
コンテンツ
Reviewing the evidence for viruses as environmental risk factors for ALS: A new perspective
https://pubmed.ncbi.nlm.nih.gov/29684753/
2018年2月13日受領;2018年4月5日修正版受領;2018年4月7日受理
ダニエル・B・セレステ、マシュー・S・ミラー⁎
Michael G. DeGroote Institute for Infectious Disease Research, McMaster Immunology Research Centre, Department of Biochemistry and Biomedical Sciences, McMaster University, Hamilton, ON, Canada
要約
筋萎縮性側索硬化症は壊滅的な神経変性疾患であり,その病因はいまだによくわかっていない。疾患の遺伝的基盤はごく一部の症例でしか知られていないため、環境因子がALSの引き金となっているかどうかに大きな関心が寄せられている。ウイルスは、ALSの誘因となりうるものとして長年注目されてきた。しかし、既存の研究では、因果関係を示す説得力のある証拠は得られていない。本レビューでは、ALSに焦点を当て、ウイルス感染と運動ニューロン疾患との関連性を裏付ける証拠をまとめている。先行研究の限界が議論され、その背景が明らかにされた。また、病気の発症にウイルスが関与していることを示す、より強力なメカニズムの証拠を示した最近の研究が紹介された。最後に、ALSとウイルスの関連性について新たな視点を提示し、この分野を将来的に前進させるためには、神経学と感染症の研究を橋渡しする学際的なアプローチが必要であることを強調した。
1. はじめに
筋萎縮性側索硬化症(ALS)は,最も頻度の高い運動ニューロン疾患(MND)であり,生涯発症率は約400分の1である[1]。筋萎縮性側索硬化症(ALS)は、運動ニューロン疾患(MND)の中で最も頻度の高い疾患であり、生涯発症率は約400人に1人である[1]。ALSの発症は、一般的に50歳から75歳の間に起こり、患者は診断から3~4年以内に亡くなることが多い [1]。ALSの病態生理には、いくつかの細胞および分子の異常な経路が関与していると考えられているが、発症・進行の原因となる特徴については、依然として議論の余地があり、完全には解明されていない[2]。現在までに、100以上の遺伝子の変異が、潜在的なALSの危険因子として同定されている。しかし、これらの遺伝子のうち、正式なALS遺伝子と考えられるほど詳細に研究されているのは、ごく一部にすぎない[3]。一部の集団では、ALSの遺伝的基盤が10%程度しか確認されていないにもかかわらず、双子をベースにした遺伝率の研究では、38〜78%の患者に遺伝的関連があることが裏付けられている[3,4]。しかし、ALS患者のうち、一般的な遺伝子変異に起因するものは11〜25%にすぎず[4,5]、したがって、希少な新規変異や遺伝子-環境相互作用が病気の病因に重要な役割を果たしていると考えられている。
