
Contents
pubmed.ncbi.nlm.nih.gov/32210840/
Homeostasis: The Underappreciated and Far Too Often Ignored Central Organizing Principle of Physiology
2020年3月10日オンライン公開d oi:10.3389/fphys.2020.00200.
pmcid:pmc7076167
PMID:32210840
要旨
2010年にFrontiers in Physiology誌の創刊号に掲載されたエッセイで述べられているように、生理学の壮大な挑戦は、分子から無傷の生物に至るまで機能を統合することにある。還元主義的なアプローチに由来し、それにますます依存するようになった膨大な量の情報を理解するためには、知的学問としての生理学を生み出した科学者たちが開発した伝統的な統合的かつより全体的なアプローチをより重視する必要がある。
ギリシャ神話に登場する体液の概念から、クロード・ベルナールの「内なる環境」、ウォルター・キャノンの「ホメオスタシス」概念の定式化、制御理論(フィードバックとフィードフォワード制御)の適用による内部環境の一定化まで、生理調節に関する理解は時代とともに発展してきた。ホメオスタシスは、生理学の中心的な統一概念となり、生物が変化する外的条件に適応しながら内部安定性を維持する自己調整プロセスと定義されている。
ホメオスタシスは、静的で変化しないものではなく、外的な課題を克服するために必要に応じて内部条件を変化させることができる動的なプロセスである。また、恒常性の調節は、単に単一の負のフィードバックサイクルの産物ではなく、より高い制御センターによって変更可能な複数のフィードバックシステムの複雑な相互作用を反映していることに留意することが重要である。
このような階層的な制御とフィードバックの冗長性により、より細かいレベルの制御と、変化する環境条件への適応を可能にする高い柔軟性が実現される。生物の健康と活力は、恒常性調節の最終的な結果であると言うことができる。
この概念を理解することなしに、正常な生理学を理解することはできない。逆に言えば、ホメオスタシス機構が破壊されることが病気につながるということであり、有効な治療法はこのホメオスタシス状態の再確立に向けられる必要がある。
そこで、このエッセイでは、ホメオスタシスに関する私たちの理解の変遷と、健康と病気における生理的調節と調節不全の役割を説明することを目的とする。
キーワード
生理学、ホメオスタシス、内部環境、クロード・ベルナール、ウォルター・キャノン、制御理論、フィードバック制御-負と正、サイバネティックス
はじめに
2009年11月、私はFrontiers in Physiologyという新しいオープンアクセス生理学ジャーナルを立ち上げることに同意し、2010年4月に論文が出版された。編集長としての私の任務の一つは、学問分野としての生理学が直面する最大の課題と思われるものを論じた簡潔な「グランドチャレンジ」論文を執筆することだった。この最初のエッセイの出版から10年が経過し、このグランドチャレンジの記事を再検討し、更新する好機となった。
生理学のグランドチャレンジ
2010年のエッセイで、私は生理学の壮大な挑戦は「分子から人間までの機能を統合すること」だと述べた(Billman, 2010)。言い換えれば、還元主義的アプローチに由来し、それにますます依存するようになった膨大な量の情報を理解することである。このことは、今日、生理学が直面している最も深刻な未解決の課題であると私は考えている。生理学という知的学問を生み出した科学者たちが開発した、伝統的な統合的アプローチやより全体的なアプローチをより重視する必要がある。言い換えれば、生理学者は今、原点に立ち返る時なのである。デ・ヴィンチの「モナリザ」やゴッホの「星月夜」の美しさを、絵の具を一粒一粒取り出して分析することで理解することはできない。また、健康維持のために様々な器官系がどのように機能しているかを、遺伝子や分子一つを調べることで理解することはできない。絵画を鑑賞するときと同じように、身体も全体を見て初めてその素晴らしさがわかる。このエッセイでは、ホメオスタシスという概念に焦点を当て、生理学という学問が構築された中心的な組織原理、すなわち、分子から無傷の生物まで機能を統合するために立ち返る必要がある概念に焦点を当てる。以下のセクションの一部は、以前、少し異なる形で出版されたものであり(Billman, 2013)、出版社の許可を得て再掲載している。

ホメオスタシス:定義
ホメオスタシス(恒常性)とは、変化する外的条件に適応しながら、生体システムが安定性を維持するための自己調整プロセスである、と現在のところ定義されている。この概念は、生物が、変化し、しばしば敵対的な外部環境に直面しても、適応し、生き残ることができるように、内部条件を多かれ少なかれ一定に保つことができることを説明している。ホメオスタシスに対する私たちの認識は、何世紀にもわたって徐々に広まり、生理学の中心的な組織的信条となった。この自己調整プロセスを理解しなければ、健康なときも病気のときも、身体の機能を完全に理解することはできない。ホメオスタシス機構が破壊されることが病気につながるのであり、効果的な治療は、自然に逆らうのではなく、自然とともに働き、ホメオスタシス状態を再び確立することに向けられなければならない。以下のセクションでは、ホメオスタシスに関する理解の変遷を説明し、健康と病気における生理的調節と調節不全の役割を評価することにする。
ホメオスタシス:歴史的な視点
真の安定は、想定される秩序と想定される無秩序が均衡を保っているときに生じる。真に安定したシステムは、予期せぬ事態を想定し、破壊されることを覚悟し、変革されることを待ち望む。
トム・ロビンス(アメリカの小説家、1936年生まれ)1
健康のためには身体の調節が必要であるという考え方は、古代ギリシャ時代にまでさかのぼることができる。ギリシャの医師であり哲学者であったクロトンのアルクメオン(紀元前500年頃)は、健康と病気を説明するために「対立物のバランス」と呼ぶべきものを提案した。彼は、健康と病気を定義するために政治的なアナロジーを用いて、次のように述べた:「健康とは、機能の権利の平等であり、湿潤と乾燥、寒さと暑さ、苦味と甘味、その他だが、いずれかの対の単一のルールは有害である。(と述べている(Freeman, 1948)。このように、権力の不平等は、政治体制では専制政治に、身体では病気につながる。この概念は、コスのヒポクラテス(紀元前460年頃~紀元前377年頃)によって拡大され、健康とは4つの体液、すなわち血液、痰、黄胆、黒胆のバランスと混合から生まれるものであると提唱した。彼はこう書いている:
