Contents
Molecules, Information and the Origin of Life: What Is Next?
www.ncbi.nlm.nih.gov/pmc/articles/PMC7917628/
オンラインで2021年2月14日公開
サルバトーレ・チルンボロ1,*、アントニオ・ヴェッラ2
ファリド・ケマート(学術編集者)
概要
生命はどのようにして誕生したのか、生命とは何なのか、その根底には何があるのか。生命の質感は、分子とその化学的・物理的法則に支えられていることが知られているが、これらの疑問を徹底的に解明することは、科学にとって不可解な課題となっている。
分子とその法則にのみ注目することで、生物学や医学に対する機械論的(還元論的)な視点が間接的に科学知識の中に定着してしまったのである。これは、実験科学の長い歴史の中で起こったことであり、その後、生命の起源とその維持に関する多くの論文や推測の始まりに影響を与えた。
実際、生命とは何かを定義するには、新しい認識論が必要である。化学や物理学、さらには哲学の分野でもその起源が問われている生命システムの組織化が、人間の複雑な性質に焦点を当てるための新たな鍵となるような基盤が必要である。
このシナリオでは、情報の役割や水の構造などの多くの問題が、生命の起源に関する理論的基盤から長い間無視され、一種の舞台裏として疎外されていた。逆に、応用科学技術は、分子を唯一の主役と考えて進んできた。水の物理学や情報力学は、予想以上に生命システムの根幹に関わるものかもしれない。
生物の性質を機械論的に説明できるのか、それとも「何か他のもの」が必要なのか。おそらく、生命システムを単に複雑で高度に秩序立った機械と考える偏見を改めるだけで、生命についての健全な基礎を得ることができるであろう。
この原稿では、分子生物学や化学生物学の多くの基本的な側面を再評価し、プリゴジンの散逸構造や情報散逸(シャノン散逸)を含む新しいパラダイムで読み解いてみたい。そうすれば、情報散逸などの新しいモデルを参考にして、生物学や化学をどのように徹底的に見直すべきか、読者に洞察のヒントを与えることができるであろう。科学にとっての生命とは何かを解明するための真っ当な貢献に取り組むことができるのではないだろうか。
この概説は、単なる哲学的な推測ではなく、生物の生命の基礎を考え、記述する方法に深く影響を与え、この分野の読者に興味深い示唆を与えたいと考えている。
キーワード:生命の起源、シャノン散逸、散逸系、分子、ボルツマン、情報、複雑性
1. はじめに
生命の起源は、科学にとって基本的な難問である。生命に関する理論的な考え方の主要なテクスチャーは、分子とエネルギーのみを包含しており、何らかの方法で地球上に生命を誕生させることができたが、その方法についてはまだ議論されていない。分子とエネルギーがどのように作用して生命を誕生させたのか。さらに言えば、カオス、つまり我々の一般的な考えである「無秩序」と、複雑さ、つまり我々の一般的な考えである「組織化された秩序」だけが、生物界を構築する力だったのか?[1,2,3,4,5,6,7]. これらの疑問はすべて、「生命の起源」をめぐるあらゆる議論において最も重要なものである。生命を誕生させ、支える上での分子の役割は議論の余地がなく、このテーマを扱うために提唱された多くの重要な理論は、分子が生命システムを構築する能力にその信条を置いている[1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16]。分子がどのようにして生命を誕生させたのかを明らかにするために、研究者たちは、偶然と必然の単純な役割に基づいて会計処理を行い、生物学的原理に言及するのが常であるが[17]、しばしばデカルト的な方法で化学物質の質量とエネルギーを分離している。このような知的習慣の結果、人々はごく必然的に自然界をある種の「メカニズム」として読むようになってしまうのである[18]。生命システムを組織化された複雑な装置として記述すると、研究者は、生命の起源に関する議論において「終末志向の最終性」、つまり、ジャック・モノが「遠隔操作」と呼んだものに似た、ある種の目的や最終的な計画を示唆せざるを得なくなる[19]。abiotic principleは、必然性に駆られた最終的なタスクなしに、偶然性とランダム性から生命を構築する方法を概念的に説明する能力を持つべきである。例えば,ある著者によれば,開かれた反応ネットワークは,たとえ非触媒の二次反応からなる最も単純なモデルであっても,自発的に定常状態に達することができ,その結果,自己組織化能力を持つようになるという[20]。自己組織化は,必ずしもテレオノミーによって引き起こされるわけではない。したがって、一般的な直感は、興味をそそる実験設定を動かしながら、少ない熱力学的ルールで生命システムの自己組織化と自己再生の特性を説明する可能性について議論している。
しかし、生命は明らかに秩序立った計画なしに自己複製と自己組織化を行う不可解な能力を持っているので、生命がどのようにして誕生したかを明らかにするために、分子とエネルギーに起因する役割を再評価するには、例えば、原始スープによって形成された混沌としたシンクから自発的に出現する散逸のような秩序立った自己複製メカニズムを考慮するなど、機械論的な視点よりも、より動的な視点を包含する必要がある。実は、「生命とは何か」というのが、私たちが取り組むべき大きな課題なのである。生命には膨大なカオスの要素が含まれているので、メカニズムとしての生命という考え方は理論的には破綻しているかもしれない。しかし、その起源について機械論的な見方が主な信条であるならば、完全な定義はまだ解明されていない[14]。例えば、機械論的な見方では、メカニズム自体を記述するために原因と結果が完全に必須であるのに対し、力学的な見方では、始まりや終わりと称されるものは別として、異なる状態における変化を単純に示し、評価することでシステムを記述することができる。生命の定義を明確にするためには、生命とは何かという私たちの概念を全面的に見直さなければならない。それは、これまで文献に記載されてきた生命システム内の分子に関するオーソドックスな生物学的な話にも影響を与える。おそらく、生命システムは、秩序立った、テレオノミック指向の装置に過ぎず、より適切には、「秩序立った」現実として保持されようと絶えず努力しているダイナミクスであるかもしれない。生命を徹底的に定義することは、哲学にも役立つが、この文脈では、外界生物学など生物学の多くの分野に洞察を与えることができるであろう。スクリプス研究所のジェラルド・ジョイスは、生命を、自己組織化および自己組立が可能で、さらにその進化がダーウィン式自然選択によって処理されるものと定義した[15]。この定義の難問の核心は、実は自己組織化にある。しかし、なぜこのようなことが起こるのであろうか。