長年にわたってALSとの関連性が調査され、議論を呼んでいる環境リスク要因の一つに、ウイルス感染がある[6]。歴史的に見て、この分野で最も広く研究されてきたウイルスは、エンテロウイルス(ポリオウイルスなど)レトロウイルス、ヘルペスウイルスなど、神経栄養学的な性質を持つウイルスである。しかし、これらの研究は、対照集団と比較して、ALSコホートの病理標本におけるウイルス抗原の陽性率および/または検出率の上昇を示す観察データに限られていた。このような研究は、仮説を立てるのには有用であるが、ある感染症の罹患率の上昇がALSの原因なのか結果なのかという重要な問題を解決することはできない。その結果、ウイルス感染がどのように病気のリスクを高めるかを説明する有力なメカニズムデータはほとんど存在しない。
上述の研究では、あるウイルスがALS患者の間で検出される率が高いことで、そのウイルスが病気を引き起こす原因になっているのではないかという仮定が中心になっている。しかし、これまでの研究がほとんど取り上げてこなかったより直接的な可能性は、一般的なウイルス感染が、ALSを発症する人の中ですでに調節されていない経路を妨害することによって病気を引き起こすということである。実際、ほとんどすべてのウイルスは、疾患の病態生理学的特徴として知られている経路の多くを刺激したり調節したりしている(図1)。酸化ストレスの誘発、タンパク質のフォールディング/ミスフォールディング経路の操作、ミトコンドリアの動態、ストレスグラニュール、炎症の誘発などは、すべてウイルス感染の一般的な結果である。遺伝的素因を持つ人がウイルスによってこれらの経路を阻害されると、病気の発症の引き金になる可能性があり、ALS患者の感染率が対照群に比べて高い必要はないと考えられる。
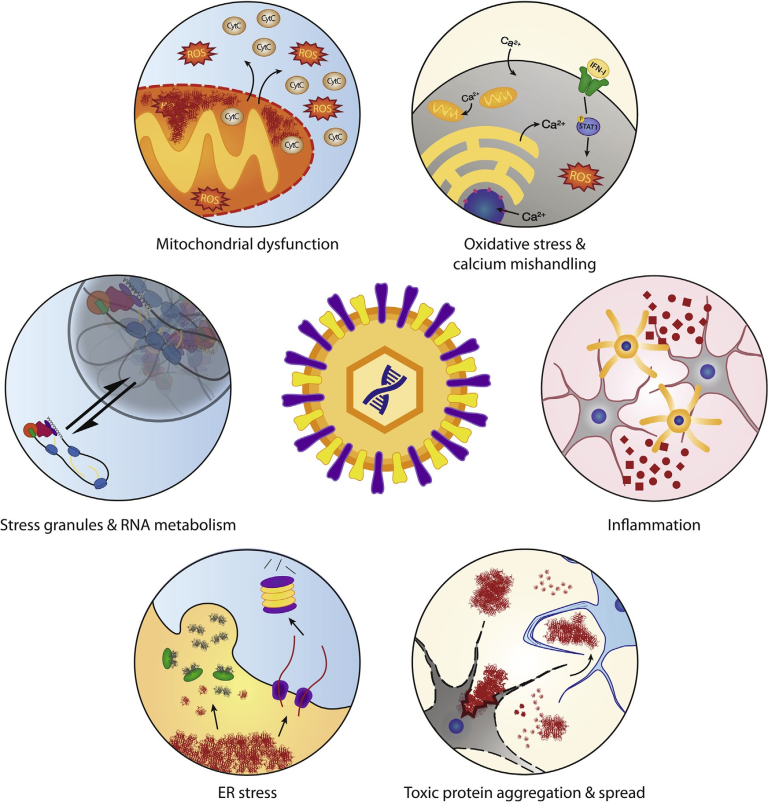
図1 ALSの病因に関わる細胞経路は、ウイルス感染時に操作される
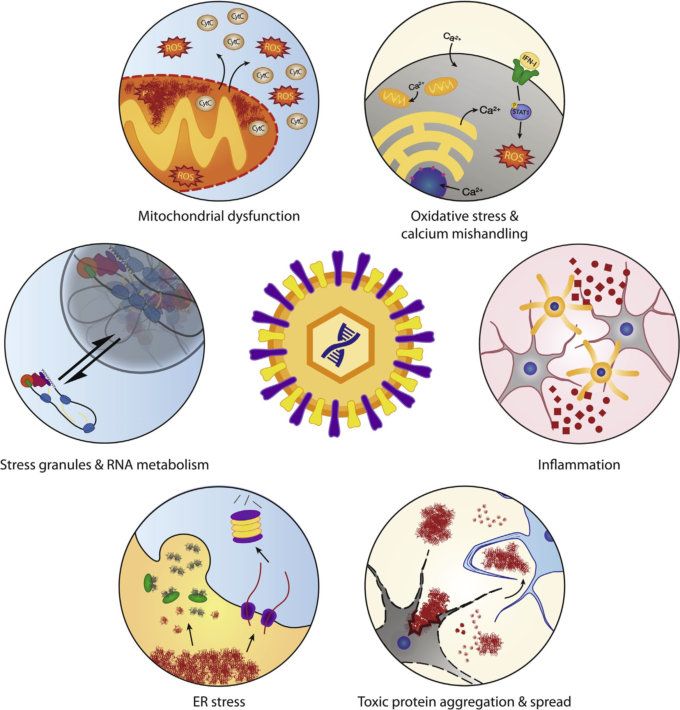
ウイルス感染は、酸化ストレス、炎症、毒性タンパク質の凝集と細胞間拡散、小胞体ストレス経路、ストレス顆粒の動態とRNA代謝、ミトコンドリア機能など、ALSの病態生理に関連する多くの経路に依存し、かつ/あるいは操作する。遺伝的要因によりすでにストレスを受けている経路にウイルスが侵入することで、ALS発症の引き金になったり、病気の進行を早めたりする可能性がある。
多発性硬化症(MS)患者は、感染後にしばしば再発/再燃を報告するが、ALSと診断された患者は、急性感染症と発症との関連性をあまり報告しない[7]。MSでの再発は、通常、感染に近接した場所で、すでに診断を受けた患者集団で起こる。対照的に、ALSの発症は非常にゆっくりと始まり、ALSを確定するための決定的な診断テストはない。初期症状を報告した後、ALSと診断されるまでには、1~2年の臨床評価期間が必要となることが多い。このように長い診断期間を経て、患者が「日常的な」感染症を発症と関連付けることは非常に困難である。重要なのは、急性感染症と症状発現の間にも遅れがあるということである。これらの要因により、感染症とALS発症の関連性を検出することは困難である。
以下のセクションでは、ALSのリスク因子としてのウイルス感染に関する文献の現状を要約し、議論する(表1)。そして、ウイルス感染が病気の引き金としてどのように作用するかを理解するための新たな枠組みを提供し、今後の研究で取り組むべき主要な未解決問題をいくつか取り上げる。
2. 報告されているウイルス感染とALSの関連性
2.1. ポリオウイルスとALSの歴史的関連性