健康とは、主にこれらの構成物質が、強さと量の両方で互いに正しく比例し、よく混ざり合っている状態のことである。痛みは、これらの物質のいずれかが欠乏または過剰であるか、または体内で分離して他の物質と混合されていない場合に発生する。(Chadwick and Mann, 1950)(チャドウィックとマン,1950)
こうして、医学は「引き算と足し算のプロセス:過剰なものの引き算、欲しいものの足し算」となったのである。(ジョーンズ、1923)。ヒポクラテスはさらに、治癒過程における自然の助けの役割(vis medicatrix naturae)、すなわち身体が自ら治癒する能力を認めた(Hall, 1975)。自然の摂理に従うように道を整えるのが医師の役割であった。この概念は、その後何世紀にもわたって、近代の幕開けまで、医学の基礎となった。
この「自然の治癒力」という概念には、生体の正常な状態が損なわれたときに、生体のサブユニットが協調的に作用して健康を回復するという前提がある。生理学とは、身体の各部分がどのように連携して健康を維持しているかを理解するための学問であり、その起源は16世紀に遡る。生理学という言葉は、1542年にJean Francois Fernel(1497-1558年頃、図1)によって初めて紹介された[De Naturali Parte Medicinae(医学の自然な部分について)]。病気の研究である病理学と区別して、健康な体の機能を研究するものである(ホール、1975)。ウィリアム・ハーヴェイ(1578-1657)は、血液の循環に関する記述で、慎重に設計された人体実験と動物実験を用いて、主要な身体器官系の機能を確立した最初の人物である。この生理学の応用は、彼の代表的な出版物「Exercitatio ANATOmica De Motu Cordis et De Circulatione Sanguinis in Animalibus」1628(ANATOmical exercises on the motion of the heart and the circulation of blood in living creatures, first English translation 1653)からの次の短い引用で説明されている:

図1 ジャン・フェルネル(1497-1558年頃)の肖像画
生理学という言葉を作った人物である。出典はこちら:国立医学図書館(医学の歴史パブリックドメイン画像ファイル)。
心室の拍動による血液が肺を流れて全身に送り出されることは、理性と実験によって示されている…動物の体内の血液は絶えず円を描いて動き回り、…心臓の作用や機能はこのポンプ作用を達成することにある。これが、心臓の運動と拍動の唯一の理由である。(ハーヴェイ、1628/1653)
その後、数世紀にわたって生理学の概念は発展し、生理学のさまざまな下位学問を統合する中心的な信条が生まれた。正常な身体機能を理解することで、病気の原因となる障害を特定し、最終的には健康な状態を取り戻すために修正することができる。
これまで見てきたように、身体機能の調節と制御に関する初歩的な理解は、紀元前6世紀のギリシャまで遡ることができる。その後、数世紀にわたって散発的な進歩はあったものの(Adolph, 1961)、系統的な生理学的調査によってこの概念が大きく前進したのは、19世紀になってからだ。生理的調節に関する現代の理解は、この分野における2人の巨人の肩にしっかりと乗っている:クロード・ベルナール(図2)とウォルター・キャノン(図3)は、それぞれ内部環境の恒常性とホメオスタシスという観点から調節を説明した。

図2 クロード・ベルナール(1813-1878)の写真。外的条件が変化しても生物は安定した内部環境を保つという「a fixité du milieu intérieur」の概念を提唱した。出典はこちら国立医学図書館(医学史パブリックドメイン画像ファイル)。

図3 ウォルター・B・キャノン(1871-1945)の写真
クロード・ベルナールの研究を基に、ホメオスタシスという言葉を作り、生物系が変化する状況に適応しながら安定性を維持する自己調整プロセスを表現した。出典はこちら国立医学図書館(医学の歴史パブリックドメイン画像ファイル)。
近代実験生理学の創始者と呼ばれるフランスの生理学者クロード・ベルナール(1813-1878)は、生命システムが、絶えず変化する外部環境に対して生体を緩衝し保護する内部安定性を有していることを、おそらく最初に十分に理解した人物である(Cooper, 2008).彼は、生体が体温や血糖値を比較的一定に保つために協調的に動作するメカニズムを有しており、この内部安定性が生体の健康にとって不可欠であることを認識した。彼は次のように結論づけた:「La fixité du milieu intérieur est la condition de la vie libre, independent”(Bernard, 1865) [内部環境の固定性(=不変性、安定性)は、自由で独立した生活の条件である]。見落とされがちであり、強調されるべきは、この記述においてベルナールは、「milieu intérieur」の安定性は、自由で独立した人生の結果(成果)ではなく、その先行条件(すなわち、必要条件)である、という新しく過激な仮説を提示していたことだ(ターナー、2017)。
ベルナールは、その生涯で最も有名なフランス人科学者として高い評価を受けたが、1854年に初めて明らかにした「内部環境の安定性は外部条件に依存しない」という仮説は、その後50年間、ほとんど無視された。Gross(2009)は、ベルナールの考えが発表されてから受け入れられるまでに時間がかかった理由として、(1) パスツールが病気の予防や治療に役立つ細菌学を発見し、生物学の研究を支配するようになったこと、(2) 進化論と一般生理学との間にギャップがあり、自然選択が制御の進化をもたらす手段であると理解するまで時間がかかったこと、(3) 内部環境の測定に必要な技術がまだなかったことをあげている。
