、この疑問に答えようとすることに基づいている。
注:ゼータ電位を回復させ、体内の液晶水を増やす方法については、ここでさらに論じる。
この記事では、そのバリア仮説を支持する証拠のいくつかをレビューする。私が出会った最も古いものは、ヴィクトール・シャウベルガー(1885-1958)の研究によるもので、彼は、水が渦を巻くようなパターンで移動することが決定的に重要だと考えており、このシリーズの前編で述べたように、液晶水の存在を増加させることが示されている。彼の設計に基づく渦流システムを水道水に利用することで得られた多くの発見のひとつは、水道システム内(パイプの内側など)でのバクテリアの繁殖を抑えることができたことである。
この研究のように、液晶水の存在がバクテリアの増殖や侵入を防ぐことを直接的に発見した研究もある:
大腸菌を使ったテストでは、細胞は2日間かけて徐々にEZ[液晶水]を伝染した。さらに、EZを含むナフィオンは、EZを含まない親水性の平滑な酸化アルミニウム表面と比較して、2日間にわたる大腸菌のバイオマス蓄積量が80%少なかった。このことは、EZが特定の親水性表面に近づく細菌に対して、空間的にも時間的にも防御の第一線を担っている可能性を示唆している。Qこれらの知見は、バクテリアの付着やバイオフィルム形成に敏感な用途向けに、バイオファウリング抵抗性材料表面を開発する上で重要な意味を持つ可能性がある。
そしてこの研究:
絹セリシンは球状タンパク質であり、その耐ファウリング性は生体材料や浄水膜への応用において重要である。ここでは、セリシンがどのようにして水排除層(液晶性)を形成し、防汚挙動を促進するかが示されている。バクテリアの表面電荷と疎水性ドメインを模倣するため、負電荷を帯びた微小球が用いられた。水に浸したセリシンは、2pH単位以上の減少を伴うプロトン勾配とともに、100μmサイズの排除層(ミクロンサイズの汚濁物質に対して)を形成した。このように、セリシンと接触すると、表面付近の水分子が再構築され、生物付着を防ぐ物理的排除バリアを形成する可能性がある。pHの低下により、水性媒体は好中性細菌にとって生存不可能なものとなる。したがって、バイオファウリングに対する抵抗性は、他の要因の中でも、水の排除現象に基づいて説明できると思われる。
多くの組織は、液晶性の水が組織を覆っていることに依存しており、これは少なくとも部分的には、液晶性の水が作り出す保護バリアによるものであろう。例えば、1970年のGreenとOtoriは、角膜とコンタクトレンズから約350μmの深さの排除層を発見した(眼を損傷から守ることは重要であり、液晶水は透明で潤滑性の高いバリアとして機能する) ポラックは約15年前にも、コンタクトレンズの製造に使われるゲルであるポリニパムの隣に同じ結果(液晶水の存在)を発見している。
体内で最も重要な組織のひとつは、血管の内皮 (しかしよく潤滑な層である)である。そこでは、大きな剪断力を生み出す大量の血液が常に血管の上を通過している。この点について、マルコム・ケンドリックは、最も大きな剪断力を経験する循環部位が、最終的に心臓病につながる慢性的な損傷を発症する可能性が最も高いと指摘している。