何がこの自己組織化・自己組立の特性を促進し、支え、生成するのかについての明確で健全な定義は、まだ完全には解明されていない。この不可解な問題に取り組んだ研究者もいるが、真の意味での成功はまだない。マトゥラーナの「オートポイエーシス」という概念に関する先駆的な研究[21,22,23]は、ごく最近の論文[22,23,24,25]でも紹介されているが、生命がどのように「自己表現」しているかについて、興味深い洞察をもたらした。しかし、マトゥラーナは、哲学的には平板なところに立っている。研究者たちは、生命がどのようにして発生したのか、また、化学的・物理的な性質を持つにもかかわらず、なぜ生命は結晶とは全く異なる存在として現れるのかについて、生物学に基づく理想的な理論をいまだに模索している[26]。「生命とは何か」という考えを特徴づける「自己複製」と「自己組織化」の概念と並んで、いくつかの論争的な問題が発生することがある。例えば、ウイルスやプリオンは、自己複製と自己組織化の能力を備えているにもかかわらず、生命システム、すなわち生物とは正しく見なされない。したがって、生命とは自己複製する物体以上のものである。典型的な「構造」モデルが、生命の理想的な「機能」概念に加えられた。例えば、疎水性膜としての独立したコンパートメントの存在や、恒常性維持メカニズムの発生などである[12]。これは、生命のダイナミクスが、化学的に駆動する単純な散逸よりもはるかに複雑であることを示唆している。私たちは、鉱物の石や結晶から生命体を完全に見分けることができるにもかかわらず、最も不可解な問題は、化学分子がどのようにして不活性な結晶体を形成するのか、あるいは、代わりに動的で自己組織化する生命システムを形成するのか、ということである。不活性の化学物質を生命体に変えることができる「マクスウェルの悪」とは何か?関連する反応はまだ生の化学や物理学の範囲内なのか、それとも何年も前にシュレーディンガーが挑発的に提案したように、凝縮物質のある種の「精神性」や「新しい物理学」を模索すべきなのか?[27].
私たちにはわからない。おそらく生物学は、新たな視点、新たな認識論から再出発しなければならない。「生命」として進化することができる何かを生み出すことができる「能力」の最深部の質感を記述する方法を再構築しなければならない。自然現象の化学的・物理的解釈のモデル化の方法を徹底的に見直し、より適切な生命の概念に光を当てることができる新しい魅力的な概念を導入しなければならないのである。生命を最も深い意味で定義するためには、どのような知的手段を用いればよいのであろうか。
実際、生命を記述する最も単純で一般的な方法は、複雑なシステムの進化を解明できる多くのモデルを生み出した統計化学から生まれた機械論的な視点である[28,29]。分子を抽象化して考える場合、通常は、完全に等しい一連の粒子が、熱対流力とブラウン運動のために自由に相互作用し、互いに移動している様子を描写する。このような現象が起こる事象空間を「ボルツマン空間」と呼ぶことができる。このボルツマン空間では、粒子(分子)の自由度が最も高く、「理想的な」ボルツマン空間と言えるが、自然界ではこの状況の生の近似値しか観察できない。この抽象的で理想的な化学統計学の世界では、分子は完全に同じ自由度を共有する粒子であり、この状態を「等方性ボルツマン空間」(E空間)と呼んでいる。E空間は自然界に存在することは非常に難しいのであるが、結晶性のモノマーや単純な分子、原子をボルツマン空間で考えると、おそらく発生する可能性がある。通常、E空間は「理想的な」事象空間であり、理論的な気体の熱力学系に非常に近いものである。逆に、それぞれが異なる粒子で構成されたボルツマン空間を「ディスキソトロピック・ボルツマン空間」(δίσ-「異なる」)またはD空間と呼ぶ。簡単に言えば、D空間とE空間を合わせて「等方性ボルツマン空間」と呼んでいる。D空間は完全な一意性を持った粒子でできている。この区別は、この原稿でさらに取り上げようとしている、生命の起源における偶然性と必然性の概念を紹介し、議論するために非常に重要だ。
偶然性と必然性は、科学的には相反するものであるという意見があるにもかかわらず、依然としてオートポイエティックな生命の基本的なエンジンであると考えられている[30]。モノッドの偶然性-必要性モデルは、宇宙とその誕生の概念における「無」の考えに劣らず、生命の概念を表現する我々の方法を形作ってきた[31,32,33]。多くの著名な意見によれば、物質もエネルギーも無から生じた、すなわちエクス・ニヒロである[33]。私たちが推し進めたい「無」という概念は、私たちの論文に関するさらなる重要な概念を説明するための序文として使われているに過ぎない。「何もない」というのは、とにかく曖昧な概念でさえあるかもしれない。「何もない」というのは、些細なことかもしれないが、「何もない場所」(つまり、何もない状態、これをN0空間と呼ぶ)であったり、もっと厄介な仮定として、「意味のないもの」(Nx空間)であったり、つまり、「何もない」というのは、完全に空っぽの場所であったり、完全に満たされているのに役に立たない場所であったりする。この概念的な枠組みは、生物学者として、生命に関する新たな重要な推測に取り組むための基本的なものである(下記参照)。
2. 偶然性、必要性、そして散逸。「無」、「無意味」、「異方性」、「端の向き」の概念
2.1. 無(N0空間)「無意味」(Nx空間)E空間、D空間。生命が誕生・進化するために必要なコンテクストは?
