
Mechanisms Behind Pyrroloquinoline Quinone Supplementation on Skeletal Muscle Mitochondrial Biogenesis: Possible Synergistic Effects with Exercise
https://pubmed.ncbi.nlm.nih.gov/29714638/
オンライン公開:2018年05月01日
Paul Hwang(MS)Darryn S. Willoughby(PhD)FACN
米国テキサス州ウェーコ、ベイラー大学健康・人間パフォーマンス・レクリエーション学部、運動・生化学栄養研究所
概要
持久力運動トレーニングは、筋肉内の適応を引き起こし、ミトコンドリア生合成、酸化能力、ミトコンドリア密度、ミトコンドリア機能の向上につながることが明らかになっている。ミトコンドリアの生合成は、転写因子であるペルオキシソーム増殖因子活性化受容体γ(ガンマ)共役因子1αの活性化によって制御されている。このミトコンドリア生合成のマスターレギュレーターは、核呼吸因子(NRF-1,NRF-2)やミトコンドリア転写因子Aを活性化し、ミトコンドリアサイズの拡大やミトコンドリアDNAの転写を可能にする。ピロロキノリンキノン(PQQ)は、酸化還元調節、細胞エネルギー代謝、ミトコンドリア生合成など様々な生理過程に関与し、強力な抗酸化作用を持つ新規サプリメントとして同定されている。運動とPQQの補給はともにミトコンドリアの生合成に関連するメカニズムを持っているため、PQQによる差動的なエルゴジェニック・ベネフィットが得られると考えられている。しかし、運動とPQQの併用についての研究はほとんどない。そこで、今回の文献レビューでは、PQQとミトコンドリアの生合成のメカニズムを包括的に紹介することを目的としている。運動はミトコンドリアの生成を示す分子反応を誘発するので、PQQと運動が相乗的な反応を引き起こす可能性は十分にある。
重要なポイント
- 持久的な運動を行うことで、骨格筋の適応が可能となり、ミトコンドリアの生合成が促進され、酸化能力、ミトコンドリア密度、ミトコンドリア機能が向上する。
- ピロロキノリンキノン(PQQ)は、レドックス調節、細胞エネルギー代謝、ミトコンドリア生合成、抗酸化力などの生理的プロセスに関与する新規サプリメントとして同定されている。
- PQQを摂取することで、骨格筋内のミトコンドリアの生合成を示す分子シグナリング反応をアップレギュレートできることを裏付ける証拠が出ていた。
- 持久的な運動とPQQの補給の両方がミトコンドリアの生合成を示す分子反応の増加を誘発するならば、PQQと運動の両方が相乗的なエルゴジェニックな反応を引き起こす可能性がある。
- しかし、PQQの摂取と運動の併用がどのような役割を果たすかについての研究はほとんどない。今後、PQQと運動の併用によるエルゴジェニック効果を検証する必要がある。
キーワード
PGC-1a; ミトコンドリア; NRF-1; mtDNA; CREB; TFAM; 骨格筋; PQQ; 運動
はじめに
骨格筋は,全身の約40%を占める非常に動的で代謝の活発な組織である[1]。ミトコンドリアは,細胞が必要とするエネルギーを供給するために,栄養基質からアデノシン三リン酸(ATP)を効率的に生成する能力を持っている。具体的には、ミトコンドリアの役割は、トリカルボン酸(TCA)サイクルに入る代謝物(糖質や遊離脂肪酸)を酸化して、酸化的リン酸化によってATPを合成することである。さらに,ミトコンドリアは,プログラムされた細胞死(アポトーシス),細胞周期の制御,酸化還元の恒常性,カルシウム関連のシグナル伝達にも関与していることが知られている[2]。
ミトコンドリアの生合成は,ミトコンドリアの数と大きさを維持する複雑な生物学的プロセスであり,核ゲノムとミトコンドリアゲノムが関与している[3,4]。核ゲノムは、ミトコンドリア内のほとんどの遺伝子をコードしているが、ミトコンドリアは37個の遺伝子からなる独立したゲノムを持っていることが知られている[1]。これらの遺伝子のうち13個は、電子輸送系の必須タンパク質をコードしていることが知られている[1,5]。そのため,ミトコンドリアの形成には,核とミトコンドリアの両方のゲノムが協調的に発現し,特定の補因子が作用して別々のミトコンドリアタンパク質がコードされる必要がある[5]。この点において,ペルオキシソーム増殖因子活性化受容体ガンマ共役因子1α(PGC-1a)は,様々な転写因子を共役化して発現を増強し,ミトコンドリアの生合成に関わる遺伝子の発現をアップレギュレートする共役転写調節因子として同定されている[6,7]。これらの重要な転写因子には、核内呼吸因子(NRF-1,NRF-2)ミトコンドリア転写因子A(TFAM)およびエストロゲン関連受容体(ERR)が含まれる[3,4,6]。NRF-1/2とTFAMは、ミトコンドリアの主要な酵素の転写を制御すると同時に、ミトコンドリアDNA(mtDNA)の合成を仲介する役割も果たしているようである[8]。したがって、このミトコンドリア生合成のプロセスは、電子輸送系の重要な酸化的リン酸化コンポーネントや、ミトコンドリアマトリックス内のTCAサイクルに関連する酵素を発現するタンパク質として転写・翻訳されるDNAを追加生成するために極めて重要である [3,4,9]。
急激な運動により,骨格筋におけるPGC-1aのmRNAの発現量が増加することが研究者らによって示されている[9]。さらに、PGC-1aを過剰に発現させると、筋細胞内のミトコンドリア含有量が増加し、酸化能力が向上するとともに、より酸化的な表現型への繊維タイプのシフトが誘導されることがわかった[10,11]。PGC-1aは、グルコネオジェネシス、熱産生、ミトコンドリアバイオジェネシス、抗酸化酵素の制御など、さまざまな代謝プロセスに関与しているほか、骨格筋の運動誘発性適応にも関与していると考えられている[3]。