しかし、19世紀末から20世紀初頭にかけては、バーナードの考えを、説明の中心概念として、また生理学の研究プログラムとして、多くの研究者が取り入れるようになった。バーナードの影響を受けたのは、イギリスのウィリアム・M・ベイリス、アーネスト・H・スターリング、ジョセフ・バークロフト、J・S・ハルデン、C・S・シェリントン、アメリカのL・J・ヘンダーソン、ウォルター・B・キャノンといった生理学上の著名人である(Adolph、1961、Cooper 2008、Gross 2009)。実際、スターリングは、一定の内部環境を維持することを表現するために、「身体の知恵」という言葉を作り出した(Cooper, 2008)。その後、ウォルター・キャノンがホメオスタシスの概念を紹介した著書のタイトルとしてこの言葉を使い、この言葉を広めた。1900年、ベルナールの弟子で後にノーベル生理学・医学賞を受賞したシャルル・R・リシェ(1850-1935)は、内部環境の動的安定性を強調した。次の引用文は、ウォルター・キャノンの定義に先立つものであった。
「生命システムは安定している…それは、それを取り囲む巨大な力、しばしば不利な力によって破壊、溶解、崩壊されないためである。しかし、一見矛盾しているように見えるが、生命システムは、外部からの刺激によって自らを変化させ、刺激に対する反応を調整することができる興奮性を持っている場合にのみ、その安定性を維持する。ある意味で、修正可能であるからこそ安定なのである。”わずかな不安定さは、生物の真の安定に必要な条件である。”(リシェ、1900)
この一定の内部環境(milieu intérieur)という概念は、アメリカの生理学者、ウォルター・キャノン(1871-1945)によって拡大された(Cooper, 2008)。彼は、ギリシャ語のǑμoις (hómoios) 「似ている」とστάσις (stásis) 「止まっている」を組み合わせてホメオスタシスという言葉を作り、生物系が環境の変化に適応しながら安定性を保つ自己調整過程を表現した。ホメオスタシス(恒常性)は、しばしば「変わらない」「停滞する」という意味に誤解されることがある。しかし、キャノンはあえてギリシャ語で「同じ」ではなく「似た」という意味の「ホモイオス」を選び、内部状態が変化しうる、つまり、似ているが同一ではない(安定しているが、生物が自由に適応できる値の範囲にある)という考えを表現した。つまり、ホメオスタシスとは、正常な状態や機能を乱すような状況や刺激に対して、各部が協調して反応した結果、システムが内部的に安定した状態を維持する傾向のことをいう。このように、ホメオスタシスという言葉は、2つの考え方を伝えようとしている:(1)ある範囲内での内部安定と、(2)この内部安定を維持するための協調的な動的反応(自己調節的な目標追求行動)である。1932年に出版され、大きな影響を与えた単行本『身体の知恵』から、次のように説明している:
「生物の定常状態のほとんどを維持する調整された生理学的プロセスは、脳や神経、心臓、肺、腎臓、脾臓など、すべてが協調して働く、非常に複雑で生物に特有であるため、私はこれらの状態をホメオスタシスと呼ぶことを提案した。この言葉は、何か決まったもの、動かないもの、停滞を意味するものではない。この言葉は、変化することはあっても、比較的一定である状態を意味する。(キャノン、1963)
キャノンが強調したように、ホメオスタシスは静的なものではなく、むしろ、変化する環境の要求に直面して生存能力を維持する動的な自己調整システムである。バーナードの言葉を借りれば、ホメオスタシスは生物に特有の性質であり、生命そのものに関与している可能性がある。より最近では、ターナー(2017)がホメオスタシスを動的な不平衡と表現した。動的とは、安定した内部環境が、自由で独立した生活が可能になるように、対立する力(彼が不平衡と呼ぶ)のバランスを維持するために、継続的に監視し調整(再び、自己調節プロセス)する必要があるからだ。さらに、「正しく理解すれば、ホメオスタシスは生命の基本的な特性であり、非生命と区別するものである」と述べている。「つまり、ホメオスタシスは生命なのである。」(ターナー、2017)と述べている。
ホメオスタシスのパズルの最後のピースは、システム工学の制御理論を応用して、生物系における自己制御を説明することによってもたらされた。物理化学的な内部状態の「恒常性」は、複数のネガティブフィードバックシステム(およびポジティブフィードバックシステム)の複雑な相互作用によって維持されている。これらの調節機構の相互作用は、システムの安定性を高めるだけでなく、冗長性(バックアップ)を提供し、ある構成要素が故障しても必ずしも破局に至るとは限らない。このように、生理学的研究は、その設立当初から、生物(微生物、植物、動物、人間など)を一つの機能的実体として理解することに向けられてきた。
フィードバック制御:ホメオスタシスの基礎となるプロセス
“Nam deteriores omnes sumus licentiate.”
統制のないところでは皆、退化する。
テレンス(ヘウートン・ティモールメノス、483行目)
これまで見てきたように、ホメオスタシスの重要な特徴は、生物の内部環境が自己調整(目標追求)システムによって狭い範囲に保持されていることである。ホメオスタシスを得るためのメカニズムとして、フィードバックとフィードフォワードがある。ここでは、まず、恒常性調節におけるフィードバックの寄与について述べ、次に、フィードフォワード(中央指令ともいう)機構について簡単に説明することにする。
フィードバックシステムとは、システムの過去の行動(内部環境の変化)の結果を(情報を介して)システムに送り込み、将来の行動を制御する閉ループ構造であり、システムは自らの行動に影響を与える(Forrester, 1976より改変)。フィードバックシステムには、目標を求め、その目標を達成できなかった場合に対応するネガティブフィードバック(安定した値域を維持する)と、その結果をもとにさらに大きな行動を生み出す成長過程(成長サイクル)を生み出すポジティブフィードバックがある。これらのフィードバックシステムは、それ自体がより高度な制御の対象となる。つまり、環境刺激に対する行動応答をサポートするために、調節された変数の動作範囲を調整することができる。ホメオスタシスは、複数の負と正のフィードバックシステム間の複雑な相互作用と競合の結果であり、生理学的調節の基礎を提供するものである。
再び、自己規制システムの起源を古代ギリシャに遡ることができる。
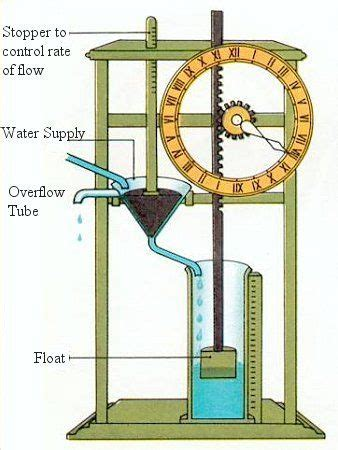