無、D空間、E空間といった概念を導入することで、生命がどのようにして誕生したのかを論理的に説明することができる。
無とは、何もない空間(N0空間)のことである。哲学的に考えれば、N0空間が理論的に生命が発生した場所であるはずがない。したがって、生命が「無」から始まったと主張する場合[34]、この「無」をもっと明確にすべきである。実際のところ、私たちは「無」を、意味や意義を持たないものと定義することに慣れている。「意味」という単純な言葉で単純に意味を持たないのではなく、「意味」を持たない物体でいっぱいの可能な「空間」である。センスは「オリエンテーション」の同義語として解釈され、何らかの形でベクトル的なアドレスが意図されていると考えられる。生物学は、方向付けられた目標、つまり「終わりの方向性」としての「意味」を持つことができる。なぜなら、生物学的システムの下にある化学的・物理的現象は、他のあらゆる自然システムと同様に、エントロピーが増大する不可逆的な力によって動かされているからである。物質とエネルギー、ひいては生命の主要な源としての「無」の概念を考慮に入れたい生物学者は、「無」は単に「何もない」(N0空間)のではなく、逆に「意味」のない要素(Nx空間)が密集して存在している可能性があることを認識すべきである[34]。
「ナンセンス」という概念は、通常、情報の秩序ある考え方とは対照的に、遠近法的な観点から誤って解釈されている[35]。セオドシウス・ドブジャンスキーの有名な文章「生物学では、進化の光の中でなければ意味をなさない」[36,37,38,39,40]以来、発展する生命体の動くエンジンは、自然選択の進化の過程であった。このモデルでは、「外部」である自然が、必要に応じて偶然を形成し、複雑さの調整、すなわち編成された機能的秩序を増大させることができると考えている。感覚の概念は、偶然と必然が生命を始めたときの役割に深く影響する。というのも、「感覚」は、単に「方向性」を意味することもあれば、力学や運動を扱う何かを意味することもあるのであるが、常にテレオノミーの概念と誤認されてきたからである[37]。簡潔に言えば、私たちのモデルでは、サシの「sense」は最終的な目的(the teleonomy)のないベクトル的な向き(「end orientation」)を意味する。オリエンテーションという言葉は、「動き」という概念の中に隠れているが、これは一般的に予想されているよりもはるかに興味深く、基本的な概念である。なぜ私たちは、無、終着点、そして動きの話をしているのであろうか。私たちは、これらの特徴が生命の根幹にあると信じている。実は、このプロセニアムをさらに解明するために、目を見張るような例がある。
真っ白でシンプルなキャンバスで描かれた絵は、N0空間、つまり「何もない」と考えられる。一方、非常に混雑した生々しい色の点々で描かれた絵は、Nx空間、つまり「意味がない」と考えられる。これは、絵が正しい方向に配置されていても、あるいは逆さまになっていても意味がないからである。このような状況は、絵が完全に等しい形で等間隔に描かれている場合にも得られる(E空間を想起させる)。これらの例は、生命がどのようにして始まり、その奥にある生命とは何かを定義する概念的な方法における3つの異なるスタートポイントを表している:1)粒子のない事象空間、2)D空間、3)E空間。キャンバス上の「無」は、真っ白なキャンバスや完全に均一な黒いキャンバスで無頓着に表現できる。逆に、真っ白なキャンバスや黒いキャンバスのどこかに赤いシミをつけると、絵に動きが出て、壁に貼る向きによって絵が違ってくるという特徴が生まれる。実際には、等方性空間にも、Nx空間にも、ある種の「多様性」を導入し、それを「異方性」と呼ぶことで、絵画全体が「動く」という認識を得たのである。この非常にナイーブな例は、生命の「始まり」において、異方性が進化の道筋を始めたのではないか、という視点で紹介したいと思う。異方性がどのように生命の誕生に貢献したかを説明すると、次のようになる。E空間に「ダイバースティ」が出現すると、どのE粒子に対しても異なる自由度を持つ異方性粒子が生成され、複合体を形成する確率が高まる事象のカスケードが発生し、それが進化の道筋となる。おそらく、E空間に異方性のある要素(「多様性」)が生じたときに、生命が誕生し、進化したのであろう。このモデルは、E空間によって作られた「Nx空間」を「何か」が乱した場合にのみ機能する。
私たちは、この推測の物語をさらに発展させようとしているのであろうか?