一貫した持久運動を取り入れることは、運動能力を高めるだけでなく、トレーニング刺激の特異性に応じた適応を可能にすることが確立されている[5,12]。持久運動トレーニングは、筋ミトコンドリアの増加を誘発する骨格筋の適応を可能にする[5]。さらに、1回の運動でミトコンドリアの生合成が促進され、その結果、ミトコンドリアのタンパク質の発現が増加する[13]。予想通り、トレーニング研究では、一定期間の持久力トレーニングによって、ミトコンドリアのコンテントの上昇、脂肪酸化の促進、耐疲労性の向上、持久力の向上など、適応がさらに強化されることが示されている[14]。そのため、PGC-1aの発現量やタンパク質量の増加が、このような適応を媒介することは明らかであると考えられる。したがって、ミトコンドリア・バイオジェネシスのアップレギュレーションは、ミトコンドリアのエネルギー代謝作用を最大化するのに有効であると考えられる。ミトコンドリア・バイオジェネシスは、基質の酸化とそれに伴うATPの生成を通じて、筋肉のエネルギー需要を最適化することができる。
研究者たちは、ピロロキノリンキノン(PQQ)のメカニズムを探り、強力な抗酸化物質として高く評価されていること、ミトコンドリア生合成の活性化を引き出すこと、成長因子として関与していること、心臓保護作用や抗糖尿病作用を示すことを発見した[15-18]。メカニズム的には、PQQがcAMP-response element binding protein (CREB)をリン酸化し[19]、PGC-1a mRNAの発現をさらに増加させることで、ミトコンドリアの生合成との関連性を明らかにした研究がある[20,21]。この点から、もしPQQがPGC-1aを活性化し、CREBの活性化に関与し、電子輸送鎖(ETC)複合体のミトコンドリア活性を高めることができるのであれば、PQQが核内呼吸因子(NRF-1,NRF-2)やTFAMのレベル上昇に関与している可能性がある[3]。これらのコアクチベーターは、エネルギー代謝やミトコンドリアの生合成、酸化能力の向上などに関与していると考えられるので、PQQも間接的に関与している可能性がある[19,20]。しかし、このようなPQQのレドックスバランス、ミトコンドリアバイオジェネシス、ミトコンドリアの機能障害・機能、さらにはヒトの運動パフォーマンスへの影響を調べた研究は少ないようである。さらに、げっ歯類におけるPQQ補給によるミトコンドリアのバイオジェネシスの作用機序が、ヒトにおいても観察できるかどうかを確認するためには、さらなる研究が必要である。
今回の動物モデルや培養細胞を用いた研究では、PQQを摂取することでミトコンドリアバイオジェネシスのマーカー(PGC-1a、CREB、NRF-1/2)が上昇し、筋肉組織がより代謝の高い酸化的な繊維タイプにリモデリングされることが示唆されているため、PQQを摂取して持久力トレーニングを行うことで、ヒトにおいてもエルゴジェニックな結果が得られる可能性があると考えられる。PQQは体内で合成することはできないが、植物性の食品から幅広く摂取することができる[15,22,23]。さらに、持久運動がPGC-1aの活性化を介して酸化能力の向上やミトコンドリアの生合成を促進するのであれば、PQQを補給することで、運動をしている人間に付加的な効果をもたらす可能性がある。しかし、現在のところ、研究が不足しているため、このような人間のエルゴジェニックな可能性の融合の可能性が損なわれている。
そこで、この文献では、PQQとその作用機序、ミトコンドリアの生体形成、およびミトコンドリアの機能・生体形成における運動の役割について、より包括的なイメージを得るために科学的な文献を調査することを目的としている。
ミトコンドリアと生体形成の分子基盤
ミトコンドリアは真核生物の必須小器官であり,生体組織の動力源として,解糖系と脂肪分解系の産物を処理してATPを生成する機能を持っている[2,6]。さらに,ミトコンドリア生合成は,新しいミトコンドリアを作るプロセスである。この非常に複雑なプロセスには,ミトコンドリア内の必須遺伝子だけでなく,多数の核ミトコンドリア遺伝子が協調して転写される必要がある[3]。ミトコンドリア遺伝子の転写は,主に転写補因子PGC-1aを介して行われる[1-3,24]。この補助転写活性化因子は,エネルギー代謝に関連する遺伝子を制御し,ミトコンドリアの生合成をコントロールする[9]。
PGC-1aの発現は,核内呼吸因子(NRF-1/NRF-2)との相互作用や共活性化を制御しており,NRF-1/NRF-2は,シトクロムc(COX)やCOXオキシダーゼサブユニットIVのコード化を介したETCを介した酸化的リン酸化,(TFAMの活性化を介した)mtDNAの転写・複製,タンパク質輸入装置やタンパク質のアセンブリに関与する遺伝子の発現を制御する機能を有している[3,9,14]。したがって,PGC-1aは,細胞内のシグナルに反応して,これらのNRF-1/2の転写因子と結合することで,標的転写産物の産生をさらに促進する。
PGC-1aは,N-terminal領域,Middle領域,C-terminal領域からなる構造をしている[9]。N末端領域はトランザクティベーションドメインに相当し,ステロイド受容体 coactivator-1とCREBを含む2つの補助的な活性化因子が組み合わされている[25]。