自己制御の原理を用いた最初の装置は、アレクサンドリアのクテシビオス(またはクテシビウス、ギリシャ語ではKτησίβις)が発明した水時計(clepsydra)である(前285-222)(Landels 2000)。水時計は、安定した水流によって時間の流れを計測するものである。水位が比較的一定でない場合、水流は時計に供給する水柱の高さによって変化する(容器が満杯だと速く、容器内の水位が下がると遅くなる)。クテシビオスが設計した水時計は、時計の水槽の水位を一定に保つために、フロート弁(現代の水洗トイレに使われているものと同じ)を使用していた。水位が下がるとフロートも下がり、弁が開いて時計の貯水槽に水が流れ込み、水位が補充される。そして、水が所定の水位に戻ると、フロートが上昇し、バルブを閉じます。このように、時計の貯水槽は、水位に正味の増減がないように調節することができ、それによって、正確な時間の推定を得ることができる一定の水の流出速度を維持することができる。このタイプの水時計の精度は、17世紀に振り子が時計機構を調整するようになるまで失われることはなかった。
古代や中世の時代にも多くの自己制御装置が発明されたが、「修正フィードバック」を取り入れた制御装置の研究が体系的に行われるようになったのは、18世紀後半、蒸気機関の発明以降である。初期の蒸気機関は、ボイラーから発生する蒸気圧とエンジンにかかる仕事量の両方によって速度が左右されるという大きな制約があった。ジェームズ・ワット(1736-1819)は、エンジンの速度を制御する遠心フィードバック弁を開発し、蒸気機関の効率と安全性を大幅に向上させた(Rosen, 2010)。この「ガバナ」(図4)は、回転する垂直軸の両側で回転する一対の金属球を使用し、エンジンの回転数が上がると、金属球(フライウェイトと呼ぶ)の回転速度も上がり、遠心力の増加により、金属球がばらばらになるように配置されていた。すると、弁が開いてエンジンに入る蒸気の量が減り、ゆっくりした速度に戻る。逆に、エンジンの回転数が下がると、フライウェイトの回転も下がり、外側の遠心力が弱まる。すると、フライウェイトがより近くに落ちて(重力で下がって)、蒸気弁が閉まり、より多くの蒸気がエンジンに入り、速度が上がる。水時計や貯水池の水位と同じように、蒸気圧の変動や作業負荷の変化にもかかわらず、人間が常に監視することなくエンジンの回転数を一定に保つことができたのである。

図4 ジェームズ・ワットの蒸気機関車フライウェイトガバナーを模式的に表現したもの
詳細は本文を参照。出典:パブリックドメイン、から修正したもの、https:www.mpoweruk.com/figs/watt_flyball_governor.htm。
19世紀後半、James Clerk Maxwell(1831-1879)は、ワットのガバナーを数学的に解析し、自己制御装置を理解するための原理を確立し、制御理論の基礎となった(Maxwell, 1868)。1927年、ハロルド・S・ブラック(1898-1983)は、大西洋横断電話の信号を増幅する電気回路にフィードバック制御を適用した(Black, 1934)。彼の負帰還増幅器(1937年特許取得)は、エレクトロニクス分野における最も重要な開発の1つであると考えることができる。システム制御理論のさらなる進歩は、第二次世界大戦中、対空兵器のサーボ制御(負帰還)機構の開発で達成された。
1943年、マクスウェルが最初に説明した制御理論の数学的原理が、生物における行動の説明に適用できることを立証する2つの有力な論文が発表された。アルトゥーロ・ローゼンブルース、ノーバート・ウィーナー、ジュリアン・ビゲローの「行動、目的、テレオロジー」(Rosenblueth et al., 1943)とウォレン・マッカロクとウォルター・ピッツの「神経活動に内在するアイデアの論理計算」(McCulloch and Pitts、1943)は、生物における生理現象の自己制御性と技術者が設計したネガティブフィードバックシステムを結びつける最初の確立となった。興味深いことに、ローゼンブルースはキャノンと密接に仕事をしており、間違いなく彼のアイデアに影響を受けていた。数年後、ウィーナー(1894-1964)は、動物の自己制御的な制御とコミュニケーションの研究を説明するために、サイバネティクス(cybernetes、ギリシャ語でガバナ(操舵手や操縦者の意)を意味するτηςから)という言葉を導入した(ウィーナー、1961)。この概念は、その後、例えばアーサー・ガイトン(1919-2003)と彼の多くの弟子たちが心血管制御に関して行ったように、生理学的システムのモデリングに広く応用されてきた。このように、生体におけるフィードバック制御の概念は、機械系における制御理論の数学的概念と共進化してきたと言えるだろう。負のフィードバック制御は、以下に述べるように、恒常性を達成するための特に重要なメカニズムである。
前項で述べた水時計や遠心式スチームガバナは、負帰還システムの典型例だ。水時計で見たように、フロート/バルブの開閉によって、水位に関する情報がシステムにフィードバックされ、水位をあらかじめ決められた一定の値に維持するように変化するサイクルが生まれる。このように、フロートは同時に水位に影響を与え、水位から影響を受けるという循環的な因果関係を形成している。重要なことは、このシステムが自動的な自己調節システムであることである。つまり、変数が調節される操作レベルが設定された後は、外部からの調節は必要ない。
閉ループフィードバックシステムの一般的な形態を簡略化して図 5に示す。(1)制御される変数(または変数のセット)、(2)対象の変数をモニターするセンサー、(3)コンパレーターまたは中央処理装置(数学的には伝達関数、入力/出力関係)、センサー(求心または感覚経路)から提供された情報はシステムにフィードバックされる。この情報は、「望ましい」状態(設定点または動作点)と比較され、エラー(望ましい状態と実際の状態との差)が検出されるほか、(4) エフェクター(求心性経路または運動経路)が作動してエラーを修正する。エフェクターの活動は、変数の変化に対抗し、それによって緩衝する。この図では、実線は構成要素間の直接的な関係(増加は増加に、減少は減少につながる)を示し、破線は逆の関係(増加は減少に、逆はその逆)を示している。負帰還制御では、変数を所望の値の狭い範囲に維持するために、奇数本の破線を含む必要がある。

図5 負のフィードバック制御を模式的に表したもの
