Contents
Chrononutrition and Polyphenols: Roles and Diseases
www.ncbi.nlm.nih.gov/pmc/articles/PMC6893786/
要旨
生体リズムは生理活性化合物の活性に影響を与えると同時に、これらの化合物の摂取により生体リズムを変調させることができる。このような背景から、生体リズムと栄養・代謝の相互作用を研究することを中心とした研究分野として時間栄養学が登場している。
本総説では、フェノール化合物が生体リズムの変調に果たす役割について、慢性疾患の治療や予防への効果を中心にまとめた。
従属栄養生物は、フェノール化合物などのファイトケミカルを媒介とした化学的な合図を感知し、環境条件への適応を促進する。これを異性ホルミシスと呼ぶ。したがって、フェノール化合物を豊富に含む果物や野菜の消費は、主にそれらの代謝産物に起因するいくつかの健康上の利点を発揮する。
しかし、植物に含まれるフェノール化合物のプロファイルは種によって異なり、農業や技術的要因によって大きく変動する。この意味で、ポリフェノールが豊富な果物の季節的な消費は、果物が含む特定のフェノールプロファイルのために、生理や代謝の調節に重要な変化を誘発する可能性がある。
このことは、より正確な食事推奨を確立するために、これらの特定のフェノールプロファイルが健康に与える影響を評価する研究の必要性を浮き彫りにしている。
キーワード
慢性栄養学、生体リズム、ポリフェノール、健康効果、疾患、メタボリックシンドローム、栄養学、クロノニュートリション
1. はじめに
1.1. 生体リズム(概日リズムと季節リズム
すべての生物に存在し、概日リズムと周年リズムを含む生物リズムは、代謝や栄養と密接に関連している。この枠組みの中で、光は、生物の行動を駆動する生理学的・代謝的シグナル伝達経路の関連する変化に極めて重要な役割を果たしている[1,2]。さらに、光は、時期や生物に応じて、生殖、移動、冬眠、発芽、開花などの季節特有の行動を調節する[3,4,5]。内生的なリズムの存在は、このようなタイプの調節過程を示す最も初期で最も原始的な生物の一つである藍藻から始まった進化において、明らかに優位性を持っている[6]。しかしながら、これらの反応が光誘起遺伝子の発現によって高度に制御されていることは知られているものの、そのメカニズムはいまだに明確には解明されていない。その意味で、体内時計に影響を与える因子、例えばコアクロック機構の遺伝子多型や明暗周期の季節的変化が生理活性に顕著な影響を与えることが明らかになってきている。
近年では、生体リズムが生理活性化合物や食事から摂取した栄養素の生理活性に影響を与えることが観察されている。さらに、栄養素の摂取によっても生体リズムが変調されることがある。この概念に基づき、朝食などの長期断食後の最初の食事が体内時計の同期に重要であることを報告している著者もいる[7]。さらに、食事からのポリフェノールなどの生理活性化合物も時計と相互作用し、活性化されたシグナル伝達経路に従って、それらの消費の正確なタイムポイントを確立することができることを強調する価値がある[8]。最近では、概日リズム、マイクロバイオーム、肥満との関連が報告されており、その中で腸内マイクロバイオームは概日リズムと関連して肥満に影響を与える可能性のある内分泌系として理解されている[9]。