ポリオウイルスの神経栄養学的性質と麻痺性小児麻痺を引き起こす性質(MNDと臨床病理学的性質を共有している)により、エンテロウイルスとMNDとの関係の可能性を調査する何十もの研究が行われてきた。ALSと過去のポリオウイルス感染との関係を調べた最初の疫学調査の一つでは、ALS患者のうち11人(127人中)が過去にポリオウイルスに感染していたという異常に高い割合を示した[8]。しかし、他の研究者は、これらのデータには統計的な誤りやALS患者の選択バイアスがかかっている可能性があると指摘している[9]。MNDにおけるウイルス感染の病因的役割の可能性を示唆する最初の疫学的報告は、若年者のポリオワクチン接種と約40年後のMNDによる死亡との間にわずかな相関関係があることを示している[10]。この報告は、イングランドとウェールズの人口レベルでの通知という形で集められたものであるため、相関関係にあり、因果関係を示唆するものではない。それにもかかわらず、著者らはポリオウイルスとMNDとの関係を明らかにすることに焦点を当てたさらなる研究の必要性を強調している。その後、MNDと過去のポリオウイルスとの関連性を評価するいくつかのケースコントロール研究が行われたが、有意な関連性は見出されなかった[11]。過去にポリオを患ったことのある人からのMNDの報告の多くは、現在我々が知っているポストポリオ症候群[11]と混同されている可能性があることは注目に値する。ポリオ経験者のMNDに対する遺伝的素因の可能性も報告されている。ポリオウイルス受容体タンパク質(PVR)のドメイン1に存在するAla68Thrのヘテロ接合型多型は、細胞培養において感染しやすくなることと関連しており[12]、進行性筋萎縮症の患者に多く見られるようである[13]。しかし、これらの変異は、対照集団と比較して、ALS患者では濃縮されていなかった。PVRの多型と進行性筋萎縮症との間の関連性が、感染感受性の増加によるものなのか、タンパク質活性の異常によるものなのか(PVRは胎児期の神経細胞分化に関与していることが報告されているため)は、不明である[14]。エンテロウイルスに感染すると、前角細胞が標的となるため、低位の運動ニューロンを含む急性の神経学的後遺症が生じることが知られている。ALSでは、上部および下部運動ニューロンが関与する。したがって、もしエンテロウイルス感染がALSの引き金となるのであれば、その特徴は急性感染時に起こるものとは異なるものでなければならない。これらを総合すると、ポリオウイルスとMNDの関係を支持する証拠は、因果関係を裏付ける機構的データが不足しているため、依然として論争の的となっている。そのため、ALSの臨床的特徴を再現し、ALSとポリオ後症候群を区別できる実験モデルを開発することが切実に求められている。このような研究は、感染症と病気の関連性について、より詳細なメカニズムの解明につながるだろう。
ALS患者の脊髄から検出されたエンテロウイルスのRNA配列が、エコーウイルス7と86~94%の相同性を示したことから、ALSとエコーウイルス7の感染との間に特異的な関連性があることも明らかになった[16]。しかし、その後のある研究では、ALSとエコーウイルス7の血清反応との関連性が報告されたが[17]、別の研究では、ALS患者の脳や脊髄からエコーウイルス7の配列を分離することができず[18]、この関連性の強さと広さに疑問が持たれている。