内皮の保護膜であるグリコカリックスは、(硫酸塩を多く含むため)周囲に大量のゲル(液晶性の水)を作り、内皮を保護する構造になっている。さらに、血液細胞が入り込めない薄い層が、どんなに細い血管の壁にも直接並んでいることが知られている。同様に 2000年の研究者たちは、健康な毛細血管(体内で最も細い血管)の内皮を覆う厚さ0.4~0.5μmのゲル化した水の層を観察しており、この内皮は様々な異なる物質が内皮に到達するのを排除している。
公式には、ワクチン・スパイクプロテインは機能的なACE2結合ドメインを持たないため、内皮に影響を与えることはできない(一方、COVID-19ウイルスは機能的なドメインを持ち、そこから細胞内に侵入する)。これは事実だが、スパイクプロテインが内皮に結合する方法は他にもたくさんある(例えば、スパイクプロテインがレセプターに結合すること、糖鎖のヘパリンと直接結合すること、ワクチンによって内皮の内側から産生された場合、事実上内皮に入ったことになる)。ワクチン接種による死亡が疑われる人物の剖検で、内皮層に深刻なダメージがあることが判明していることから、スパイクプロテインが何らかの形で、通常は保護されているバリアーに侵入する能力を持っていることを示す強力な証拠となる。
これは、スパイクプロテイン(および脂質ナノ粒子のプラス電荷を帯びた成分)が帯びるプラス電荷が、内皮のような細胞を保護する液晶性の水バリアに侵入するためではないかと多くの人が考えている。血液粘度の上昇に関連する他の状態(例えば、コントロールされていない糖尿病)は、内皮にダメージを与え、心血管系疾患への進行を促進することが知られているので、スパイクプロテインのゼータ電位破壊によるものとも考えられる。
相転移:
液晶水は生命維持に必要だが、多すぎるのも問題である。例えば、細胞全体がゲル状態の水で覆われていたら、細胞内に入る必要のある多くのものが入ることができなくなる。この問題を解決するために、膜にはプラス電荷の領域が存在し(例えば、ポラックは、これが細胞膜内に金属イオンが配置される重要な理由であると主張している)、その領域上に液晶水が形成されるのを防ぎ、その結果、水が移動するための開いた通路ができる。
注:私たちは細胞を、リン脂質とあちこちにあるタンパク質からなる流動性のモザイクと考えるように教えられているが、細胞膜の約50%は通常タンパク質で構成されている(さらに多い場合もある)。
しかしほとんどの場合、身体は必要に応じて水をゲル(液晶)状態とゾル(コロイド懸濁液を含むバルク水)状態の間で変化させることができることに依存している。しかし、細胞生物学における相変化の基本的重要性は、過去70年の間に徐々に認識されるようになったものの、科学分野の大部分は、細胞膜は常に液体であるという流体モザイク仮説に固執している。
簡単に説明すると、ゲル(天然のものも合成のものも)には通常、ある状態から他の状態へ急速に転移する温度(転移点)がある(ここでさらに詳しく説明する)。私の知る限り、どの場合も、高温の方がゾル状態(コロイドが浮遊したH2O水)に対応し、低温の方がゲル状態(H3O2)に対応する。さらに、圧力の上昇もゲル状態を促進する:
生物学的活動は、ほとんど普遍的に、膜が流動状態にあることを必要とする;