単回の運動でPGC-1aのmRNAおよびタンパク質レベルの発現が増加することはよく知られている[3]。さらに、運動を繰り返すと、NRF-1やTFAMと並んで、PGC-1aのタンパク質発現が増加する。PGC-1aは、転写因子である筋細胞エンハンサー因子2(MEF2)およびCREB、ならびに活性化転写因子2(ATF-2)のPGC-1aプロモーター結合活性によって、転写レベルで制御されている[26]。MEF2は、PGC-1aの転写を促進する機能を持つが、PGC-1aの標的でもあるため、PGC-1aが自身の発現を制御する自己調節的なフィードバックループを形成している[27,28]。PGC-1aによって活性化されたNRF-1/2は、転写因子MEF2の発現を誘導し、COX遺伝子やグルコーストランスポータータイプ4 (GLUT4)を活性化するとともに、PGC-1aの発現を増強することで、間接的にミトコンドリア遺伝子や代謝遺伝子の発現に影響を与える [3,6]。また,NRF-1/2は,ミトコンドリア呼吸複合体のサブユニット,ヘム生合成酵素,ミトコンドリア輸入装置に関連するタンパク質,ミトコンドリアリボソームタンパク質,tRNA合成酵素などをコードする多くの核内遺伝子の発現を制御する機能も有している[29,30]。運動は、核内でのPGC-1aやNRF-1との相互作用を強め、ETCタンパク質の核内ゲノムにコードされた遺伝子の転写を増加させることができる。
PGC1aとNRF-1は、ミトコンドリアの重要な転写因子であるTFAMを活性化する。TFAMが活性化されると、ミトコンドリアの生合成を促進する役割を担うmtDNAの転写や複製の制御を助ける働きをする。また、TFAMは、DNAを曲げたり、巻いたり、ほどいたりする能力があることも特徴である[3]。Dloopと呼ばれる非コード領域は、mtDNAの転写を制御している。さらに、DNAのこの領域には、ETCに対応するタンパク質をコードする13個の転写を開始するのに必要なプロモーターが含まれている[6]。ミトコンドリアのRNAポリメラーゼ、TFAM、ミトコンドリア転写因子B2がmtDNAの転写を制御する役割を果たしていることが研究で明らかになっている[31]。興味深いことに、運動に反応してPGC-1aがミトコンドリアに移動し、mtDNAのDループ上でTFAMと複合体を形成して、ミトコンドリアゲノムにコードされた遺伝子の転写とミトコンドリアの生合成のためのmtDNA複製を促進することを示唆するデータがある[32-34]。他にも、PGC-1aと共働して様々なミトコンドリア遺伝子を活性化し、遺伝子発現を高める役割を担うERR(ERR-a, ERR-b, ERR-g)などの転写因子が発見されている[29]。これらの受容体は核ホルモン受容体ファミリーの一員であり、様々なホルモンシグナルに応答してミトコンドリアの生合成を促進する機能を持つ。特に,ERR-aは,酸化的リン酸化,脂肪酸酸化,TCAサイクル,ミトコンドリアの融合・分裂に関与するようなミトコンドリア関連因子をコードする核内遺伝子の転写を制御することが確認されている[35]。運動に応答して、PGC-1aはロイシンリッチ核受容体相互作用モチーフ(LLKYL)を介してこれらのERRと協調的に働き、多くのミトコンドリア機能に関与する核ゲノムにコードされたミトコンドリア遺伝子の転写を誘導することが知られている[36]。興味深いことに、C2C12筋管内でのPGC-1aとERRによる転写活性は、PGC-1 and ERR-induced regulator in muscle (PERM)-1と呼ばれるタンパク質に依存している可能性を示唆するデータがある[24]。運動時には、PGC-1a mRNAの転写とPGC-1aタンパク質の細胞内移行の両方を制御する様々なシグナル伝達経路が存在する[1]。
運動に伴い、PGC-1aの発現は、活性酸素種(ROS)の産生の増加、収縮に伴う細胞内カルシウムの増加、ATPの代謝を介した細胞のエネルギー状態(アデノシン一リン酸(AMP)の上昇、AMP活性化プロテインキナーゼ(AMPK)の活性化)などによって制御される[3]。[3]. 骨格筋の代謝マスタースイッチであるAMPKの活性化は,アセチル-CoAカルボキシラーゼ(脂肪酸合成に関与)やAS160(GLUT4輸送に関与)などの様々な下流のエフェクターターゲットをリン酸化・不活性化するとともに,COX,クエン酸シンターゼ,コハク酸デヒドロゲナーゼなどのミトコンドリア含有量に関連する様々な遺伝子の発現を増加させる[37-39]。
耐久運動トレーニングは,最終的なエネルギー枯渇と慢性的な代謝ストレスに関連しており,そのためAMPK活性がアップレギュレートされる。AMPKは,PGC-1aを直接リン酸化することで,ミトコンドリアの生合成を促進する[40]。注目すべきは、運動時以外にも、断食時にもAMPKの活性化を介してPGC-1aの発現を高め、酸化能力を上昇させることが示されていることである。長時間の運動や断食・エネルギー制限を行うと、遊離脂肪酸の濃度が上昇し、筋肉内の転写因子PPARδ(デルタ)の発現が高まり、その結果、ミトコンドリアの数が増加する[4]。さらに、AMPKは、細胞内のニコチンアミドアデニンジヌクレオチド(NADC)レベルを上昇させることで、SIRT1(silent mating type information regulation 2 homolog 1)の活性化を促する。これを受けて,SIRT1はPGC-1aを脱アセチル化し,酸化的な代謝とミトコンドリアの生合成のプロセスがさらに促進される[41].また、AMPKがプロテインキナーゼAアンカープロテイン1と結合できるという証拠もあり、AMPKとミトコンドリア呼吸の制御が相互に関連していることが示唆されている[42]。重要なことは、筋収縮活動の期間中、PGC-1aの転写活性およびPGC-1aプロモーター活性を高めるためには、カルシウムの存在とAMPKの活性化が不可欠であるということである。細胞質のカルシウムレベルが上昇すると、カルシウム/カルモジュリン依存性プロテインキナーゼおよびカルシニューリンという酵素が活性化され、CREBのリン酸化を介してPGC-1aの発現を増加させるように機能する[4]。細胞質のカルシウムが上昇すると、p38MAPKが活性化され、PGC-1aの発現がさらに増加する可能性がある[4]。p38MAPKはPGC-1aを直接リン酸化し,この共転写因子を転写的に制御することが知られているため,もう一つの上流のシグナル伝達経路としてMAPK経路が挙げられる[37]。さらに,p38MAPKは,運動によって生じる活性酸素の上昇に対しても高い反応性を示すことが知られている[43]。さらに,p38MAPKは,MEF-2やATF-2などの転写因子をリン酸化することで,PGC-1aを転写的に制御することもできる[44]。