実線は、接続された成分が直接的に関連していることを示す(ある成分の増加は接続された成分の増加につながり、減少は接続された成分の減少につながる)。破線は、連結成分が反比例の関係にあることを示す(ある成分が増加すると連結成分が減少し、減少すると連結成分が増加する)。破線の数が奇数であることは、ネガティブフィードバックによる因果関係のサイクルに必要な条件である。負帰還は、制御される変数を狭い範囲の値で維持するように作用する(詳しい説明は本文を参照)。
負帰還の一般的な例として、図6に示すようなサーモスタット制御の冷暖房システムによる室温の調節がある。室温は調節される変数であり、センサーは温度計、コンパレーターはサーモスタット(所望の温度(動作点)と実際の温度(エラー検出)を比較する装置)、エフェクターは暖房または冷房システムである。この例では、外気温の上昇をセンサーが検知し、その情報がサーモスタットに伝わる。温度情報は動作点と比較され、実際の温度と所望の温度に十分な差がある場合、冷却システムが作動し、加熱システムが不活性化される(エラー信号が減少する)。逆に環境温度が下がれば、冷房は停止し、暖房は作動する。このように、さまざまな外的条件の変動にもかかわらず、安定した室温を維持することができる。

図6 負のフィードバック制御の概念を説明するために、室温の調節を模式的に表したもの
実線は、接続された成分が直接的に関連していることを示す(1つの成分の増加は接続された成分の増加につながり、減少は接続された成分の減少につながる)。破線は、連結成分が逆相関していることを示す(ある成分が増加すると連結成分が減少し、減少すると連結成分が増加する)。負のフィードバックは、周囲の温度が変化しても室温を狭い範囲の値に維持するように作用する(詳しい説明は本文を見てほしい)。
生物系(生体)におけるフィードバック制御は、前項で述べた機械系の単純な「時計仕掛け」のフィードバックシステムよりもはるかに複雑であることを強調しておかなければならない。この注意点をしっかりと念頭に置きながら、生体システムにおける自己調節の概念を血圧の調節で説明することができる。19世紀半ばには、動脈血圧は中立的な反射調節の活性化によって狭い範囲に維持されていることが明らかになった(Adolph, 1961)。しかし、動脈血圧の恒常性調節を説明するために負のフィードバックの原理が適用されたのは、1960年代になってからだ。血圧調節の複雑さを詳細に説明することは、本小論の範囲を超えている(最近のレビューについては、Dampney, 2016を参照)。それにもかかわらず、室温に用いたものと類似した簡略化したフィードバックサイクルを図7に示す。

図7 負のフィードバック制御の生理的な例として、動脈血圧の調節を簡略化した模式図
実線は、連結した成分が直接関係していることを示す(ある成分の増加は連結した成分の増加をもたらし、減少は連結した成分の減少をもたらすであろう)。破線は、連結成分が逆相関していることを示す(ある成分が増加すると連結成分が減少し、減少すると連結成分が増加する)。負のフィードバック調節は、動脈血圧を狭い範囲の値に維持するように作用する(詳細な説明は本文を参照)。NTS = nucleus tractus solitarius、感覚情報が処理され、求心性反応が開始される部位。室温調節における「サーモスタット」に類似した「バロスタット」として機能する。SV = ストロークボリューム(心室収縮ごとに心臓から送り出される血液量)、HR = 心拍数、1分間の拍動(心室収縮)数、TPR = 全末梢抵抗、血液の前進に対する抵抗(血管直径に反比例する)。
この図について説明する前に、まずオームの法則の式を用いて動脈圧を数学的に定義しなければならない(電気回路ではなく水力回路の場合)。代数的には、血圧(BP-電気回路の電圧Eに相当)は、心拍出量(CO-電気回路の電流Iに相当)と全身血管抵抗(TPR-電気抵抗Rに相当)の積である。心拍出量は、1回の拍動で送り出される血液量[ストロークボリューム(SV)]に1分間の拍動数[心拍数(HR)]を掛けたものである。
つまり、BP=SV×HR×TPRとなる(電気回路ではE=I×R)。
動脈圧の変化は、心臓からの出力(SVおよび/またはHR)または血管を通る血液の移動に対する抵抗(血管径を調整することによって、直径はTPRに反比例する)、またはその両方を修正することによって対抗できることが明らかだ。図7に戻ると、センサーは動脈血管(大動脈弓と頸動脈洞)にある受容体(圧受容器)で、動脈圧の変化に反応する(血圧が上昇すると受容体の活性が高まる)。コンパレータ機能は、脳の髄質[孤束核(NTS)]内の神経細胞群によって行われ、そこで信号が処理されてエフェクターシステムの出力に影響を与える。これは、図6に示す室温調節のサーモスタットに似た機能で、「バロスタット」として機能する。信号はNTSで処理された後、髄質内の興奮性領域(ニューロン間結合を介して腹側側髄質(RVLM)と抑制性領域(単シナプス的に両性核(NA)に作用して運動反応を引き起こす(詳細は図8参照)。中枢神経系から標的臓器への運動出力は、心臓への2組の神経によって行われる。副交感神経(NAに由来)はHRを減少させ、交感神経(脊髄の中側柱、IMLに由来し、RVLMからのニューロンによって調節される)はHRおよびSVを増加させる。交感神経は血管にも行き、その活性化によって血管径が小さくなり、それによってTPRが上昇する。したがって、万一、血圧が上昇した場合には、いわゆる圧受容器反射が活性化される。副交感神経の活動が高まり、交感神経の活動が低下すると、心拍出量が減少し(HRとSVが減少)、TPRが低下する。血圧が低下すれば、逆の変化が起こる。このように、負のフィードバック制御は一過性の変化を緩衝し、環境や行動条件の変化にもかかわらず、一日中、一拍ごとに安定した血圧を維持するのに役立つ。

図8 動脈圧受容体反射による血圧調節に関与する中枢神経構造を簡略化して示す。