多くの場合、遷移点のすぐ上にある
全身のゲルは、相転移点が体温付近になるように設計されている。これは、ほとんどの哺乳類や鳥類が体温を35℃~42℃(95°~107.6°F、人間の平熱は37℃または98.6°F)の間に保っている理由を説明するものだろうが、彼らの体や環境がどれほど異なるかを考えると(例えば、砂漠の小さな鳥とシロナガスクジラ)、そうでなければ不思議な事実である。さらに、この温度は温血動物にとどまらない。マグロ科の魚は活動時、体温を30℃(華氏86度)に保つが(海はもっと冷たい)、爬虫類の多くは体温を40℃(華氏104度)近くに保つ(太陽を探すなどして)。昆虫の多くも、同じような温度範囲(例えばマルハナバチの場合、38℃または100.4°F)に温まるまで飛ぶことができない。
同様に、相転移点はゲルの材料(細胞膜の成分など)に大きく影響される。冷血動物もまた、環境に応じて細胞膜の組成を変化させることが観察されている。
ある実験では、カロテス・バーシカラー(一般的な庭用トカゲ)を16℃、26℃、36℃(60.8℃、78.8℃、96.8℃)に30日間順化させた。トカゲは、ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルイノシトール、カルジオリピン、スフィンゴミエリンといった主要なリン脂質(細胞膜の構成成分)の濃度に劇的な変化を示した。温度順化によって、コイの血球とともにコイの筋肉ミクロソームの脂質組成が変化することも観察されており、ニジマスの脳の脂質組成には季節変動が観察されている。
いったんゲルが転移温度付近に保たれれば、その転移温度を正確に変化させることは比較的容易である。強い陽イオン(プラスに帯電したイオン)はゲルを破壊し、強い陰イオン(マイナスに帯電したイオン)はゲルを強化するからだ。私が知る限り、これらのイオンの効果は、生理的なゼータ電位を最も効果的に支持する価数(電荷)の高い陰イオン(-)と、それを最も効果的に破壊する価数の高い陽イオン(+)に相関している。
ゲルを破壊するために、身体が転移温度をシフトさせるために使用する主な陽イオン元素は、カルシウム(Ca)である。カルシウムの効果を図式化する(Ca濃度が突然臨界閾値に達することに注意):
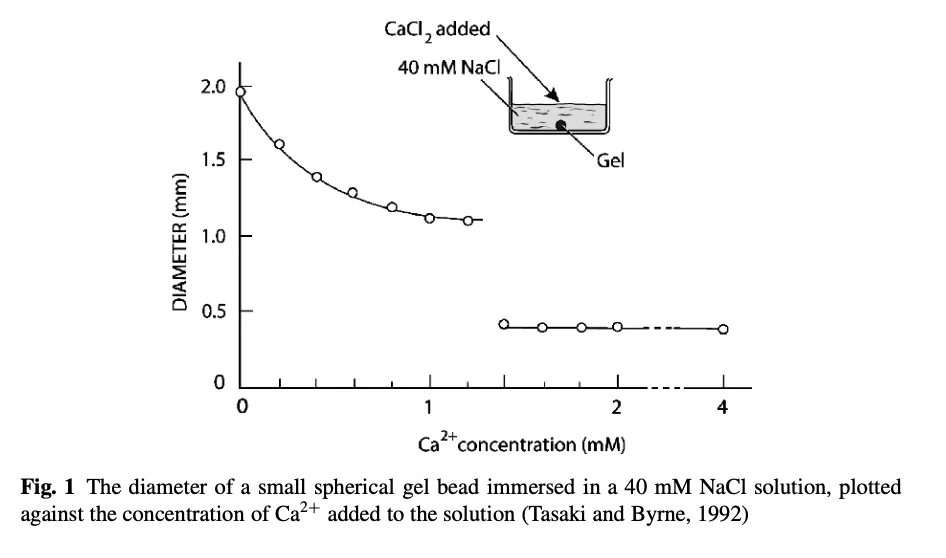
これらの結果は、価数の高い陽イオンであるCa2+が、価数の低い陽イオンであるNa(1)+に比べて、いかに小さなゲルを形成するかも示している。
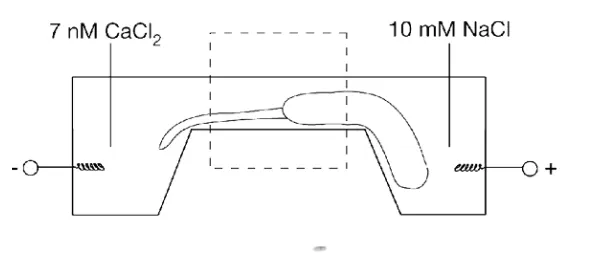
これらのことをまとめると、事実上、細胞内でこのような状況が起こり、そこでカルシウムの小分泌物がその水分を流動状態からゲル状態に移行させることができる。
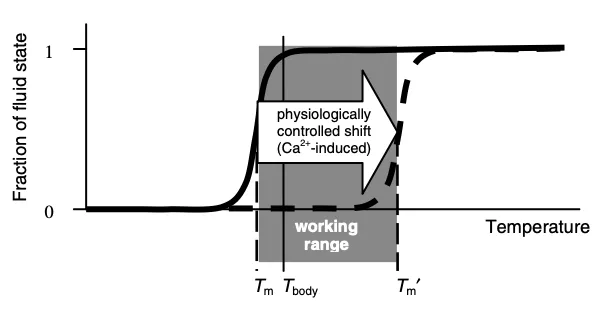
逆に言えば、もし体がゲルを分散させることができるのであれば、それを再現する方法も必要になる。ゲルを作るために使われる主な強い(原子価の高い)陰イオンは、アデノシン三リン酸(ATP)のリン酸塩である。