また、機能性食品の摂取は、少なくとも一部ではサーカディアンリズムの変調を介して、肥満やそれに伴うメタボリックシンドロームなどの多くの慢性的な生理的疾患を予防または改善することで有益な効果があることが示されている[10]。これらの知見はすべて、私たちの体の生理学的および代謝応答における概日リズムの重要性を示しており、メタボリックシンドロームなどの病態を予防または改善するために、この知見をどのように利用できるかを示している。
1.2. 分子メカニズム
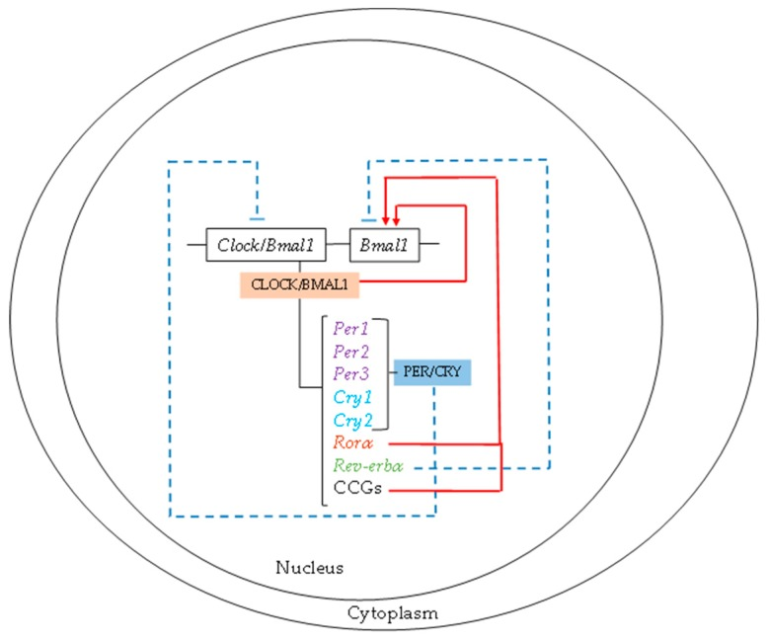
概日リズムは、すべてのプロセスが一日を通して最大値と最小値の機能を発揮することを保証している。現在、生物学的リズムを駆動する分子機構は、2つの連動する転写翻訳フィードバックループ、概日運動器出力サイクルカプト(CLOCK)と脳と筋肉のARNT様タンパク質1(BMAL1)のセットで構成されていることが認められている。CLOCK/BMAL1ヘテロダイマーは、すべての細胞に共通の中心時計であり、3つの周期(Per)と2つのクリプトクローム(Cry)遺伝子の転写活性を刺激する[9]。PER/CRYヘテロダイマーは、CLOCK/Bmal1転写産物発現の負のフィードバックループとして作用する[11]。さらに、CLOCK/BMAL1ヘテロダイマーはBmal1遺伝子の発現を活性化する。さらに、CLOCK/BMAL1によって発現が制御されるRorαとREV-erbαという2つのフィードバックループが存在する。RORαはBmal1の活性化因子として、REV-ERBαは抑制因子として作用する(図1)。
図1 生体リズムの分子機構
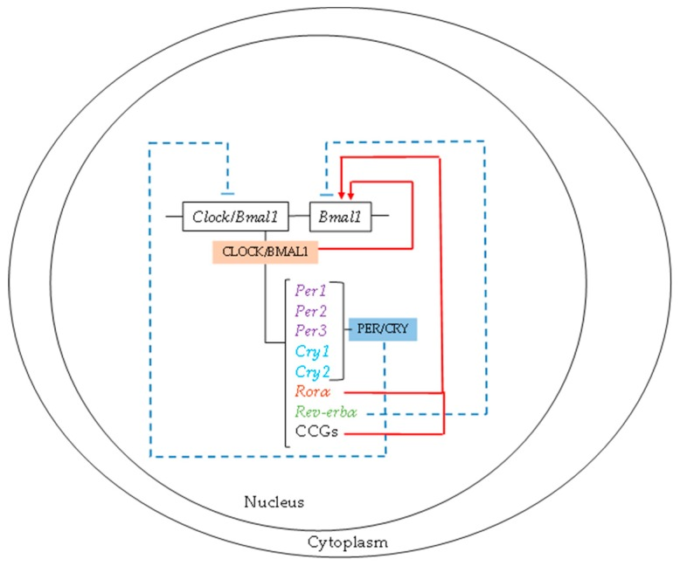
概日運動器出力サイクルkaput/脳と筋肉ARNT様タンパク質1(CLOCK/BMAL1)ヘテロダイマーは、すべての細胞の中心的な時計であり、そのヘテロダイマーがClock/BMAL1転写発現の負のフィードバックループとして作用するピリオド(Per)とクリプトクローム(Cry)遺伝子の転写活性を刺激する。RorαおよびRev-erbα発現の2つのフィードバックループは、CLOCK/Bmal1によって制御されている。
一方、CLOCK/BMAL1ループは、クロック制御遺伝子(CCG)の転写を誘導する。このループの活性は、これらのCCGの概日リズム発現を決定し、したがって、代謝や機能的な細胞リズムを決定する。さらに、サーチュイン1(SIRT1)ニコチンアミドホスホリボシルトランスフェラーゼ(Nampt)ニコチンアミドアデニンジヌクレオチド(NAD+)が関与するエピジェネティックループがある。CLOCK/BMAL1が過剰発現するCCG遺伝子であるNamptは、NAD+の細胞内濃度を上昇させ、それに伴ってSIRT1を活性化させ、BMAL1とヒストンの脱アセチル化を誘導し、CLOCK/BMAL1の転写活性を抑制する。NADの他に、時計制御機能を持つ主要な細胞内生化学成分として、一リン酸活性化プロテインキナーゼ(AMPK)と環状アデノシン一リン酸(cAMP)がある。細胞型に応じて特異的なCCGが存在する[12]。このように、分子時計は、代謝経路や転写因子、核内受容体を介してCCGを介して細胞の機能や代謝を制御しており、酵素やその他の代謝因子の発現を誘導している[13,14]。さらに、光はこれらすべてのプロセスを制御し、体内のさまざまなシグナル伝達経路の活性化を誘発する(図2)。この点で、メラトニンは暗黒サイクルの間、光サイクルの代謝をオフにするという極めて重要な役割を果たしている。