ミトコンドリア内でのミトコンドリアバイオジェネシスとマイトファジーを制御するために存在するシグナル伝達経路を調査する研究はまだ増え続けている。PGC-1aの発現増加とミトコンドリア生合成の誘導に対応する分子経路を複雑にする可能性のある要因は数多く重なっている。この点からも、ミトコンドリアの生合成に関わる分子経路の制御に関する研究を進める必要がある。さらには、ミトコンドリアのターンオーバーを決定する要因の解明にも役立つと考えられる。
PQQの歴史と化学的性質
PQQ (4, 5-dihydro-4, 5-dioxo-1 H-pyrrolo [2, 3-f] quinolone-2, 7, 9-tricarboxylic acid) は,アルコールや糖のデヒドロゲナーゼのような原核生物の様々なデヒドロゲナーゼのレドックス補酵素として機能する芳香族三環式O-キノンであることが確認されている[15,18,23]。この化合物のレドックス関連の挙動は,o-キノンのグループ化によるものと思われ,キノン型(PQQ)に酸化されるか,ラジカルまたはセミキノン(PQQH)に半還元されるか,あるいはキノール型(PQQH2)に完全に還元される可能性がある[45]。さらに,PQQは,酸化還元バランスを整えるために1つまたは2つの電子を受容する機能を持つ補欠基であることが指摘されている[23]。繰り返しになるが,PQQはセミキノン中間体との反応により,ピロロキノリンキノール(4, 5-dihydroxy-1 H-pyrrolo [2, 3-f] quinolone-2, 7, 9-tricarboxylic acid, PQQH2)に可逆的に還元される[15]。PQQが様々な有機基質から酸素分子(O2)への効率的な電子伝達触媒として作用することを示す研究があり,キノプロテインモデルに基づく反応を提示している[15]。さらに,PQQは,アスコルビン酸,NAD(P)H,グルタチオンの存在下で,2電子還元反応を起こしてPQQH2を生成することができる[46,47]。その後、生成されたPQQH2は、2還元当量の分子状酸素(O2)をスーパーオキシドアニオン(O2¡)に還元することで、元のキノンに酸化される。さらに、スーパーオキサイドアニオンは、スーパーオキサイドディスムターゼによって過酸化水素(H2O2)に分解される。したがって,PQQは連続的な酸化還元サイクル反応を触媒する能力を持っていると考えられる[15]。
ここで重要なのは,PQQが継続的な酸化還元サイクルを行う能力を持つことで,活性酸素であるH2O2とO2¡の生成を通じてプロオキシダント作用を引き起こすことである[48]。さらに,PQQは,システイニルチオールの酸化を介して,タンパク質の酸化的修飾を誘発することもある[48]。PQQは水溶性で熱にも強く,求核剤(特にアミノ酸)と反応してイミダゾピロロキノリン(IPQ)化合物を形成することが確認されている[15,23]。
歴史的には,1960年代に細菌のメタノール脱水素酵素とグルコース脱水素酵素の補酵素を別々の研究室で研究した結果,細菌内に同じ新規補酵素PQQが存在することが明らかになった[45]。さらに、研究者たちは、メチル化細菌のキノタンパク質であるメタノール脱水素酵素からこのプロテスタント基を抽出し、X線結晶構造解析によってその分子構造を明らかにすることができた[15]。その結果、PQQの構造を特定することができた。繰り返しになるが、PQQはメタノールデヒドロゲナーゼの補酵素として、連鎖栄養細菌で初めて同定された[49,50]。PQQは、補酵素としての役割だけでなく、細菌の成長や代謝に関わる栄養因子としての役割も示唆されている[18]。PQQと相互作用するタンパク質をキノプロテインと呼んでいた[23]。また、トパキノンやトリプトファン・トリプトフェニルキノンなどの補酵素も、PQQと同様にキノプロテインとして同定された[23]。また、真核生物で初めてPQQ依存性の糖酸化還元酵素をキノコ(担子菌Coprinopsis cinerea)の中から発見した[51]。また、PQQは細菌の成長を促進することも確認されている[52]。KillgoreらはPQQの哺乳類への影響を観察しているが[53]、哺乳類内でPQQを補酵素として持つ酵素はこれまでに確認されていない。
さらに、Killgoreら[53]は当初、げっ歯類の明確な食餌からPQQを除去すると、成長障害、免疫不全、生殖能力の低下、呼吸量の減少などの様々な反応が起こることを確認した。この重要な発見に基づいて、研究者たちは、動物の体内でPQQを補綴基として利用できる酵素を特定しようとした[54]。この点について、日本の研究者たちは、哺乳類の遺伝子の中で、PQQ依存性キノプロテインと一部相同性を示す1つの遺伝子を同定し、報告した[55]。さらに、その遺伝子産物がマウスのリジン代謝において2-アミノアジピン酸セミアルデヒド(AAS)を2-アミノアジピン酸(AAA)に酸化するデヒドロゲナーゼであることから、PQQは新たなレドックス・コファクター・ビタミンとして作用する可能性があると報告している[55,56]。さらに、この研究者たちは、ネズミの肝臓と血漿中のAASとAAAの活性が、哺乳類におけるリジン分解の指標になると結論づけた。ネズミにPQQを摂取させると、AAAの濃度が低下し、リジン代謝の変化を示すことがわかった[55]。このことから、PQQ依存性の脱水素酵素が同定され、リジンというアミノ酸の分解に重要な役割を果たしていると考えられ、ビタミンB群の一種であることが示唆された。さらに、PQQを欠いたマウスでAAAが減少したことから、PQQを欠いたマウスでは、マウス内のAAS-デヒドロゲナーゼ活性が障害されていることが示唆された。このことから、動物のリジン代謝に必要なAAS-dehy-drogenaseが全体的に正しく機能するためには、PQQが酸化還元の補酵素として必要であることが示唆された[55]。