このATPの代替機能を考える際には、リンによって最初に提唱された考え方のいくつかを考慮する必要がある(178ページの8.4参照)。彼らは、ATPから得られるエネルギーの多くは、ATPの結合が切断されることによるものではなく、ATPが水を誘導して液晶構造を形成させることによるものであることを示唆している。要するに、ATPから発生するエネルギーの多くは、最終的にこれらの重合(ゲル化)の引き金となる放射エネルギーから生じる。
相転移は細胞生物学ではごく一般的な現象であり、この分野ではその重要性はあまり強調されていないが、ポラックの最初の2冊の本の中心的主張は、相転移は細胞の多くの機能を説明するために必要であるというものである。
注:後にポラックと私の論文について議論した際、彼は、彼の初期の研究における主な見落としは、ゲルが負に帯電していることをまだ知らなかったことだと教えてくれた。
膨張するゲル:
体内でのゲルの主な用途の一つは、ゲルが急速に膨張し、その膨張から効果を生み出すことである。例えば、細胞は重要な成分を含む小さな球体(小胞として知られる)を頻繁に排泄するが、この小胞は細胞を出ると急速に膨張する。例えば、単離されたムチン産生血管は、40ミリ秒で600倍に膨張する。
細胞から出る前に、小胞の内部成分はCa2+によって架橋される(ゲルタンパク質を架橋することにより、価数の高い陽イオンはゲル形成を制限し、ナトリウムのような一価陽イオンはゲルを膨張させる傾向があることを覚えておこう)。細胞から分泌されると、Ca2+は除去され、ゲルは各細胞を取り囲むNa(1)+溶液中で急速に水和する。逆に、ゲルは蒸留水中では急速に膨張せず、脱イオン水中では液晶水の形成も減少することが観察されている。
注:小胞の中には、局所の環境因子を利用して相を変えるものがある。例えば、ヒスタミンは中性のpHでは一価の陽イオン(電荷は+1)だが、酸性のpHでは二価の陽イオン(電荷は+2)になる。研究者たちは、ヒスタミンが酸性環境では(価数が高いため)マスト細胞顆粒を収縮させ、一方、中性環境ではマスト細胞顆粒を膨張させることを示すことができた。
筋肉生理学:
患者が医師を求める最も一般的な問題の多くは、筋肉に起因している。そのため、私はこれらの問題を治療するためのさまざまな方法を研究してきた。その結果、一貫して効果のある方法をいくつか見つけたが、筋肉生理学の既存のモデルでは説明がつかないため、その理由を説明することができない。
医学(そしてしばしば科学)の大きな問題のひとつに、私が「機械論的な罠」と呼んでいるものがある。簡単に言うと、何かがなぜ効くのかを説明するメカニズムがない場合、それはデマであり、実際には効かないと思われてしまうということだ。特に、薬が効く理由を説明するために利用するモデルの多くは、実際には意味をなさないのだから。同様に、ある薬物のメカニズムについて以前受け入れられていたモデルが破棄され、別のモデルに取って代わられた例はたくさんあるし、ある治療法のメカニズムについて、正統派でないモデルの方がはるかに優れた説明を提供しているように見える例を、私はもっとたくさん知っている。
注:これは多くの薬物以外の代替療法にも当てはまる。