ALSの病因となるエンテロウイルスを広く調査している多くの研究では、ALS患者の中枢神経系(CNS)からのサンプルをRT-PCRで分析した後、様々な結果が得られている[16,18-21]。最近では、242名のALS患者と354名の対照群から採取した脳脊髄液を分析した結果、ALS患者のサンプルではエンテロウイルスRNAの頻度が著しく高いことが明らかになった[21]。しかし、ALS患者が脳脊髄液中にエンテロウイルス配列を保持している可能性が高い場合でも、この関連性が疾患の原因なのか結果なのかは不明である[22]。
2.2. ALSへのエンテロウイルスの関与を示す生化学的証拠
最近になって、ALSの発症にエンテロウイルスのプロテアーゼが関与していることを示唆する生化学的証拠が出てきた。それは、重要なALS関連タンパク質であるTDP-43を切断して、潜在的に病原性のある断片を生成することである[23]。Fungらは、Cox- sackievirus B3プロテアーゼ2AがTDP-43を核から細胞質に移動させ、そこでウイルスプロテアーゼ3CがTDP-43をそれぞれ約35kDaと約8kDaのN-末端とC-末端の断片に切断することを明らかにした[23]。安定したN末端側の断片は溶解性が低下し、ストレスグラニュールに局在してユビキチン化されたタンパク質の凝集体を形成した[23]-この表現型は、ALSの病的なTDP-43で観察されたものと同様である[24,25]。このデータは、エンテロウイルスが特定の疾患を引き起こす魅力的なメカニズムモデルを提供しているが、プロテアーゼ2Aで切断されたTDP-43がALSと関連しているかどうか、またどのような状況で関連しているかを明らかにするには、さらなる研究が必要である。
2.3. ALSと他の神経疾患におけるレトロウイルス
ヒト免疫不全ウイルス(HIV)に感染した患者の約70%が、通常は認知的な性質のものであるが、非医学的な合併症を経験している[26]。HIVやヒトTリンパトロピックウイルス(HTLV)に感染すると、ごく少数の患者がALSのような症候群に移行することから、外因性レトロウイルスがALSの発症に関与しているのではないかと考えられている。具体的には、HTLV-1感染者の0.25-3.8%が関連性脊髄症/熱帯性痙性側索硬化症(HAM/TSP)を発症し[27]、少なくとも35例のHAM/TSP症例がALS様症候群の診断基準を満たしている[28]。しかし、ALS発症のリスクとHTLV-1血清陽性率はいずれも60代で最も高くなるため、これらの関連性の信頼性を評価することは困難である[28]。さらに、HIVに関連したALS様症候群が少なくとも29例報告されている[28]。これらの症例は、一般的に、発症年齢が若く、進行が早く、脳脊髄液中のタンパク質濃度が高く、抗レトロウイルス療法を開始すると安定化または改善することを示している[28]。HIVは神経細胞ではなく、中枢神経系のミクログリアやマクロファージに感染するため[29]、これらの感染した免疫細胞が神経毒性のあるウイルスタンパク質やサイトカインを産生し、運動神経細胞を損傷する可能性がある[30]。また、まれに神経栄養性のHIV準種が個人の中で発生し、ALSのような症候群を引き起こす可能性も示唆されているが、現在のところその証拠はない[28]。
興味深いことに、マウスレトロウイルスがマウスのMNDを引き起こすことが知られている。特に、マウス白血病ウイルス(MuLV)は、マウスの中枢神経系に侵入し、おそらくenvを介したメカニズムで海綿状脳症を引き起こすことが示されている[31]。その後,トランスジェニックマウスの神経細胞にHIVの全ゲノムを侵入させると,神経疾患を引き起こすことが明らかになった[32].