この発見は、PQQが新規のビタミンB型であるという主張を確認するために、研究者たちがこの発見を再現しようとする大きな議論を引き起こした[56,57]。さらに、PQQのB型ビタミンとしての主張は、構造的なβプロペラ配列をPQQ結合モチーフとして不適切にラベル付けしたデータベースの配列解析に依存していたため、大きな批判を受けた[56]。簡単に言えば、リジン代謝のAAS-デヒドロゲナーゼ酵素の同定は、7枚羽根のβ-プロペラ構造を持っているようだが、PQQ依存性のデヒドロゲナーゼであることを示す証拠はない[56]。さらに、PQQはバクテリアの多数のデヒドロゲナーゼ酵素(qui-noprotein)の必須の補因子として同定されているにもかかわらず、哺乳類との類似性を示す直接的な証拠はない[56]。したがって、笠原と加藤[55]の主張とは対照的に、これらの研究者は、必須の補酵素として酵素に必要とされる必要性が証明されるまで、PQQはビタミンとして同定されないと述べている[56]。Ruckerら[57]はKasaharaとKato[55]の所見の再現を試み、ラットの肝臓抽出物からのAAS酸化速度がPQQ状態の変化によって有意に変化しないことを観察した。さらに、PQQの欠乏はAAA/リジン比に影響を与えないようで、PQQを動物のレドックス補酵素として同定するという主張に疑問を投げかけた。興味深いことに、研究者のKasaharaとKato[55]は、リジン代謝におけるPQQの役割についての発見をさらに立証する回答を発表した[57]。リシルオキシダーゼ、ドーパ脱炭酸酵素、ドーパミンβ-水酸化酵素、プラズマアミンオキシダーゼ、ジアミンオキシダーゼなど、他の哺乳類の酵素にもPQQが補酵素として含まれていることが示唆された[22]。しかし、これらの示唆された知見に異議を唱える対照的な証拠がある[22]。現時点では、動物においてPQQを補酵素として含む正確な酵素を特定するための真のコンセンサスは得られていないが、PQQが成長、発達、および健康上の好ましい結果において生理的に重要であることを強調するデータは数多く存在する[15,18,23]。PQQの補給が動物や人間にとって生理的に有益であることを示す研究については、で紹介する。
PQQの供給源
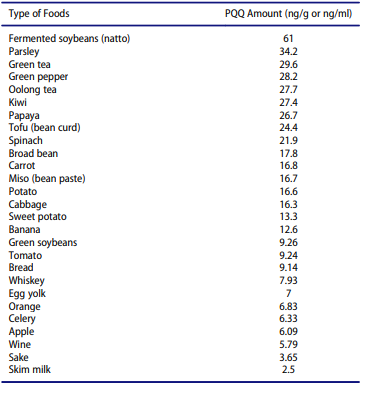
PQQは自然界に遍在していることが知られており、納豆、お茶、ピーマン、パセリ、キウイフルーツ、ヒトの牛乳など様々な食物に含まれている[22]。熊澤ら[58]は,水酸化フェニルトリメチルアンモニウムで誘導体化した遊離PQQを同位体希釈法を用いたガスクロマトグラフィー/質量分析法で測定するという画期的な方法を開発した。この方法により、様々な食品(野菜、果物、お茶)中の遊離PQQを定量することができ、生重量で3.7~61ng/g、液体食品ではng/mlの範囲であることが報告された(Table 1)[22]。また、液体食品中の遊離PQQ量を0.19〜7.02ng/g(ng/ml)の範囲で特定するために、信頼性の高い液体クロマトグラフィー/エレクトロスプレーイオン化タンデム質量分析法によるPQQの分析方法が開発された[59]。具体的には、レドックスサイクリング法を用いて、卵やスキムミルクに高濃度の遊離PQQが含まれていることを報告している[22]。興味深いことに,これらの知見に基づいて,人間は1日平均0.1〜1.0 mgのPQQおよびその各構造誘導体を摂取することができると示唆されている[60]。繰り返しになるが、高等生物におけるPQQの生合成は示されていない。したがって、PQQの生産は、食事由来または腸内細菌によってのみ得られる[15,22]。微生物におけるPQQの生合成は、様々な細菌の保存された遺伝子の機能を前提とした推定経路を介して行われているというデータがある[61]。微生物におけるPQQの生合成については、本レビューの範囲を超えるが、以下の論文[15,45]でさらに詳しく説明されている。以上のことから、ヒトの腸内細菌は最小限のPQQを合成することができるが[62,63]、人体のPQQの主な供給源は食事であると考えられている。
PQQ補給の生理的役割
人でも動物でも、細菌の常在菌株や植物にPQQが遍在していることから、常にPQQにさらされていることが示唆された[18]。この点に関して、研究分野ではいくつかの生理学的特性がPQQの補給と関連している[15,18,23]。ビタミンとしてのPQQの決定的な役割はヒトでは立証されていないが、PQQが細胞シグナル伝達経路に関連していること[19,64]、強力な抗酸化剤としての役割[20,65]、成長促進活性[15]や抗糖尿病効果[17]を示すこと、心臓や神経の保護剤として機能すること[17,66,67]、ミトコンドリア含有量の調節に役割を果たすこと[16,19,20]などの強い証拠が残っている。以下では、主にPQQのミトコンドリア含有量の調節効果と抗酸化作用についての先行文献を紹介する。ミトコンドリアは、シグナル伝達を介して代謝経路を制御し、代謝の恒常性や細胞機能全体を調節することが知られている。PQQを摂取することで、ネズミのミトコンドリアの機能と生合成が促進されたというデータがある[15]。PQQを摂取していないマウスの肝臓のミトコンドリア量は、PQQを摂取したマウスに比べて少ないことが、断面積の推定値[16]や、定量的リアルタイムポリメラーゼ連鎖反応によるセリンパルミトイルトランスフェラーゼ活性[20]から明らかになっている。Stitesら[16]は、PQQ欠乏マウスの呼吸商の値が、6時間の間にPQQ補給マウスと比較して低下していることを観察した。ここで重要なのは、6時間の呼吸商(RQ)の許容暴露量を調べるために、マウスを28℃から 16℃の温度変化に導入したことである。しかし,Bauerlyら[20]は,12時間の明暗サイクルの中で最も活動的な時期に,ミトコンドリア量と正の相関があると考えられる実際のエネルギー消費量には有意な差があったものの,治療法によるRQレベルの変化はほとんど見られなかった。このようなRQの異なる結果に代わって、Stitesら[16]は、PQQまたはその関連誘導体(IPQ)が、ミトコンドリアの機能とミトコンドリアの生合成のプロセスに不可欠であると報告した。興味深かったのは、比較的低いPQQ濃度(nmol/g of diet)と低い組織濃度(pmol/L~nmol/L)が、PQQの不足によるミトコンドリア機能の低下を逆転させる可能性があったことである。