頸動脈洞と大動脈弓にある動脈圧受容体(動脈圧が上昇すると神経発火が増加する)は、それぞれ舌咽神経(IX番目)と迷走神経(X番目)を介して求心性の情報を脳に伝える。この情報は、まず至陰核(NTS)にあるニューロンで処理される。そして、NTSは、副交感神経と交感神経の放出活動を変化させる。具体的には、NTSは、曖昧核(NA、副交感神経前部神経節)にあるニューロン(単シナプス)と尾側腹外側髄質(CVLM)にあるニューロン(多シナプス、ニューロン間結合を介して)の活性を変化させる。CVLMは、次に、吻側腹外側髄質[RVLM、脊髄の中間側柱(IML)に位置する交感神経前ガングリオンニューロンを制御する]に由来する緊張性交感神経活性を制御する。+ は興奮性神経伝達物質(黒色で表示)、-は抑制性神経伝達物質(青色で表示)、SANは洞房結節である。例えば、動脈血圧が上昇すると、圧受容器神経の発火が増加し、NTSニューロンの活動が高まり、介在ニューロンを介して、NAにある副交感神経前駆ニューロンの活動の増加とIMLにある交感神経前駆ニューロンの発火減少を引き起こす(RVLMの緊張活動の抑制を介したCVLMによる直接的な影響は少ない)。その結果、心拍数(心臓副交感神経活動、心臓交感神経活動)、ストローク量(心臓交感神経活動)、細動脈血管収縮(末梢総抵抗、心臓交感神経活動)が低下することになる。動脈血圧が低下すると、逆の方向に変化する。心臓での符号変化(SANに対する副交感神経の作用)と髄質内での符号変化(CVLMを介したRVLMの抑制)に注意してほしい。この「符号の変化」は、負のフィードバック調節に必要である。
フィードフォワード制御は、環境刺激に対する行動応答の一部として、恒常性を修正し維持するもう一つのメカニズムである。フィードフォワード制御は、しばしばセントラルコマンドとも呼ばれ、制御される変数の状態に関するフィードバックなしに反応が引き出される。つまり、制御される変数に実際に変化が生じる前に、妨害が評価され調整がなされる。例えば、室温を一定にする場合、冬の寒い日に窓やドアを開けると、サーモスタットが周囲温度の変化を検知する前に、フィードフォワード制御によって炉が作動することになる。フィードフォワード調節は、調節される変数の変化とは無関係に働くが、潜在的な妨害の性質と範囲に関する情報を必要とすることを強調しておく。室温の場合、窓やドアの状態(開いているかどうか)をモニターする必要がある(これらの開口部にセンサーを設置する)。そうでなければ、室温が設定温度から十分に逸脱してサーモスタットに検知されるまで(つまり、先に述べた負のフィードバック応答が作動するまで)、応答が引き出されることはない。生物では、学習と経験によってフィードフォワード制御に必要な情報が得られる。猫は、ネズミ(食べ物)と近所の犬(危険で吠える厄介者)の違いをすぐに学習し、それに応じて反応する(食欲や嫌悪の刺激に対して適切な行動や生理的調整を行う)。
前項で述べた単純な負のフィードバックスキーマでは、生物が機能し、変化する環境条件に適応することを可能にする恒常性維持プロセスの複雑さを十分に伝えることはできない(Carpenter 2004)。例えば、負のフィードバック制御の動作点(より正確には動作範囲)は、より高いレベルの制御によって調整され、あるいは上書きされることもある(Goodman, 1980)。このような自動的な(例えば、フィードバック)調節の調整により、生物は適応し、変化する外的条件に適切に対応することができる。この階層的な制御は、図9(Goodman, 1980より改変)に示すように、多階層・多目標追求システムである。
この模式図では、第1レベルは生理化学的プロセス、器官や組織の機能、ホメオスタシスが作用する構成要素を表している。第二のレベルは、自律的(自己)調節、恒常性(例:圧受容体反射)である。ここでは、ある変数の変化が感知され、より高いレベルの制御からの入力なしに、第1レベルのプロセスの調整が開始される。
第三のレベルは、第二のレベルから送信された情報を処理し、他の感覚入力からの情報と統合して、変化する環境条件に対する生理的および行動的応答を調整する中央のコマンドおよび制御センター(中枢神経系)に見出される。上位の中枢は「介入」することができ、自律神経(=自律的・自動的)プロセスをサポートするために必要な調整を行う。この制御は、意識的または無意識的なレベルで行われる。意識的な介入の例としては、室温の変化に対応するための行動(衣服の着脱、日陰や日向を求める窓の開閉など)を開始することが挙げられる。- 一方、無意識的な制御の例としては、運動中の血圧調節の調整(安静時と比較して血圧が上昇してもHRとSVがともに上昇するような圧受容器反射の作動点の変化;Raven et al., 2006)などが挙げられる。このように、第3のレベルは、快適さを維持し、生存を確保するために、外部環境に対する行動と生理的な反応を調整するものである。しかし、第1レベルの構成要素が適切に機能しなければ、より高いレベルの制御は不可能であることを強調しておかなければならない。最後に、さらに高いレベルの制御として、生体の外側の要因を想定することもできる。

図9 恒常性維持制御の高次制御を簡略化して模式的に表したもの
このような階層的な制御により、制御のレベルがより細かくなり、生物が変化する環境条件に適応できる柔軟性が高まる(詳細は本文参照)。CNS = 中枢神経系。
現代のジェット旅客機のcを使って、制御のレベルを説明することができる(Wiener, 1961)。好ましい方位、姿勢、対気速度が設定されると、風速の変化や小さな乱気流があっても、自動操縦はロール、ピッチ、ヨーの許容範囲内で水平飛行を維持する。しかし、離陸と着陸は(少なくとも「自動運転」技術が完成するまでは)人間のパイロットが直接介入する必要がある。このように、旅客機の構成要素であるジェットエンジンや機体(胴体、翼、フラップ、ラダーなど)を第1階層、自動操縦装置を第2階層、人間のパイロットを第3階層とする。この例では、パイロットに指示を出す航空管制官が第4の管制を行い、さらに上位の管制は航空管制官の方針を決める連邦航空局(FAA)が行うことになる。