2.4. HERV-Kの神経毒生成物
ALSの誘因として外因性レトロウイルスの役割を支持する証拠はまだ限られているが、現在、ヒト内因性レトロウイルス(HERV)がALSの病因において重要な役割を果たしていることを示唆する証拠が数多くある。 HERVとALSとの関係を詳しく調べるきっかけとなったのは、HIVおよびHTLV陰性のALS患者の血清中の逆転写酵素(RT)活性が、健常対照者と比較して予想外に高かったという報告である[33]。ALS患者の死後の脳組織を分析したところ、HERV-Kのenv、gag、polの発現が増加しており、さらにHERV-KのRTとTDP-43の発現および共局在が確認された[34,35]。興味深いことに、TDP-43はHERV-Kのロングターミナルリピートにも結合し、その発現を直接制御している[35]。ALSの発症にHERVが関与していることを裏付けるこれまでで最も説得力のある証拠として、Nath研究室が行った実験では、試験管内試験でenvの神経毒性が証明された[35]。 このデータは、envを発現しているトランスジェニックマウスが、上下の運動ニューロンの変性を起こすことからも裏付けられている[35]。
エンテロウイルスと同様に、ALS患者におけるHERV遺伝子の濃縮の観察が、病気の結果として得られたものなのか、それとも病気がHERVの発現の結果として得られたものなのかを明確に判断するには、さらなる研究が必要である。HERV-Kを介した毒性は、HIV感染による神経学的な症状に大きな役割を果たしていると考えられている。HERV-Kの発現は、HIV感染時に上昇し、抗レトロウイルス療法によって減少する可能性がある[36]。TDP-43の発現もHIV感染時に上昇するが、これにはリン酸化が伴っており、タンパク質本来の機能が失われている[36]-その一つがHERV-Kの発現の負の制御である[37]。したがって、HIV感染時のTDP-43の異常な発現と翻訳後修飾は、HERV-Kの発現を増加させ、その後の神経炎症を引き起こすと考えられる。HERV-K要素の発現とALS発症/進行の他の古典的マーカーとの間の時間的関係を明らかにすることを目的としたより詳細な研究は、困難ではあるが、この問題により多くの光を当てることができるであろう。
2.5. ALS患者におけるヘルペスウイルスの陽性率の上昇
ヒトヘルペスウイルス(HHV)もまた、神経変性疾患における役割の可能性について研究されている。アルツハイマー病における単純ヘルペスウイルス1型(HSV-1)の役割を支持する比較的強い証拠が蓄積されている[38]。しかし、ALSの発症においては、これらのウイルスがALSの発症に関与していることを裏付けるデータは非常に少ない。HHVがALSの危険因子として作用していることを示唆する実験的証拠は、ALS患者の間でHHV-6および-8の血清陽性率が上昇しているという単独の報告のみである[17,39]。しかし、HHV-6の再活性化は、様々な病気に反応して非特異的に起こるため、ALSとの関連性は非特異的なものである可能性が示唆されている[40]。とはいえ、これらの結果は実際の研究で検証されるべきである。なぜなら、異なるALSコホートを用いてこれらの結果を独立して再現すれば、証拠の質が高まり、より詳細な研究の正当性が得られるからである。
3. ALSとウイルスの関連についての新しい展望
かなりの努力が払われているにもかかわらず、これまでALSに関連するウイルスの数は比較的少なく、ほとんどの場合、疾患における因果関係を示唆する証拠はほとんど存在していない。それにもかかわらず、環境リスク要因としてのウイルスを研究することには説得力がある。ALS患者のコホートにおいて、特定のウイルス感染が濃縮されていることを示す強力なデータが少ないのは、ALS患者が発症前に特定のウイルスに対する感受性に健常者と比較して大きな違いを示さないことを反映しているのかもしれない。しかし、一般的なウイルス感染は、遺伝的素因によって機能不全に陥っている経路にストレスを与えることで、疾患を引き起こす可能性があるという未解明の可能性もある。ウイルスとALSの関連性を報告している既存の研究では、この可能性が見落とされている。このようなシナリオでは、ALSを発症する人としない人の間に、感染率の違いはないと考えられる。