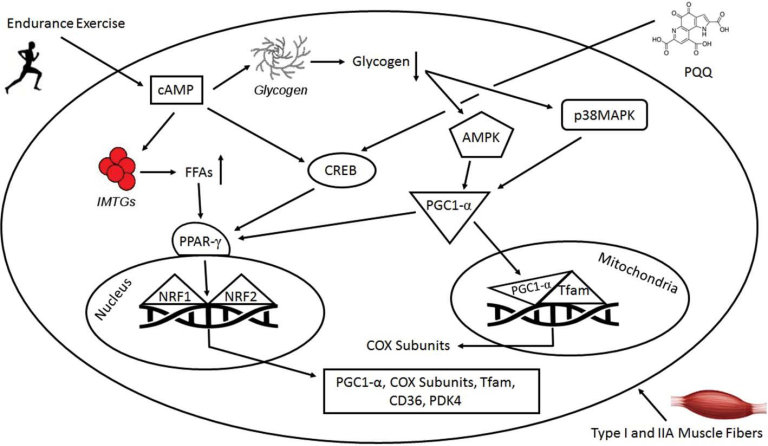
図1 持久運動とピロロキノリンキノン(PQQ)の補給が、ミトコンドリアの生合成に関わる分子機構上のシグナル伝達経路において果たす役割をまとめて表現したもの
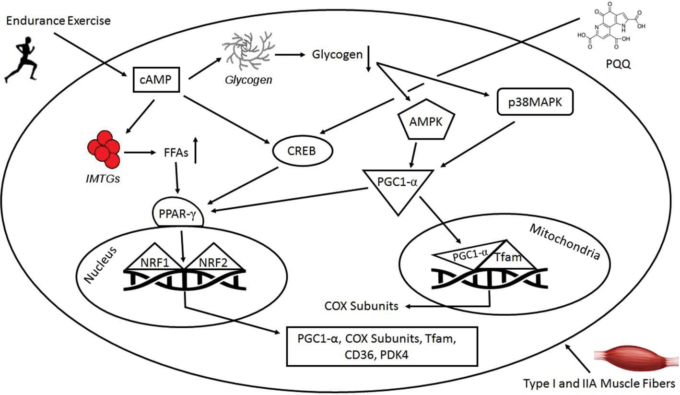
AMPK D AMP-activated protein kinase; cAMP D cyclic AMP; CD36 D cluster of differentiation 36; CREB D cAMP-response element binding protein; COX D cytochrome c; FFAs D free fatty acids; IMTGs D intramuscular triglycerides; PPAR-g(ペルオキシソーム増殖因子活性化受容体γ)PGC-1a(ペルオキシソーム増殖因子活性化受容体γコアクチベーター1α)NFR-1および2(核内呼吸因子)TFAM(ミトコンドリア転写因子A)PDK4(ピルビン酸デヒドロゲナーゼキナーゼ4)など。
PQQの栄養状態を評価するために遺伝子アレイ解析を実施することで、PQQが様々な遺伝子、特にミトコンドリア関連の活動に影響を与えることが示唆されている[68]。PQQの栄養状態の変化は、細胞ストレス、ミトコンドリア生合成、細胞シグナル活性に関連する遺伝子に影響を与えることが示されている[19]。マウスのHepa1-6細胞にPQQを投与すると,ミトコンドリアの生合成に直接関与するCREBとPGC-1aが活性化されることが示されている[19]。PGC-1aは,NRF-1とNRF-2に結合し,それらの活性を同時に高めることでミトコンドリアの生合成を誘導する転写性のコアクチベーターである。マウスのHepa1-6細胞にPQQを投与すると、NRF-1とNRF-2の活性が上昇し、TFAMも上昇するというデータがあり、PQQがミトコンドリアの生合成を促進する分子メカニズムが示されている[19]。さらに,CREBのリン酸化とそれに伴うPGC-1aの発現上昇は,ミトコンドリアの生合成の誘導を修飾する。Chowanadisaiら[19]は、3つの主要な発見を示唆するデータを先駆的に示した。Chowanadisら[19]は、PQQ暴露によるPGC-1aプロモーターの活性化は、cAMP応答要素サイトが保持されている場合にのみ上昇すること、(2)PQQ暴露は、NRF-1とNRF-2の両方の応答要素を含むTFAMと転写因子B1/B2のミトコンドリアmRNAの発現を上昇させること。(3)ミトコンドリア阻害剤を投与する24時間前に、10,20,30マイクロモル単位のPQQを培地に添加した細胞は、PQQを添加しなかった細胞に比べて、細胞生存率が上昇した。これらの結果は、食事で摂取したPQQがミトコンドリアの生合成を促進するという作用機序を確立する上で重要なデータとなる(図1)。しかし、PQQによるCREB-PGC-1aシグナル伝達経路の活性化の分子メカニズムを明らかにした研究はまだ少ない。しかし,今回の結果は,食事によるPQQの欠乏がミトコンドリア数の減少につながるという理解をさらに深めるものである[16]。したがって,PQQが欠乏すると,ミトコンドリアの機能が損なわれ,ミトコンドリア生合成の誘導が低下し,ミトコンドリア数が減少するため,アミノ酸代謝(主にミトコンドリアで代謝されるアミノ酸)に欠陥が生じる可能性があることが示唆される[69]。
Bauerlyら[20]は、PQQを摂取したラットは、PQQを摂取していないラットとは対照的に、二酸化炭素排出量(VCO2)や酸素摂取量(VO2)が低下し、虚血に対する死亡率が30%から40%になったことから、PQQを摂取することでラットの心臓のミトコンドリアが保護されることを確認した。この点から、本研究の虚血・再灌流障害モデルでは、薬理学的用量のPQQが心筋保護作用を示していると考えられた[20]。Taoら[17]は、PQQによる心筋保護効果は、PQQ暴露による抗酸化防御の上昇によるものではないかと提唱している。しかし、この知見を明らかにするためには、さらなる研究が必要である。以上、PQQ暴露とミトコンドリアに関する知見から、PQQはミトコンドリアの生合成や代謝機能、さらには心筋保護にも関与する新規の代謝調節物質であると考えられる。また,PQQは,グルタチオン,ニコチンアミドアデニンジヌクレオチドリン酸(NADPH),システインなどの還元剤と反応して容易にPQQH2に還元されることから,抗酸化作用を示すと広く認識されている[15]。興味深いことに,PQQH2のアロキシルラジカル消去活性は,ビタミンCの約7.4倍であるというデータがある[65]。また、PQQH2の一重項酸素消去活性は、ビタミンCの効果よりも約6.3倍高いことが分かっている[71]。PQQH2は、2分子のα-トコフェロキシルラジカルをα-トコフェロールに速やかに変換する機能を持つことが分かっている[72]。そのため,PQQH2の働きにより,α-トコフェロキシルラジカルのプロオキシダント作用を抑制することができる。さらに、ヒトを対象とした研究では、PQQ Na2(「0.2mg/kg体重」)を単回投与したところ、脂質ヒドロペルオキシドから生成されるマロンジアルデヒドで測定されるチオバルビツール酸反応性生成物の活性が、研究期間中に有意に減衰したという結果が出ている[60]。