残念なことに、生化学的なメカニズムが提案されない限り、FDA(あるいは従来の訓練を受けた多くの医師)がその療法を研究対象として検討することはほぼ不可能である。これは、医薬品が体内の特定の酵素を標的として設計されているため、私たちの厳格なメカニズムパラダイムに適合する一方、幅広い作用スペクトルを持つ治療法は、特定の生化学的受容体に関連付けることができず、利益を得ることも容易ではないという事実の反映だと私は考えている。
このようなことを考えると、私はポラックを通して、既存の筋生理学モデルの多くの側面が意味をなさないこと、そして筋肉に関する厳密な実験を行ったときにモデルの期待する結果が得られないことを発見し、大喜びした:
しかし、筋生理学の代替モデルも存在する。それを考えるとき、筋肉は収縮することしかできないと認識されていることに留意すべきである。身体のどこかを長くする必要がある場合、適切な場所にある筋肉を短縮させ、その位置関係から、そうすることで別の場所を長くすることで達成される(例えば、腕の上腕三頭筋)。
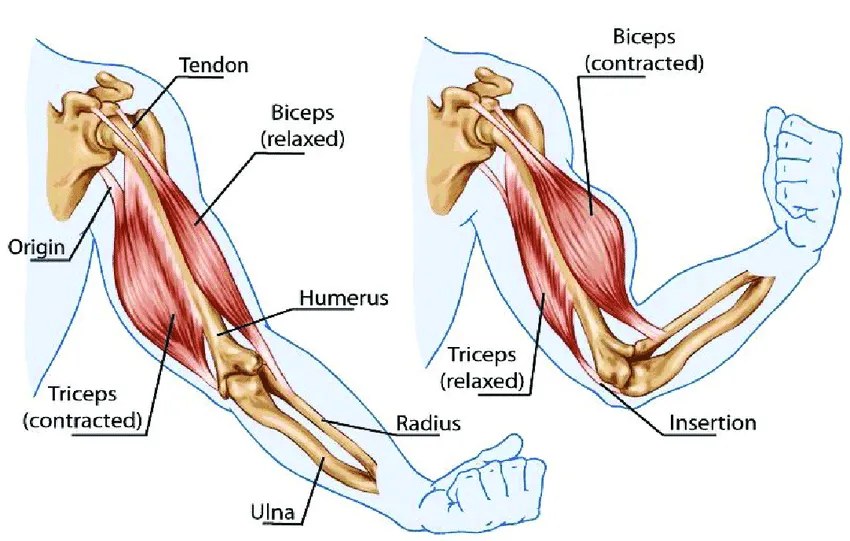
ポラックの研究から、筋肉は収縮時にしか力を生み出せないが、実はこれは、その構成要素が力強く伸びることしかできないことの結果であることに気づいた。
ポラックのモデルは、弛緩した筋肉には構造化された水が多量に含まれているのに対し、収縮した筋肉には主に半固体のゲル状態ではない水が含まれているという観察に基づいている。筋の発火は、カルシウム(Ca2+)イオンが筋に入るのを指示する神経信号に従うことが知られているので、このことは、カルシウムが誘発する相変化がこのプロセスの根底にある可能性を示唆している。
逆に、切り開かれた筋細胞は、塩溶液中ではカリウムとナトリウムの勾配を保つことができるが、ATPが欠乏している部分ではそれができないことも(勾配の項で前述した実験から)知られている。このことは、ATPが水の液晶状態を維持するために必要であり、ナトリウムイオンが筋細胞内に侵入するのを防いでいることを示唆している。同様に、ATPがないと、筋肉は収縮した状態のまま動かなくなり、伸びることができなくなることも知られている(「死後硬直」と呼ばれる)。
注:私が長年にわたって聞いてきた説得力のある逸話のひとつに、ワクチンで重篤な障害を負った子供たち(例えば、生涯衰弱し続ける自閉症の子供たち)が死亡した場合、死後硬直がはるかに早く進行するというものがある。これを観察した人たちは、これらの子供たちは慢性的なエネルギー不足に陥っており、彼らの身体は生きていくのに十分なATPを作るのに苦労している、と示唆した。これは非常にあり得ることだと思うが、ゼータ電位が低いことを示している可能性もある。これは他の慢性的な衰弱性疾患でも見られるのではないかと私は考えているが、もし実際に起きているのであれば、それを観察したであろう人々とは交流がない。
ポラックはまた、環境要因に反応した筋肉の変化が、おなじみの臨界閾値を超えた後に起こることも観察した。このこともポラックには、筋肉内で相変化が起こっていることを示唆しているように思えた。
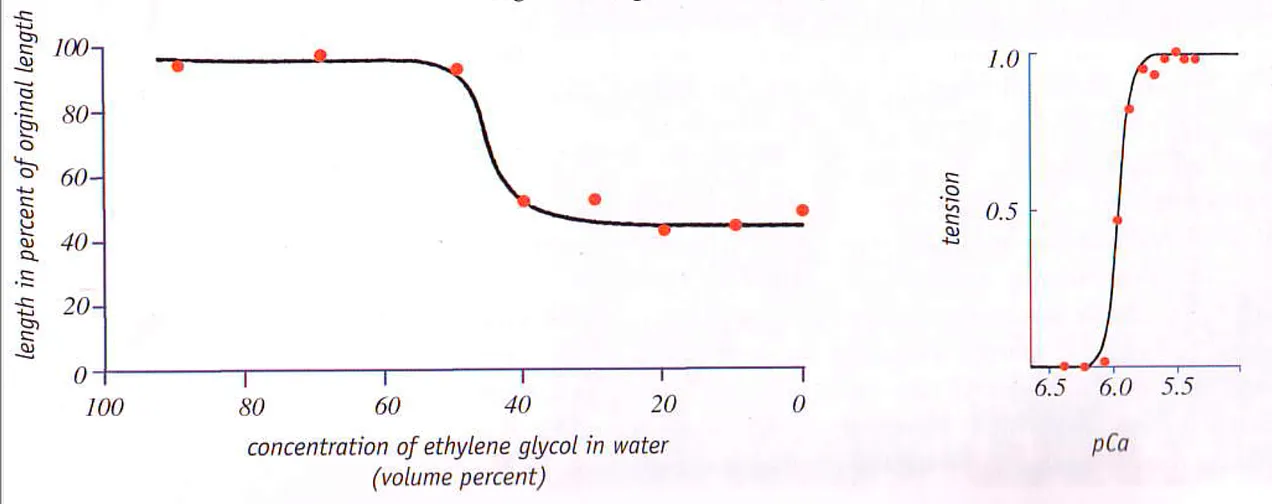
では、ポラックのモデルとは何か?