E空間は、理論的には等しい粒子でできており、それらの粒子は相互作用を受けて結合し、初期ペアを形成することができる。同じ特徴を持つ粒子(等方性)は、結合する確率も離脱する確率も同じなので、偶然に粒子同士が結合したり離脱したりする確率が同じであるため、発展途上のシステムを構成することはできない。このように考えると、このE空間は、進化的な意味での「Nx空間」と考えることができる。「異方性」を導入することで、E-Dペアを形成する確率がE-Eペアとは完全に異なり、さらに複雑な相互作用を形成するきっかけとなり、最高の組織化に向けてシステム全体の進化の道筋を劇的に変化させることができる。
複製される事象のない理想的なE空間から出発して、進化の経路は必然的に最終的にD空間に到達する。D空間は、完全に、あるいはわずかに異なる要素が空間内に存在することで完全に特徴づけられ、この空間を「独自性の空間」と定義することができる。この概観の時点では、単純に、あるいはその異方性のみに基づいて、進化のシステムを推し進めることができる粒子はない。なぜなら、システム内のどの粒子も、少なくとも理論的には、それをする可能性があるからである。そして、システムは、進化の方向からブロックされ、進化の用語では、「無」(Nx空間)になる。
E空間の情報負荷は、1つの高度に反復された情報によって表され、したがって最も低いシャノンのエントロピーを持つ(それは、非常に大きな声で正しく発音され増幅された1つの単語に似ている)。一方、D空間の情報負荷は最も高いシャノンのエントロピーを持ち、異なるユニークなメッセージの混雑した集団に匹敵する。E空間の進化の道は確率によって妨げられ、D空間の進化の道は情報性によって妨げられる。
しかし、進化の過程で、秩序が形成されることがある。これはどのようにして起こるのだろうか。この概念については後述するが、秩序は、複雑さを伴いながらも、偶然性と必然性にのみ基づいているならば、D空間によって形成される「Nx空間」の最終的な達成を避けることはできない。これは説明できる。
秩序が「偶然に付随する可能性」を変化させることを示す、理論的根拠に基づく可能な計算を試みることができる。
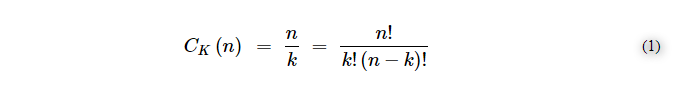
ここで、n = 1000 e k = 2であり、順序や繰り返しがない場合の組み合わせの概算値は≈499,500個となる。順番を導入すると、次の式(2)のようになる。
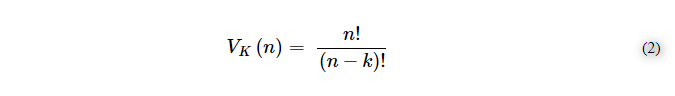
そして、結果として、繰り返しを含む場合には、式(3)を含む≒999.000-1,000,000のバリエーションになる。
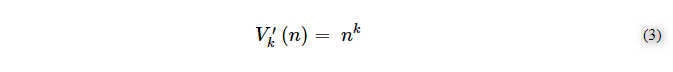
1000個の粒子の量は、正しい組み合わせ値を予測するにはあまりにも多すぎる。ボルツマン空間で動き、相互作用する粒子が21個しかないとすると、式(1)に従って、k=1の場合の組み合わせCは21個、k=2の場合はC=210個、k=3の場合はC=1330個となる。順番を導入するk=3では、バリエーションは7980となる。秩序は組合せ過程の中の変動数を増加させるので、変動数としてシャノンエントロピーを増加させることになる。つまり、秩序が偶然と必然によって形成されるならば、D空間内の変動性はシャノンエントロピーを増加させ、Nx空間に到達する可能性を高めることになる。
このように、偶然性と必然性のみを考慮してダイナミックな進化を開始する方法を推測的に説明することで、異方性が生命のエンジンに点火するものであっても、散逸のような更なるダイナミクスなしには進化の道を維持できないことを評価する機会を与えてくれる。
2.2. 消失のエンジン シャノンの散逸
散逸とは、内部の秩序を構築するために、複製と反復のメカニズムを導入するダイナミクスである。それでは、「プレバイオティック・スープと呼ばれる化学物質が密集した環境から、どのようにして生命が誕生したのか」という未解決の問題に戻ってみよう。
熱力学のバランスの中で働き、不可逆性を持つシステムは、熱力学の第2原理に従って、物質とエネルギーがフラックスに入り、熱で分解された物質がフラックスから出る、物質とエネルギーの複雑なフラックスと考えることができる。これらは開放的で不可逆的なシステムであり、熱力学的平衡から遠くないところで働き、エントロピーを増加させている。一方、生命力学は熱力学的平衡から遠く離れたところで働き、システム外の総エントロピーも同様に増加させている[41,42,43,44]。生命が自己組織化や自己複製を行い、複雑さを増していくと同時に、環境から「学ぶ」能力を獲得していく理由を説明するために、多くの試みがなされた。MaturanaとVarelaによる概念的枠組み[45]とは別に、ごく最近、Jeremy Englandは「散逸駆動型適応」に基づく進化モデルを提案した。その理論的テーゼは、生命の物理的な自然発生、すなわち生命起源の統計物理学に基づいている[46]。彼の理論では,分子は自由エネルギーをよりよく取り込み,熱を放散するためにランダムな方法で自己組織化することができるので,複雑で進化的なプロセスを自発的に生み出すことができるという[46]。