図2 光が生体リズムに及ぼす影響
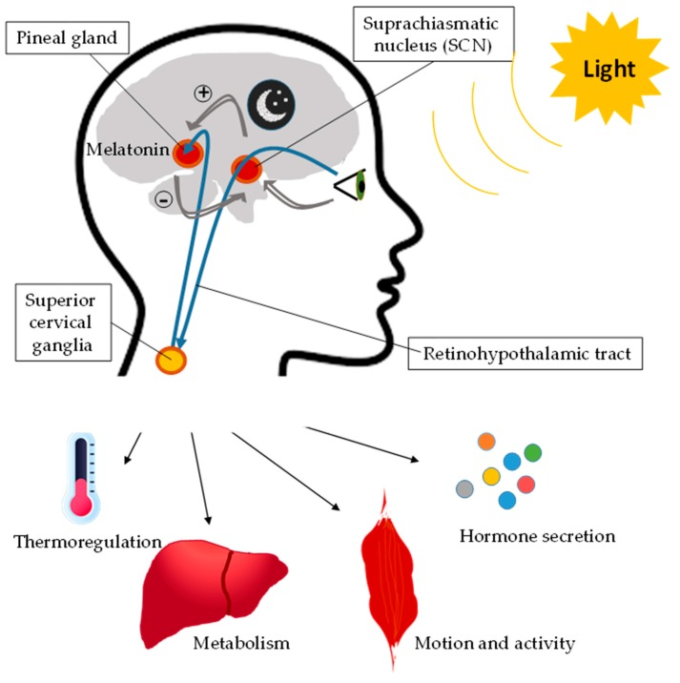
光は、視交叉上核の活性化によって生体リズムを調節し、これが上頸神経節を活性化して、体温調節、代謝、運動、活動、ホルモン分泌を調節する経路など、体内のさまざまなシグナル伝達経路を誘発する。暗黒期には松果体はメラトニンを合成し、このメラトニンが視交叉上核の作用を阻害する。
ClockおよびBmal1の変異体は、肥満、メタボリックシンドローム、および2型糖尿病を特徴とする異常な代謝表現型をもたらすことが十分に確立されている。これらの障害は、膵臓での急性グルコース投与後の低血糖とグルコースクリアランスの上昇[15]、膵臓での高血糖と耐糖能の低下[16]、骨格筋でのインスリン依存性グルコース代謝の低下[17]、白色脂肪組織特異的な時計の障害で視床下部の食欲中枢を調節する脂肪細胞からの多価不飽和脂肪酸の異常分泌[18]によって起こる。近年、我々のグループは、グルコースおよび脂質代謝の季節変動を分析することにより、代謝恒常性の周年リズムを研究してきた[19]だけでなく、高血圧やインスリン抵抗性などの肥満に関連するいくつかの併存疾患も分析してきた。我々の研究は、概日リズム関連遺伝子Cry1,Bmal1,Per2,およびNr1d1に関連するシグナル伝達経路を調節することによって、生物学的リズムがこれらの病理学的パラメータをどのように調節することができるかに焦点を当てていた[20]。本レビューでは、メタボリックシンドロームなどの慢性疾患の治療または予防に潜在的な焦点を当てて、生体リズムの変調におけるポリフェノールの役割をまとめている。
2. 時間栄養学
2.1. 定義
ここ数年、体内時計が栄養に与える影響についての認識が高まってきており、エネルギーバランスや代謝へのさまざまな影響、健康や病気への影響がある。この概念は 2005 年に出版された栄養と健康に関する日本語の本 [22] で初めて言及された時間栄養学[21] として知られる新しい学問分野の発展につながった。それ以来、時間栄養学は、生体リズム、栄養、代謝の相互作用の研究に焦点を当てた研究分野として浮上してきた[21]。具体的には、食物摂取のタイミングや生体リズムが健康、代謝、栄養にどのような影響を与えるかだけでなく、栄養(食事の構成や大きさ)が体内時計システムにどのような影響を与えるかについても研究されている[23]。
この分野で行われた臨床研究の大部分は、主に食事のタイミングの影響に焦点を当てている。この点では、朝食を抜く、夕方に高エネルギーの食事を摂る、食事や間食の頻度を増やすといった食事のタイミングパターンが、太りすぎや肥満のリスクの増加、およびヒトにおける代謝への悪影響と関連していることが観察されている[24]。実際、概日食パターンの変化は、ヒトとげっ歯類の両方でエネルギー摂取量の増加をもたらす [25,26,27,28,29,30]。さらに、過体重女性と肥満女性では、食事のタイミングの変化後に体重減少に対する差動効果が示されている[31]。