筋繊維は周囲に大きなゲルを形成するように設計されており、その中の液晶水が膨張すると、繊維内のコイル状でないタンパク質をらせん状に引き伸ばす(タンパク質の架橋がそれ以上の引き伸ばしを妨げるまで)。筋肉を収縮させる必要が生じると、カルシウムイオンが筋肉内に入り込み、ゲルが除去され、伸長されなくなったタンパク質は弾性的に元のコイル状でない状態に戻る。ゲルを再構築するためには、カルシウムによって作られた架橋を切断し、それによってゲルを再構築する位相シフトを引き起こすATPが必要である。
このように、筋肉の強さは、液晶水が放射エネルギーを貯蔵することによって生み出されたポテンシャル・エネルギーが放出されることによって生み出されるものなのだ。言い方を変えれば、筋肉の強さは、実は筋肉が自発的に伸びる能力にあり、私が出会った多くの効果的な筋肉治療法が機能する理由を説明している。
大きな機械的剪断によって、細胞骨格マトリックス[相変化に応じて膨張・収縮するもうひとつの重要な構造]が流動化するが、これはSGM[ある種の合成素材]の物理的若返りに似た現象である。この流動化に続いて、機械的特性がゆっくりとスケールフリーで回復していくが、これはSGMにおける物理的老化に似た現象である。驚くべきことに、一過性の伸張に応答して、細胞骨格は異なる細胞種に普遍的なパターンで流動化する。この発見は、通常想定されているような特定のシグナル伝達経路によってではなく、むしろ(以下に説明するように)物理的な力による非特異的な作用によって媒介されるメカニズムを示唆している。
他の多くの構造物も、その内部ゲルが膨張と収縮を引き起こす能力を持っているが、骨格筋とは異なり、ほとんどの場合、この膨張と収縮は、高度に架橋された骨格筋内で観察されるような強固な拘束は受けていない。平滑筋、収縮リング、合成アクチンゲル(アクチンは体内で最も一般的な収縮タンパク質のひとつである)、共有結合の架橋が少なく半ランダムな構造を持つ他の小器官なども、体が生命を維持するためにこの原理を利用している場所の例だ。
最後に、他の繊維(例えばコラーゲン)の周囲の塩分濃度を変えることで、筋繊維と同様の収縮をもたらす力を発生させることができるところまで膨張・収縮させることができることも観察されている。
結論
液晶水を探求する中で、いくつかの中心的な疑問を解決しようとしてきた。このシリーズでは、多くの伝統的な医療システムにおいて、身体全体に内在する膨張力が存在し、それが活力を与える上で重要な役割を果たしていること、そしてこの膨張力が損なわれると様々な病気が発症することを説明するメカニズムを探った。
私はこの考え方に同意し、生命を維持するための広がりが、窮屈な空間に置き換えられていることを認識できるようになった例を数多く見てきた。同様に、その広がりを回復させようとする方法が、患者の活力を回復させることも見てきた。逆に、加齢に伴う可動性と耐久性の低下(そして高齢の患者にとって最大の懸念事項のひとつ)は、加齢とともに一般的に経験する液晶水の喪失に直接関係していると私は考えている。
この記事では、体内における液晶水の構造的効果を図解してみたが、そうすることで、膨張する力がどこから生まれるのかを説明するメカニズムを提供できたのではないかと期待している。このシリーズの次のパートでは、水の液晶状態がどのように勾配を作り出し、それがエネルギーとして収穫され、生命が依存する循環を生み出すかを説明する。その次の記事では、液晶水とゼータ電位の絡み合った関係を説明することに全力を尽くし、このシリーズの最終回では、私が出会った、どちらかを回復させるように見える方法について論じた。
これらのトピックはそれぞれ非常に複雑であるが(例えば、この記事は多くの本や論文を総合したものである)、それでも生命の謎を解明するためには、飛び込んでみることが重要であると私は考えている。また、COVID-19の結果、私たちの社会に広く起きた覚醒が、このような型破りなテーマを探求する好奇心を再び呼び起こすことを願ってやまない。
この比較的複雑なテーマを理解するために、私にとっては身近で大切なことであるにもかかわらず、各位が注意を払ってくださったことに感謝したい。あなたにも、その意味するところのいくつかを日常生活に取り入れていただけることを心から願っている。