運動による心肺反応は、このような恒常性調節の階層的な制御を示す生理学的な例となる。第1階層は、心血管系と呼吸器系を形成する組織と器官(心臓、肺、血管のほか、塩分と水分の保持、ひいては血液量を調節する腎臓と内分泌腺)、第2階層は圧受容器(直接効果)と心皮質反射(血液量の調節による間接効果)、第3階層は中枢神経系の髄質(NTS)で起こり、感覚情報の処理と発散反応の始動を行う。髄質構造は、それ自体、脳の高次中枢(視床下部や運動中枢など)によって制御されている。実際、視床下部は、内部環境の変化と外的課題に対する行動反応の調整(マッチング)に大きな役割を担っている。前述のように、運動中はHRと血圧が同時に上昇するため、圧受容器反射の調節が変化していることがわかる。このような調節は、運動する筋肉の代謝需要の増加に対応できるように、酸素供給量を増やすために必要である。Ravenら(2006)は、これらの調整は、圧受容器反射の抑制ではなく、圧受容器反射をより高い動作点に移動させる(すなわち、恒常性調節の範囲を変更する)ことによって生じることを実証した。フィードバック(運動筋の感覚情報、いわゆる運動圧反射)とフィードフォワード(中央指令:例えば、走る前にレースをイメージするなど、運動開始を予期するとHR、BP、骨格筋血流が増加する)の両方が、こうした反射調整に寄与する。最後に、より高いレベルのコントロールとして、レースの開始時刻を決定するスターター、どのようなレースを行うかを決定するイベント主催者、イベントを管理するルールを設定するスポーツ規制機関(オリンピック委員会、FIFA、NCAAなど)などがある。

したがって、内部環境の恒常的な制御は、単純な負のフィードバック制御をはるかに超えるものである(Carpenter, 2004)。階層的なコマンドとコントロールのレベルにより、生物は、変化し、しばしば敵対的な環境から課される課題に対応し、適応し、その内部条件を調整することができる。適応は、実際には、ホメオスタシスの創発的な性質とみなすことができ、生命のユニークな性質の原因となっている可能性がある(Turner, 2017)。
ホメオスタシス:還元主義への示唆
…All kings’ horses and all kings’ men
ハンプティ・ダンプティを元に戻せなかった
えいじゅのはやし
ホメオスタシスの概念は、無傷の生物の生理学を理解する上で重要な意味を持つ。近年、生理学のみならず科学全般において、全体論的なアプローチではなく、還元論的なアプローチ(複雑な現象をより小さく、より単純な構成要素に還元して説明しようとする考え方)が主流になってきている。
タレスやソクラテス以前のギリシャの哲学者たちは、すべての物質は地、空気、火、水という4つの重要な要素のさまざまな組み合わせで構成されていると考えていた(身体の4つの体液はこれらの要素に対応している)(ホール、1975)。ギリシャの還元主義の頂点は、レウシッポスとその弟子デモクリトスの研究にあり、彼らは、万物は、原子と呼ばれる無限に多数の不可分な小さな粒子から構成されていると提案した(Hall, 1975)。科学における還元主義の近代的な適用は、フランシス・ベーコン(1561-1620)とルネ・デカルト(1596-1650)にまでさかのぼることができる。
ベーコンは、自然哲学の知識を得るための新しい経験的方法(Novum Organum 1620、アリストテレスのオルガノンとは対照的に、論理とシロギズム、すなわち演繹的推論に関する論考)(Bacon, 1620)に、帰納推論とともに、後に科学方法として知られるようになった中心要素として還元論を取り入れた。
デカルトも同様に、ベーコンが提唱した帰納法(経験主義)ではなく、演繹法(合理主義)を重視しながらも、知への道筋として還元主義を採用した。デカルト(1637)は、「理性を正しく働かせ、科学において真理を探究する方法に関する講話」の中で、生物学的研究に大きな影響を与えることになる2つの概念を紹介している。デカルトは、この最も影響力のある論文で、知識を得るための4つの教訓を述べている。特に第2と第3の教訓は、還元主義者のアプローチを次のように例証している:
「第二は、検討中の各難題を、可能な限り多くの部分に分割し、その適切な解決に必要であろうとすることである」
「第三は、最も単純で容易に理解できる対象から始めて、少しずつ、いわば一歩一歩、より複雑なものの知識へと昇るような順序で、私の思考を行うことである」(デカルト、1637)
彼の第二の結論は、「身体は単なる機械に過ぎない」というもので、より遠大な結論となった。つまり、デカルトの還元主義を適用すれば、個々の器官やその構成部分(分子レベルから細胞内小器官、細胞、組織、器官、そして最終的には無傷の器官に至るまで)の、おそらくより単純な機能を理解することによって、無傷の器官の複雑な生理学を推論できるとしたのである。
このアプローチの威力を否定することはできない。DNAが遺伝の分子であることが判明してからわずか数十年で、約30億の塩基対の配列がヒトや他の種についてマッピングされ、タンパク質合成の遺伝的「コード」が解読され、多数のタンパク質を制御する2万から2万5000のヒト遺伝子が決定された。ハンプティ・ダンプティは、文字通り10億個の破片に砕け散ったのである。

しかし、還元主義には、「部分には全体がある」「複雑さは不完全な理解の産物に過ぎない」という前提がある。言い換えれば、十分な情報(ビッグデータ)を集め、十分な計算能力(超高速コンピュータ)を開発すれば、ハンプティを元に戻すことができるという仮定である。では、この仮説は正しいのだろうか。私たちは、多くの生物種のゲノムを解読しているが、ゲノムが生物になる過程については、ほとんど理解していない。また、神経細胞の活動電位やシナプス伝達の仕組みは詳細に分かっているが、電気的、化学的な事象がどのように意識を生み出すのかは分かっていない。複雑さは、かつてナイーブに考えられていたような錯覚ではないかもしれない。150年以上前、クロード・ベルナールによってエレガントに表現されたように:
生理学者、医師は、生き物がそれ自身の個性を持った生物であることを決して忘れてはならない。物理学者や化学者は宇宙の外側に立つことができないので、身体や現象をそれ自体として、また個別に研究し、必ずしも自然全体と関連づける必要はない。しかし、生理学者は、動物生物を全体としてとらえ、その内部に入り込みながら、全体の調和を考慮し、その各部の機構を理解しなければならない。その結果、物理学者や化学者は、観察した事実の最終的な原因についての考えをすべて否定することができる。
一方、生理学者は、組織化された身体における調和のとれた、あらかじめ確立された統一性を認める傾向があり、そのすべての部分の作用は相互に依存し、相互に生成し合うものである。私たちは、生体を分解して異なる部分を分離する場合、それは実験的な分析を容易にするためであって、決してそれらを別々に考えるためではないことを、本当に学ばなければならない。
実際、ある生理学的性質にその価値と真の意味を与えようとするとき、私たちは常にこの全体を参照し、全体におけるその影響についてのみ結論を出さなければならない。(中略)バーナード、1865)
全体は部分の総和よりも大きいというのは、言い過ぎではないだろう!