通常,周囲の水クラスターのメゾスコピックな配列によって利用可能となる無視できる量の自由エネルギーを利用して自己組織化するという概念は,科学において特に強調されている[20]。イングランドのモデルは魅力的であるが、分子と水のナノ環境という非常に密接な境界に限定されており、「押し付け」のダイナミクスがないため、進化論的な意味合いに欠ける。私たちの考えでは、進化の過程につながる最終的な方向性に従うためには、「何か」が熱だけでなく、マクロステートにおけるエントロピーの増加に伴って得られる情報の負担も解消する必要がある。興味深いことに、Englandは進化の形成者としての環境を退け、単純な散逸駆動型のテレオノミーのみを考慮しているが、結論として、その魅力的な視点にもかかわらず、単純な統計的ダイナミクスは、「無意味な」自動散逸型の熱力学的システムでは、いつでも停止する可能性がある。生き物におけるテレオノミーの不可解な問題に対する可能な解決策は、ある「一定」のダイナミクスを考慮するだけで、進化の過程における偶然性の寄与を大幅に減らすことである。実は、これには散逸的なメカニズムが必要になるかもしれない。
散逸は、ノーベル化学賞を受賞したイリヤ・プリゴジンによって生まれた概念であり、生命の熱力学を含む多くの生態力学モデルで広く用いられている[47,48,49,50,51,52,53]。基本的に、散逸系とは、熱力学的平衡から外れて動作する熱力学的不可逆系であり、自己組織化臨界(SOC)と呼ばれる臨界状態に向かって自らを駆動する能力を備えている。SOCは、揺らぎから始まるアクションの雪崩を引き起こす特徴であるが[54]、生体などの熱力学系では、SOCは特定の条件でのみ発生する状態である[55]。散逸系の化学的な例としては、ベナール-レイリー細胞やベローゾフ-ジャボチンスキー反応のような興味深いモデルがあり、これらは基本的にある種のパターンの周期的な繰り返しによって特徴づけられる[56]。さらに、繰り返しや複製は、ほぼ一定のダイナミクスであり、生命のオートポイエティックなメカニズムの基本的な特徴でもある。したがって、一部の研究者は、生命は少なくともそのミクロの状態では、基本的に散逸的なシステムとして始まったのではないかという仮説を提案している[57,58]。興味深いことに、このレベルでは、ボルツマンとシャノンはエントロピー現象において比較可能な法則を共有している。というのも、1870年にルートヴィヒ・ボルツマンとJ.ウィラード・ギブスが統計的物理化学で確立した熱力学的エントロピーの数学的解釈は、20世紀の40年代にクロード・シャノンとラルフ・ハートリーが開発した情報エントロピーに似ているからである。
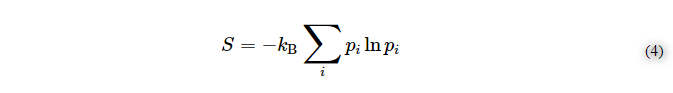
ここで、Sはボルツマン・エントロピー、piは平衡状態にあるミクロの状態の確率であり、次の式に似ている。
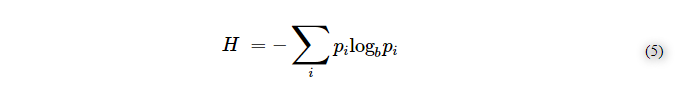
ここで、Hはシャノンのエントロピーであり、piはシャノン空間における定義されたメッセージの確率である[59]。しかし、このことは、マクロステートレベルでは、活発な散逸系において、ボルツマンのエントロピーとシャノンのエントロピーが全く逆の振る舞いをするということを意味するものではない。プリゴジンの散逸力学の根底にある熱力学と情報との密接な関係は、おそらく物理的自然の中で生命が進化してきたことの核心的な意味を表しており、今後はこのボルツマンとシャノンの調和した関係が、生命を「説明」する新しいモデルを試みる我々の道筋となるはずである。この文脈では、例えば、情報の変化が熱力学的な結果をもたらすことが明らかになったマクスウェルの悪魔のシナリオに対するシラードのモデルを思い出したいと思う[60,61]。生命が散逸的な現象として現れるのであれば[48]、ボルツマンとシャノンの関係は、ミクロ状態よりもマクロ状態に関わるはずである。化学的な散逸構造が、どのようにして目的指向の経過に向かって進化し、進化的なシステムを生み出したのかを解明する目的で、情報が発達過程全体で主要な、傑出した役割を果たしたことが推測されている。散逸系の最も顕著な巨視的側面の1つは、パターンの反復であり、これは、形態におけるチューリング・パターン、例えば、ベナール細胞(生物学的細胞のようなもの)やベルソフ・ジャボティンスキー振動(生物学的システムにおける多くのカオス的シグナル伝達経路のようなもの)に例示されている[62,63]。生物は、複製されたメカニズムによって完全に特徴づけられており、複製は生命システムの深い特徴である。我々の論文では、複製は、散逸メカニズムのためにボルツマン空間で発生する以外に、シャノンエントロピーにとっても基本的な目的があると考えている。
複製が異方性空間で起こると、ボルツマンはそのエントロピーを増加させるが(それぞれ自由度を持つ粒子がシステムに追加される)シャノンは「単一」のメッセージが高度に反復される、すなわち増幅されるため、そのエントロピーを減少させる。この現象を簡単に説明するには、シャノンのエントロピー方程式を見てみるといいだろう。
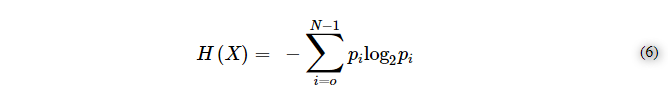