我々や他の研究者は、以前、神経変性疾患には炎症が大きく関与していることを指摘し、ウイルスによって誘発された炎症が病気を引き起こし、永続させるのではないかと仮定した[41-43]。この仮説を裏付けるように,リポ多糖を用いて自然免疫系を慢性的に刺激すると,ALSのSOD1G37Rマウスモデルでは,病気の経過が早まることが示されている[44]。しかし、これまでに報告されているALSの病態生理学的特徴をより注意深く検討すると、病気に関与していると考えられている経路とウイルス感染との間には、はるかに広範な関連性があることがわかる。実際、ALSに関連することが知られている経路(図1に示す)の1つまたは複数を操作することは、ウイルス感染の普遍的な特性である。これらの経路をウイルスがどのように操作してALSに貢献するかについて、以下のセクションにまとめた。
3.1. ウイルス感染による小胞体ストレスの誘発
小胞体(ER)ストレスの誘導は、ALSや他の神経変性疾患でよく見られる[45]。SOD1の変異を特徴とするALS患者や関連するマウスモデルでは、粗面小胞体やゴルジ体の拡張を含む小胞体の形態異常が見られ、これはカルシウムのホメオスタシスの変化と一致している[46]。誤って折り畳まれたSOD1は,小胞体に蓄積し,小胞体関連タンパク質分解経路の構成要素を阻害し,重要なシャペロンタンパク質を隔離することが知られている[47].C9ORF72のリピート拡張や,ALSに関与するFUS(fused in sarcoma)やVAPB(vesicle-associated protein B)の変異も,アンフォールドタンパク質応答(UPR)の構成要素との相互作用を通じて小胞体ストレスを誘発し,最終的にプロテオスタシスを変化させることが報告されている[45]。神経細胞を含むある種の細胞は、小胞体ストレスに特に敏感で、アポトーシスを起こしやすいようである[48]。しかし、小胞体ストレスを受けた非神経細胞の死はストレスを受けた非神経細胞、特に神経細胞の健康に直接寄与する細胞(例:ミクログリア、アストロサイト)の死は、病気を永続させる可能性がある。
ウイルス感染は、小胞体に多大なストレスを与える。多くのウイルス(例えばエンテロウイルス)は、小胞体膜をハイジャックして、細胞質内に複製コンパートメントを生成する[49]。さらに、感染後期に高発現するウイルスの糖タンパク質は、宿主細胞のグリコシル化機構を過剰に刺激し、UPRを刺激するミスフォールドしたタンパク質の集積を引き起こす。このストレスが解消されなければ、細胞死に至る可能性がある[50]。小胞体はまた、ウイルスにコードされた膜チャネル(「ビロポリン」)によって頻繁に標的とされ、細胞内のカルシウムホメオスタシスを変化させ、しばしば感染細胞からのウイルス放出を効率的に引き起こす目的で使用される[51]。ALSの場合、タンパク質のミスフォールド(SOD1など)が病気の病因に重要な役割を果たしている可能性があるので、小胞体のこうしたウイルスによる変化は、特に問題になるかもしれない。ウイルスに感染した細胞にミスフォールドしたタンパク質が蓄積され、続いてそれらの細胞が溶解すると、中枢神経系以外の組織からでも、非細胞自律的にこれらの毒性分子の拡散が促進される可能性がある[52]。
3.2. ウイルスによるミトコンドリア機能の操作
ミトコンドリア機能障害は、ALSの特徴としてよく知られており、代謝、カルシウムホメオスタシス、活性酸素種、アポトーシス経路の変化がすべて報告されている[53]。これらのプロセスの制御におけるミトコンドリアの多面的な役割を考えると、ウイルスとミトコンドリアの広範な相互作用は驚くべきことではない。ミトコンドリア抗ウイルスシグナルタンパク質(MAVS)は、ウイルスの感染を感知すると活性化されるシグナルカスケードにおいて中心的な役割を果たし、複製を制限し、アポトーシスを刺激する抗ウイルス遺伝子の転写を引き起こす。したがって、多くのウイルスは、ミトコンドリアに局在することができるタンパク質をコードしており、抗ウイルスシグナルを制限したり、アポトーシスを調節したりすることができる[54]。また、ウイルスは、生産的な感染を維持するために必要なミトコンドリアの動態(分裂、融合、マイトファジーなど)を変化させる[55]。したがって、ウイルス感染時にミトコンドリアにかかる大きなストレスは、疾患の発症や進行を促進する可能性がある。
3.3. ウイルス感染時のストレスグラニュールの制御