このように,PQQは継続的に酸化還元サイクルを行うことができるようで,アスコルビン酸の触媒サイクル数が4回と報告されているのとは対照的に,潜在的な触媒サイクル数は2000回近くになると考えられている[18]。ここで重要なのは,PQQが濃度依存的に抗酸化物質として働くことである[73]。Jurkat細胞を用いた実験[74,75]では,10マイクロモル単位まではPQQは主に抗酸化剤として作用しているようだが,50マイクロモル単位を超えるとPQQはプロオキシダントとして作用すると考えられている[73,74]。具体的には,PQQは細胞培養液と相互作用して過酸化水素を生成し,それに伴って細胞死を誘発することで,潜在的なプロオキシダントとして作用する[23,74]。したがって,著者らは,PQQの活性はその環境に依存しており,様々な生物学的システムにおいて抗酸化剤またはプロオキシダントとして作用する可能性があると示唆している[74]。しかし,PQQは,最適な濃度であれば,生体内では細胞を酸化損傷から保護し,生体外では活性酸素の人工的な生成から生体分子を保護する能力があり,非常に強力な抗酸化剤と考えられる[23,76]。
運動中に活性酸素や反応性窒素種(RNOS)が生成されると、骨格筋の損傷や疲労が誘発され、回復に悪影響を及ぼすという説が一般的に信じられてきた[77,78]。このような活性酸素やRNOSによる悪影響を考慮して、運動による活性酸素やRNOSの生成を抑制するために、抗酸化作用のあるサプリメントを摂取することが一般的なアプローチとなっている。しかし、適度なレベルの運動誘発性ROSが、ミトコンドリアの生合成などのプロセスにおいて、細胞のストレスに適応するためのシグナル刺激として重要な役割を果たしていることを論じた文献が増えてきている[77,79,80]。さらに、ミトコンドリア・バイオジェネシスの増加は、一貫した持久力運動トレーニング後の骨格筋における適応である。運動によって活性酸素が生成されるメカニズムや、酸化還元に敏感なシグナル伝達経路については、この総説の範囲を超えているが、この話題をさらに詳しく論じた論文がいくつか引用されている[77-79]。持久力関連の活動におけるエルゴジェニック補助のために抗酸化サプリメントを取り入れることを検討している文献の中で、真のコンセンサスは得られていない[79]。抗酸化サプリメントのパフォーマンスへの効果の不一致は、トレーニングプログラム、対象となる生物種、抗酸化サプリメントの種類と投与量、およびそのような実験プロトコルの期間が異なることが原因であると考えられる。とはいえ、持久運動トレーニングがミトコンドリア生合成のマーカーの上昇を促し、その結果、骨格筋のミトコンドリア含有量と酸化能力が上昇することは広く理解されている[77]。しかし、抗酸化物質の補給が、急性の運動によるミトコンドリア生合成のシグナルマーカーの上昇に悪影響を及ぼす可能性を示唆する強力な証拠がある[80-83]。このように、食事による抗酸化物質の補給が、運動に伴う有益な酸化還元シグナル効果を軽減するかどうかについては、まだ明確なコンセンサスが得られていないようである[79]。抗酸化物質の補給によってミトコンドリアの生合成にどの程度の影響があるかは、補給の期間、タイミング、トレーニングの状態、運動プログラム、利用した抗酸化物質の種類など、様々な要因によって複雑になる。PQQは抗酸化作用を示すことが知られているので、運動中に補給するとミトコンドリアの生合成のためのシグナル伝達が阻害され、運動適応に悪影響を及ぼす可能性があると考えられる。しかし、PQQの補給がミトコンドリアの生合成を促進するメカニズムを明らかにした研究がいくつか引用されている[16,19,84]。さらに、ヒトにおけるPQQ補給のエルゴジェニック効果を調査した研究は少ない。このため、PQQの摂取と運動との組み合わせによる効果を理解するためには、さらなる研究が必要である。また、今後、PQQの補給と運動トレーニングの役割を検討することで、ミトコンドリアの生合成の分子メカニズムをさらに解明することができる。PQQ補給によるその他の生理学的効果については本レビューの範囲を超えるが、その他の健康上の利点について簡単に紹介する。PQQは神経成長因子(NGF)の産生を促進することで神経保護効果をもたらすことが示唆されている[85]。さらに、PQQの補給は、げっ歯類の酸化ストレスによる神経衰弱の予防に有効であると報告した研究がある[86]。Kuoら[21]の興味深い研究によると、PQQの投与は、除神経により誘発される(C57BL6/Jマウスに後肢坐骨軸索切除術を施した)筋萎縮とそれに伴うミトコンドリア活性の低下を弱めることが示されている。右後肢の坐骨神経を切断して片側の除神経を行うと、著しい筋萎縮(術後21日目)と酸化繊維表現型への特徴的な変化が見られた。酸化的繊維表現型は、ヘマトキシリン・エオジン染色による繊維断面積の減少と、ニコチンアミド・アデニン・ジヌクレオチド(NADH)NADH-テトラゾリウム還元酵素、COXオキシダーゼ染色の強度が高いことが特徴であった。左肢は内部対照とした。PQQを継続的に補給することで、除細動後の筋萎縮を遅らせることができた。さらに、PQQの投与により、PGC-1aの発現量やタンパク質量、ミトコンドリアの電子輸送鎖複合体活性が改善されたことも報告された。さらに、このことは、ミトコンドリアの酸化的リン酸化のバイオエナジェティックな変化を伴うI型およびII型の両方の筋線維の保存につながった[21]。この研究結果は、解糖系から酸化系への筋線維の切り替えを制御するPGC-1aの役割と、骨格筋の除神経後に酸化的リン酸化複合体の機能を回復させるためのPQQの補給が有益であることを示している。
また,認知機能に関して,ヒトにおけるPQQ補給の効果を調べた研究がある[87]。PQQを補給することで、ストレス、疲労度、睡眠の質が良好に改善され、様々なアンケートを実施して測定した[87]。また、PQQ-Na2を摂取することで、肌の状態や脂質代謝が改善されることが確認されている。