例えば、ヨーロッパ(イタリア)の言語のアルファベットのように、21個の異なる記号からなる文字列を考えると、シャノンのエントロピーは特に高い(H(X)=4.39)。異なる記号は、D空間を例示することができる。同じ空間に、ある特定の記号、つまり21個の記号(文字)の中に「a」という文字が5回複製されていることを導入すると、H(X)は3.74に減少する。また、21個の文字の中に10個の「a」を複製した場合、H(X)=2.95,15個の「a」を複製した場合、H(X)=1.60,20個の場合、H(X)=0.28となる。ボルツマン空間におけるある部分の複製(増幅)は、その空間におけるシャノンエントロピーを減少させる。
ここで、なぜ生物は自発的に散逸したカオス的なシステムの一部を複製するようになり、複雑性が増大していくのかという疑問が生じた。私たちは、可能な限りの説明を試みることができる。
生物の自然界では、複製されたものが「完全に」似ているわけではないという紛れもない現実を、誰もが観察することができる。したがって、例えば葉っぱのような定義された構造に集められた同じ情報の複製によるシャノンのエントロピーの見かけ上の減少は、事象空間でより多くの複製が行われ、入ってくるE空間により多くの「不完全さ」が追加され、等方性粒子に最終的な固有の「わずかな」ディスキストロピーを「与える」という意味で、ある程度の固有のエントロピーの増加を保持している。簡単に言えば、平面の木の葉は平面の木の典型であり、ハンノキやモミとは絶対に違うという事実にもかかわらず、定義された木のそれぞれの葉は、同じ木の中で、また同じ種の中の他のどの葉とも、わずかではあるがかなり似ていないのである。さらに、ボルツマン空間での複製は、散逸メカニズムを介して行われるが、内部に秩序を生み出しているにもかかわらず、基本的には熱力学第二法則に従わなければならないため、環境にエントロピーを放出する。外部の世界は生体よりもはるかに広く大きいので、増加するエントロピーの環境への放出は非常に容易である。システム外部のエントロピーの増加は、パターンの複製と秩序の創造を進めるようにシステムを後押しする[64,65,66]。この意味で、システムが高度に秩序だった状態に(何らかの形で)「凍結」されるまで、相反する力を散らし続け、より頻繁に、熱力学的平衡から遠くないところで動作し始める。そうなると、熱力学的な第二法則にしたがって、秩序が劣化することもある。いずれの場合も、システムは、過剰な秩序か、無秩序なカオスのいずれかにブロックされ、進化の道筋が見えなくなる。このような状況では、散逸エンジンが停止しているため、物体は「生きていない」ものに非常に似ているかもしれない。したがって、散逸システムの「内部」にある何か、つまり熱力学によって保持されているシステムが、複雑な秩序の増加とは別に、散逸マシンを作動させ続けるはずである。
私たちが強調しているのは、生物学において、散逸による自己複製は、生物を自然界に存在する他の秩序ある物体と区別するための絶対的な基本であり、それは高度な秩序(結晶)や無秩序(アモルファス状態)である可能性があるという概念である。複製は、散逸系において定義された情報的能力を持っている。複製は、熱力学的(ボルツマン的)散逸から生じる単なる秩序的なものではなく、シャノン空間の主要な特徴である。しかし、ボルツマン空間では、外部からのエントロピーの増加が内部の秩序の生成を押し進めるのに対し、シャノン空間での「隠された」エントロピーの増加は、我々が「散逸的散逸性」と呼んだもので、システムの「内部」で発生するため、システムを内部から継続的に散逸させることになる。簡単に言えば、ボルツマン空間でコンポーネントが複製されている間、シャノンエントロピーは減少しているが、同時に、ディスキオトロピーの下向きの増加により、正のシャノンエントロピーのシンクが形成され、システムの散逸ダイナミクスの再起動を余儀なくされるのである。このような散逸により、システムは「生き続ける」ことができ、ボルツマン散逸は進化的な複雑さに向かって「進む」ことができるのである。
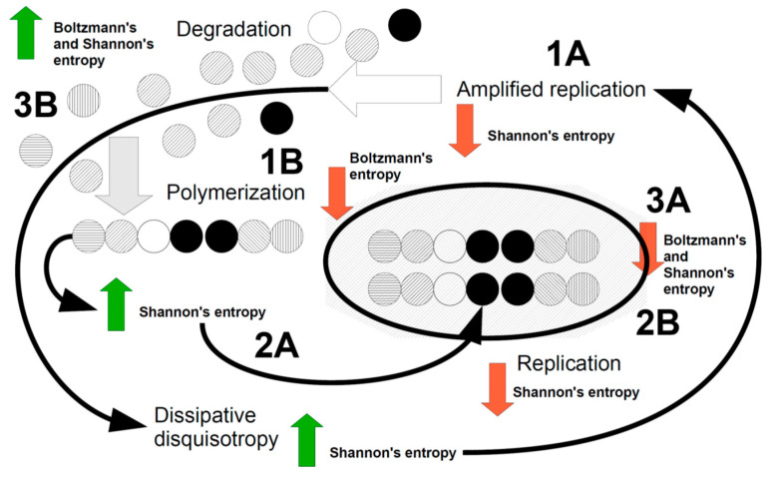
図1はこれらの点をまとめたものである。
図1 生物を表す主要な力学的事象を示した漫画
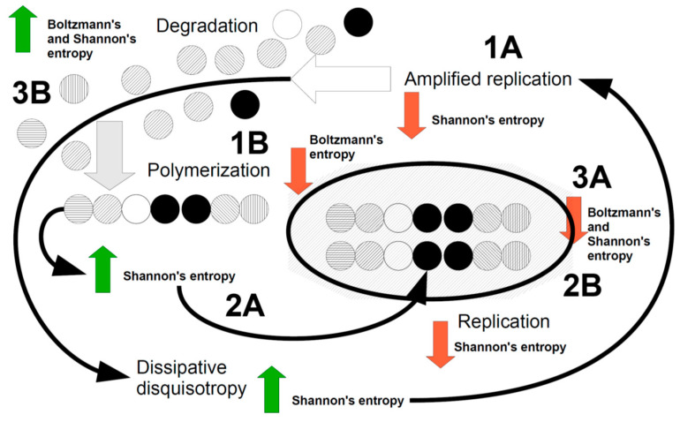
散逸機構は、相反する力を散逸させるために、構造や機能を複製し(1A)通常はボルツマン空間のE成分を増加させ、シャノンエントロピーを減少させる(赤矢印)。このエネルギーは、水やロックダウンられた水のメゾスコピック構造の再編成を介して、消散そのものによってもたらされる。複製は、ボルツマン空間内の単一の構成要素、例えば単一の分子や単一の細胞を対象とすることができる(増幅)。複製はボルツマン・エントロピーを増加させるため、熱力学的な散逸がボルツマン空間の要素を結合させ(エネルギーは散逸メカニズムから得られる)ポリマーを作り、それがシャノン・エントロピーを増加させる(緑矢印)(2A)。ポリマーが結合してマクロ構造になり、より複雑な物体になると、ボルツマンエントロピーとシャノンエントロピーの両方が減少し、複雑さが始まる(3A)。この段階では、分解が起こらなければ(3B)システムが停止してブロック化されてしまうかもしれない。ボルツマンエントロピーとシャノンエントロピーの両方が増加することで、サイクルが再開される。この段階のエネルギーは、熱力学(ボルツマンの場合)と、どんな複製イベントにも「不完全さ」が含まれるため、複製に伴う散逸性の増加(1A)によって与えられる。この「散逸的な異方性」は、シャノンが再び複製を行うことでエントロピーを減少させ、生命システム全体の散逸ダイナミクスを促進させる。