ALSに関連する遺伝子の多くは、RNAメタボリズムやストレスグラニュール(SG)の制御に関与している。SGの形成は通常、細胞の生存を促進するが、ALSの場合、SGは異常なタンパク質の凝集部位として機能する可能性がある[56]。自身の再増殖を促進するために、すべてのウイルスは、細胞の翻訳機構をハイジャックする戦略を採用している。過去10年間で、SG経路と相互作用するウイルスの数は増え続けている[57]。一部のウイルス(HSV-1など)はSGを阻害しているようだが、他のウイルス(リ・スピラトリー・シンシチアル・ウイルスなど)は、ウイルスの複製を促進するためにSGを誘導している[58,59]。感染時にSGの形成が促進されると、ALSの文脈では、ミスフォールドしたタンパク質の凝集が促進されると予測される。しかし、SGの形成能力が低下すると、ALSで発生することが知られている他の多くの種類のストレスに細胞が耐えられなくなるため、SGを阻害するウイルスであっても、注意が必要である。ウイルスとSG経路との接点の複雑さは、特定のウイルス感染と特定のALS関連遺伝子との関係をより深く理解する必要があることを示している。
遺伝子を理解する必要がある。
4. ALSの誘因としてのウイルスに関する今後の研究の枠組み
ALSに関連することが知られている経路と、ウイルス感染時に操作される経路が広範囲に重複していることから、ウイルスが病気の経過に影響を与えるかどうかを明らかにするためには、より詳細な調査が必要であると考えられる。ウイルス感染は、広く普及しているため、研究する上で特に重要かつ困難なきっかけとなる。上述したように、既存の文献では上述したように、既存の文献では、特定のウイルスがALSの原因となることを、検出率の高さに基づいて支持する説得力はない。しかし、これらの研究では、ALSを発症する人としない人の間で同程度の割合で発生するウイルス感染が、ALSに罹患しやすい人では基礎的な遺伝子変異によってすでに脆弱になっている経路を撹乱することによって病気を引き起こすという、より直接的な可能性については言及されていない。上述の研究は、ある種のウイルスが他のウイルスよりも強力な誘因となる可能性を否定しているわけではないことに留意する必要がある。したがって、ALSモデルにおける感染の影響を明らかにするためには、より機構的なアプローチが必要である。
モデルシステムにおける還元主義的な研究は、ヒトにおける可能な環境的誘因の研究に関連する本質的な課題のために、特に重要である。ALSのほとんどの症例は散発的に発症し、疾患の遺伝的基盤は不明なことが多いため、前向きなコホートで環境誘因の影響を明らかにすることは非常に困難である。ALS集団と対照集団の暴露履歴の違いを明らかにすることは、事後解析により比較的容易にできる。しかし、同じ割合で発生する曝露が、診断前の疾患にどのような影響を与えるかを理解することは、非常に困難である。 ALSの場合、症状を最初に認識してから最終的にALSと診断されるまでの期間が比較的長いため、この課題はさらに大きくなる。さらに、症状が出る前にウイルスに感染した場合、特に風邪のような日常的な感染では、その影響をすぐに実感することはできないだろう。
ウイルス感染がALSに及ぼす影響を調べるためのより現実的な出発点は、すでに病気と診断された人に感染がどのような影響を与えるかに焦点を当てることであろう。この方法では、ALS患者の感染に対する生物学的反応を対照群と比較して評価することができる。また、病気の遺伝的基盤がわかっている場合、ウイルスが病気を悪化させる能力は一部の遺伝的背景でしか起こらないので、患者を層別化することも可能になる。このような状況は、通常の環境下でALS患者が異質であることが知られていることを考えると、驚くべきことではない。
ALSの病因は依然として謎に包まれており、有効な治療法がない現状では、この複雑な疾患を研究するための学際的なアプローチが不可欠である。最近、アメリカ大陸でジカウイルスやウエストナイルウイルスなどの病原体が出現したことで、神経-ウイルス-免疫学的な軸の理解に大きな進展が見られた。しかし、ALSにおけるウイルスの役割を裏付けるデータがない現状では、このような複雑な関連性を調査するための新たな枠組みを構築する必要がある。感染症の専門家と神経変性の研究者がより緊密に協力することで、将来的に治療につながる大きな生物学的洞察が得られるかもしれない。