PQQの安全性と投与量に関する考察
PQQ-Na2を含む栄養補助食品は 2009年に米国食品医薬品局(Food and Drug Adminis-tration)が正式に通知を受理した後、米国内で商品化された。また、副作用の報告はない。ヒトでの利用特許を作成するために、ヒトでの安全性試験がいくつか実施されている[18]。1つの試験では、PQQを20または60mg/日で4週間、成人10人ずつの2群に二重盲検法で投与した(プラセボまたはPQQ)。これらの試験は市販の医薬品試験施設で実施され、両レベルの用量で標準的な臨床試験において副作用は報告されなかった[90]。肝毒性に関する機能検査の報告結果も正常であった。なお、PQQを1日60mgの高用量で投与しても、尿中のN-アセチル-β-(D)-グルコサミニダーゼ活性は上昇しなかったという。この化合物は、腎尿細管細胞のリソソーム画分内に存在する腎加水分解酵素として同定されている[90]。腎尿細管機能の異常は,尿中の本化合物の上昇を伴うと考えられる。ラットを用いた経口毒性試験が実施され,14日間の予備試験に続いて28日間の反復投与試験が行われた [91]。この試験の最終段階では,13週間の亜慢性試験が実施された。致死量の中央値は,雄で1,000~2,000 mg PQQ-Na2/kg体重,雌で500~1,000 mg PQQ-Na2/kg体重であったと報告されている[91]。13週間の亜慢性試験では、毒性学的に有意な所見は観察されず、臨床化学的および病理組織学的変化はいずれも対照範囲内であり、用量依存性はなく、対照群と同様に発生した[91]。このように,ラットにおける「観察不能な副作用レベル」は約100 mg PQQ-Na2/kg体重と報告されており,これは13週間の亜慢性試験で試験された最高用量と相関していた。興味深いことに、これらのラットの死後の病理検査では、腎尿細管の損傷や炎症の可能性があるため、PQQ投与による急性の負の副作用が腎臓で観察されることを示唆する証拠が示された[92]。しかし、低用量であれば、病理学的な観察結果が出ないことを言及することは重要である。以上、PQQ補給の安全性と経口毒性を強調した研究結果から、PQQを1日60mgまで摂取しても、急性の副作用や毒性の証拠はないと考えられる。
興味深いのは、PQQは他の生物学的因子(レスベラトロールなど)と比較してユニークな点であり、ネズミの体内でPQQを比較的低い薬理学的用量または健康補助食品で、ミトコンドリア関連の障害に効果があることである[20]。レスベラトロールはミトコンドリアのバイオジーンを誘発することができるが、動物モデルを用いた研究では、反応を誘発するための用量は200から500mg/kg食であった[93]。実際には、これは通常の赤ワイン1本に含まれる量の約100倍に相当する[94]。この点、マウスの成長・発達を促すには、PQQを0.2〜0.3mg/kg食とするのが妥当であると考えられる[95]。吸収の推定については、[14C]PQQをマウスに経口投与すると、PQQは下腸で容易に吸収され(”62%”)最終的には24時間以内に腎臓から排泄された(”81%”)と考えられる[15]。さらに、PQQを単回投与(0.2 mg PQQ-Na2/kg体重)すると、投与3時間後には血清中の濃度が10 nM程度でピークに達すると考えられている[60]。これらの研究から、PQQ-Na2の薬物動態は、他の水溶性ビタミンB群化合物と類似しているのではないかと考えるのが妥当である[15]。PQQ-Na2の血清中での上昇とそれに伴うクリアランスは、尿中で観察された変化に追随している可能性がある[60]。
結論と今後の展望
持久的な運動を行うことで、ミトコンドリアの生成が促進されることが明らかになった。これらの顕著な適応は、PGC-1aの高い発現と連続したタンパク質の生成によって明らかになった。持久力トレーニングによる筋肉内の適応には、酸素輸送と利用の増加、ミトコンドリアと毛細血管の密度の上昇、酸化的リン酸化酵素の濃度の上昇、疲労への耐性の向上、ミトコンドリアバイオジェネシスのアップレギュレーションなどがある[14]。このように、持久運動トレーニングを繰り返すことで、筋肉の酸化能力が最適化され、ATP生成のための基質(糖質と脂肪酸)の酸化が効果的に行われ、さらに重要なことには、運動によって筋肉内に発生した活性酸素に対する抗酸化防御のための内因性酵素の発現が上昇すると考えられる。PGC-1aは、AMPKやp38MAPKの活性化、運動による活性酸素の発生、細胞質のカルシウム上昇などを介して、有酸素運動に反応して活性化される。このように、PGC-1aの発現量の増加は、ミトコンドリアの生合成の増加と並行して、有酸素運動能力の向上、脂肪酸酸化の増加、筋肉疲労の遅延、有酸素運動のパフォーマンスの向上をもたらす可能性がある。これまでのセクションでは、PGC-1aがNRF-1/2とTFAMを活性化し、mtDNAの転写と複製を促進するミトコンドリア生合成のメカニズムを明らかにした。
同様に、PQQを補充することで、CREB-PGC-1a活性化経路のミトコンドリア生合成への影響を増加させるというデータもある[19]。このように、持久力のある運動とPQQの補給を綿密に組み合わせることで、エルゴジェニックな効果を得ることができると考えられる。しかし、現在、運動刺激下でのPQQの役割を検討した研究はないようである。また、PQQを摂取することで、ミトコンドリア・バイオジェネシスの指標にどのような影響があるのかを調べたデータもない。そのため、サプリメントと運動関連の研究という未知の領域をさらに探求していく必要がある。ミトコンドリア・バイオジェネシスが酸化能力を向上させるという理論的裏付けに基づき、PQQの補給が有酸素トレーニングによる筋肉内適応の効果をさらに高めるかどうかを検討するために、今後の研究が必要である。