図1を説明してみよう。
おそらく、私たちは最初、ボルツマン空間とシャノン空間の両方が高いエントロピーを持つ異方的な事象空間にいると思われる。このような状況下で、定義された粒子、サイクル、パターンの複製が行われると(図1のポイント1A、増幅複製)シャノンのエントロピーは減少するが、ボルツマンのエントロピーは増加するため、ポリマーを作るなどして、このエントロピーを減少させる必要に迫られる(図1のポイント1B)。ポリマーは、散逸的なディスコトロピーと同様に、その組み合わせの多様性のためにシャノンのエントロピーを増加させ(図1,ポイント2A)その状況は、シャノンにポリマーの複製をさせ(図1,ポイント2B)ボルツマンにそのエントロピーを減少させるために、複雑さを形成するポリマーやマクロ構造を結合させることを再び義務付ける(図1,ポイント3A)。この時点で、ボルツマンもシャノンもエントロピーを減少させているが、ボルツマンは熱力学の第2原理にしたがって、劣化した物質を放出しなければならない(図1のポイント3B)。これにより、オーケストラによるボルツマン-シャノンの散逸システム全体が再起動するはずである。熱力学的フラックスの中で相反する力を散らす必要性が、ボルツマン空間の特徴として認識されていることが「末端指向」と呼べるなら、シャノン空間でも同じことが考えられるはずである。言い換えれば、生物システムはシャノン散逸のおかげで、「オートポイエティック」な本質的な特徴の中で生き続け、前進しているのである。私たちがここまで進めてきたこの推測的な論文は、情報(シャノン)が熱力学(ボルツマン)を形成する能力に言及したシラードの論文やToyabeらの証拠[60,62]に合理的な根拠があるかもしれない。生体内部の組織化された水は、疎水性ポケットの寄与があったとしても、そのメゾスコピックな再配列を介して、ボルツマン散逸を情報の変化に結びつけ、生体の情報的負担の散逸のようなものをもたらしたと推測される。私たちの仮説は、上述した編成機構(図1参照)が、生体内、細胞内、ロックダウンられた水によって編成され、移動するのではないかというものである[67,68,69,70,71]。
3. 複雑さから始める 初期生命から複雑な生物まで
3.1. 複雑さはどのようにして生まれたのだろう。自由度、関係性、記憶の概念
これまでに、ボルツマン系とシャノン系という2つの散逸系が絡み合って、生物のダイナミクスを最終的な方向に向かって進めていく上で、基本的な役割を果たしていることを紹介していた。複雑さとは、エントロピーの減少から最終的に上昇する秩序の産物であると定義されている。根本的には、生命がどのように進化してきたかを概観する際に、秩序と同様に複雑性も必ずしも本質的なテレオノミー(目的論説)を持っているとは限らない。それは、生命システムが感じている散逸の単純な必要性から生じているのである[72]。複雑性についての最も広範な考え方には、規則性の概念が含まれている可能性があり[73]、したがって複雑性は、複製がシャノンエントロピーの減少と関連しているように、必ずしもシャノン散逸から生じる秩序化された産物でなければならない。生物学では、還元主義の概念を克服することが大きな関心事であったため、複雑性の概念に対処するための新しいモデルが提案されていた[74,75,76]。根本的には、「オミック」な視点で複雑性を読み解き理解しようとする考えには、間違いなく遠近法的なビジョンが含まれていなければならない。というのも、最終的な機能的目的がなければ、動的ネットワーク全体の同じ解釈は意味をなさずに突然失敗するからである。我々の考察は根本的に逆で、複雑性は散逸的な副産物として生じ、同様に秩序も生じ、その遠隔操作はおそらくこの段落で議論する不可解なメカニズムの結果である[77]。
図1の概要を考慮すると、散逸的なシステムが化学的資金によるメカニズムであることをやめ、生物学的なシステムになるための「閾値」は、分解に対する複製の割合が増加することであるかもしれない。
再び、私たちの魅力的な歴史が起こったイベント空間に目を向けると、最初に、相互作用と関係性の間で区別しなければならない。ボルツマン空間の2つの粒子は、相互作用によってそれぞれの自由度が変化したときに関係を持つことになる。この出発点は、進化の歴史の中で複雑さが生じた理由を説明するために重要だ。基本的には、相互作用する2つの粒子は、それぞれボルツマン空間内で自由に動き、相互作用することで最大の自由度を持っている。それらの相互作用が、参加している各粒子の自由度を変化させるとき、「関係」に到達することができる。しかし、理論的には、どの粒子も自分の自由度を失いたくないと思っているので、ペアの自由度が単独の粒子よりも高い場合にのみ、関係が形成される。この「強制力」は、自由度の「獲得」がかなり情報的な性質のみであるため、シャノン散逸によって引き起こされる。この「強制力」は、自由度の「増加」がかなり情報的な性質だけであることから、シャノンの散逸によって引き起こされることがある。