2010年に発表されたポストゲノム時代(Billman, 2010)において、現代の生理学が直面する大きな課題は、試験管内で得られた膨大な情報をいかに統合し、生体内の機能に関する一貫した理解に結びつけるかである。機械は多くの部品から構成されているが、部品が単独で機械になるわけではない。クリスマスイブに子供用自転車を組み立てようとしたことのある人なら、部品があれば機械ができるわけではないことを証言してくれるだろう。

同じように、生物は分子からできているが、分子は生物ではない。1つの遺伝子、1つのタンパク質、1つの機能という概念は、生物の目もくらむような複雑さと驚くべき美しさ、つまり「自由で自立した生命」に必要な恒常性維持機構の複雑なダンスを説明するには、極めて不十分なものでしかない。DNA分子の塩基対の配列が生命の複雑さを説明することは、コンパクトディスクの録音にある一連の1と0が音楽に対する感情的な反応を説明すること以上にできない(Noble, 2006)。人間や他の生物は、利己的であろうとなかろうと、遺伝子を永続させるための単なる乗り物ではない。ハンプティ・ダンプティを元に戻すには、よりホリスティックで統合的なシステムアプローチが必要である。今こそ生理学者は原点に立ち返り、クロード・ベルナールが提唱したように、生物を全体として考える時なのだ。
第二の課題は、臨床応用の前提となる無傷の動物や臓器を用いた統合的研究を行うことができる次世代の研究者を育成することであり、決して重要ではない。残念ながら、統合生理学のトレーニングプログラムの数は減少の一途をたどっており、その結果、全動物生体内試験実験に必要なスキルを持つ人材が不足している。この問題は、医学部内の生理学教室が改名されたり、実際に廃止されたりして、さらに悪化している。現在、生理学教室は”Department of Molecular Biology/Physiology”(分子生物学・生理学教室)と改名するのが流行っている。原子生理学教室はどうだろうか。
分子的・遺伝的アプローチが重視されるようになり、生理学の入門コースすら受講していない生理学教室のメンバーが珍しくなくなった。生理学という知的学問の面白さは、学生の研究室でこそ味わえるものなのに、これは実に残念なことである。このような古典的な生理学の実験を行うことほど、実践的な学習やホメオスタシスの概念に対する理解を深めることができるものはないだろう。学生実験室では、教科書に載っているような生理原理を説明するだけでなく、実際に動いているところを見ることができる。カエルの骨格筋と神経の機能を初めて調べたり、ネクトゥルス(マッドパピー)の糸球体で色素がクリアになるのを見たり、哺乳類(ラット、ウサギ、イヌ)の心肺調節の謎に思いを馳せたとき、初期の研究者が抱いたであろう興奮と驚きを、学生は直接体験できる。このように、「分子から無傷の生物まで機能を統合する」という壮大な課題に、十分な数の適切な訓練を受けた研究者が対応できるかどうかは、まだ未解決の問題である。
まとめ
生理的調節に関する私たちの理解は、体液のバランスに関するギリシャの考えから、クロード・ベルナールの”milieu intérieur”、ウォルター・キャノンのホメオスタシスの概念の定式化、制御理論(フィードバック制御)の応用へと、時代とともに進化してきた。ホメオスタシスは、生理学の中心的な統一概念となり、生体が変化する外部条件に適応しながら内部安定性を維持できる自己調整プロセスと定義されている。
ホメオスタシスは、静的で変化しないものではなく、外的な課題を克服するために必要に応じて内部条件を変化させることができる動的なプロセスである。このことは、キャノンがホメオスタシスという言葉を作る際に気を配ったことからも明らかだ。
彼は、ギリシャ語で「同じであること」ではなく「似ていること」を意味する言葉を意図的に選び、内部条件が変化しても安定(単一の値ではなく、値の範囲内)することを強調した。つまり、ホメオスタシスは「停滞」を意味しない。また、ホメオスタシスの調節は、単に単一の負のフィードバックサイクルの産物ではなく、より高い制御センターによって変更可能な複数のフィードバックシステムの複雑な相互作用を反映していることも重要である。このような階層的な制御とフィードバックの冗長性により、より細かいレベルの制御と、変化する環境条件への適応を可能にする高い柔軟性が生み出されている。
生物の健康と活力は、内部環境の恒常的な制御の結果であると言え、この概念を理解することなしには、正常な生理学を理解することはできない。逆に言えば、ホメオスタシス機構が破壊されることが病気につながるということであり、効果的な治療は、自然に逆らわず、むしろ自然とともに働き、ホメオスタシス状態を再確立することに向けられる必要がある。
利益相反行為について
本研究は、潜在的な利益相反と解釈されうる商業的または金銭的関係がない状態で実施されたことを宣言するものである。
謝辞
前述の通り、本エッセイで紹介した内容の一部は、「Handbook of Systems and Complexity in Health」(Billman, 2013)の第10章として発表されたものであり、出版社の許可を得て(修正・更新を加えて)再掲載している。