秩序化された複雑さの「内部」での自由度の増加が、複雑さを成長させるエンジンであるため、さらなるシャノン散逸が起こっても、連続的な情報形成が保持され、その結果、我々が「テレオノミック」または「最終的な終わりを志向する」と分類する複雑さの「種類」につながるのである。関係は、複製とシャノン散逸によって、それらの関係が複雑な構造や機能に結合されると、散逸的なダイナミクスを伴って現れる(図1を再度参照)。しかし、この関係は、情報の概念の基礎でもある。物体はそれ自体が情報ではなく、「他のもの」と「関係している」場合に情報となる。コップは飲み物を飲むためのものであるが、砂を集めるためにも、心地よい音を出すためにも、紙に円を描くためにも使うことができる。「グラス」という情報は、単にその化学的性質や形状、用途によって定義されるものではない:私たちはグラスが何であるかをよく知っているにもかかわらず、それは「依存」している。したがって、力学に「関連する」出来事だけを情報とするならば、「記憶」とは、その反復された形である。記憶とは、収集されたファイルや一定の固定された物体ではなく、再帰的な(複製され、増幅された)情報である。
3.2. いくつかの理論的・推測的帰結
つまり、初期の生命は、シャノンの散逸メカニズムが発生した時点で発生したのである。しかし、散逸的な化学システムがいつ生物進化システムに変わるのか、すなわち散逸的なシステムにシャノン散逸が生じて、生命という終末指向のダイナミクスが始まるのか、今のところ確認できていない。この分野の既存の文献に基づいて、いくつかの推測されるアイデアを提示したが、さらなる洞察が必要である。無機物で構成された散逸系が生命体に進化するための「閾値」はあるのか?いくつかの仮説を提示したが、まだ推測の域を出ない。最も可能性が高いのは,ボルツマン空間内のディスキストロピーのレベルが,この空間を等方的な空間に変化させるシステムの能力に打ち勝ったときに起こるかもしれないし,もっと単純に言えば,プレバイオティック・スープにもっと複雑な分子が現れて,はるかに多くの散逸的ディスキストロピーが起こりうるときに起こるかもしれない[78,79]。私たちにはわからないが、複製が分解に打ち勝った場合にのみ、この現象が起こるかもしれない。地球以外の太陽系でもプレバイオティック・スープが観測されたことを考慮すると[80]、生命とは何かを問うことは、現代科学にとって、いまだに端的で重要なテーマである。
結論として、私たちの推測によれば、生命は化学的・物理的な散逸システムとして誕生した。ここでは、鉱物、粘土、水が最初に触媒反応を起こし、次に散逸メカニズムを起こし、後にボルツマン/シャノンが編成した散逸を起こした。シャノン(または情報)散逸と呼ばれる種類の散逸が設定されたときにのみ、化学的散逸システムは生物学的進化システムに変わり、秩序と複雑さを生み出したのである[77]。この複雑さは、とにかく無限ではない。図1で説明したような複雑性は、シャノンによって「複製」されるはずである。したがって、生物システムがある程度の複雑さに「制限」できるかどうかは、熱力学の第2原理を考慮して、ボルツマンが全体として散逸を維持できるかどうかにかかっているのである。このことは、地球上の生物が、バクテリアから人間まで、最も幅広い複雑な生物を含んでいることを示唆しているかもしれない。ボルツマンとシャノンのある種のダイナミックなバランスが、自然界にバクテリアとクジラが無関係に共存している理由を説明しているのかもしれない。
4. 研究の限界
この「視点」の論文は、現在議論されているテーマについて、新しい提案をしようとしたものであるが、いずれにしても、まだ完全に解明されていない。私たちは、生命の起源についての議論における基本的な要点に焦点を当てたが、特に、健全な数学的アプローチのモデル化や、私たちの推測をさらに明確にすることができる実験データによって、さらに多くの洞察が必要であることを絶対に認識している。このような試みは、原因と結果の問題にとどまるが、「原因」と「結果」を求めることは、この文脈では特に難しいかもしれない。正確な原因とその影響が明確に特定されない限り、このテーマは推測の域を出ないが、オリジナルの提案を動かすためには、現時点ではおそらく必要なことだと思う。生命の起源に関する問題は、「原因はいったい何で、その結果は何であるか」という問題に還元されるべきであり、言い換えれば、原因と結果の起源である。これは本稿の決定的な弱点に見えるかもしれない。しかし、シャノン散逸の原因は何となく示されているが、このメカニズムを解明するには、さらに多くの知見を得る必要がある。現代物理学は、量子力学的な現実の定式化に深く入り込んでいるようで、マクロな現実においても、明確な原因と結果を特定することは究極的には不可能であり、少なくとも根本的には曖昧である。
5. おわりに
生命とは何か?数年前にシュレーディンガーが投げかけたこの巨大な問いは、ついに完全かつ網羅的な定義を欠いてしまったようである。近年、多くの基礎理論が報告されており、おそらく生命は、偶然のランダムな能力や必然の強制的なパルスだけではなく、おそらく情報と熱力学が密接に絡み合った散逸的なメカニズムによって、最終的には水によって保持されるカオス的なダイナミックさで誕生したのではないかと考えられる。このレビューでは、生命の起源に関する新しい理論に光を当て、この魅力的な分野に携わる読者が興味を持ってくれることを願っている。