一方、短い睡眠時間は、高血圧、2型糖尿病、肥満などのさまざまな慢性疾患の発症と関連している[32,33,34]。これらの影響の根底にあるメカニズムはまだ完全には解明されていない。しかし、近年、様々な研究により新たな知見が得られている。概日リズムの変化は、グルコースおよびインスリンレベルの上昇、動脈圧の上昇、レプチンレベルの低下、睡眠不足、コルチゾール分泌リズムの変化につながる[35]。満腹感を調節するレプチンシグナル伝達は概日変動を示し、時計遺伝子の変動と糖尿病や肥満などの代謝性疾患との関連が考えられる[35]。実際、異常な食物摂取タイミングは、満腹感シグナルの変化および血清レプチンレベルの低下と関連している[25,36,37]。レプチンレベルのこの減少は、エネルギー支出を減少させながら食欲の増加をもたらし、したがって、肥満および他の代謝性疾患の発症を促進する[35]。代謝調節に対する概日時計の影響を説明するもう一つの要因は、熱発生およびエネルギー消費に対する影響である[25]。このように、熱発生は食事の摂取によって誘導され、概日リズムに沿っており、午前中に最も高く、午後から夜にかけて続く[25]。したがって、朝食を抜くことが体重増加と関連している理由は、このことが説明できるかもしれない。最近では、午前中に増加した食事誘発性の熱発生は、午前中に増加するノルエピネフリンとエピネフリンの循環の概日リズムに起因する可能性があることが示されている;これらの化合物は食物摂取量の調節に影響を与えることが知られている[38,39]。さらに、健康な人を対象とした研究では、夕方に同じ食事を摂取すると、安静時代謝率(RMR)が低下し、血糖値/インスリン血症反応が増加したことから、エネルギー支出および代謝パターンの概日変動が示唆されている[40]。飢餓ホルモンであるグレリンもまた、サーカディアン振動を示しており、朝に最も低い値を示し、褐色脂肪の熱産生の抑制を介してエネルギー支出と熱産生を調節している[38]。
2.2. マクロ栄養素と生物活性物質
一方、上述のように、食事のパターンは生体リズムに影響を与える可能性がある[23,41]。したがって、Per1/Per2などの時計遺伝子の発現レベルは、日中の摂食によって肝臓で変動を示した[42,43]。さらに、摂食リズムの不在と副腎ホルモンの不在は、肝内時計遺伝子のリズムを枯渇させた[44]。栄養素および食物因子はまた、全身の細胞性概日時計または時計系を調節することができる。したがって、高脂肪食(HFD)の消費は、体重調節の変化に寄与し、そのようなレプチンとインスリン、クロック調節核因子RORαとPPARα、視床下部神経ペプチドAgRPとNPY、および脂質代謝に関与する因子などの循環代謝マーカーの変化した概日パターンに寄与する可能性があるマウスの光の期間中に増加した食品の摂取につながる[45]。また、HFDの消費は、通常の食事[46]に関連付けられている発現と比較して異常なクロック遺伝子発現につながる、エネルギー不足の状況で活性化されているAMPKキナーゼシグナリングを変更することが報告されている。このHFD媒介AMPKシグナル伝達の変化は、肥満、高インスリン血症、肝性ステアトーシス、および炎症[47,48]に対する保護につながる、自由摂取のアクセスと同等のカロリー消費にもかかわらず、時間制限付き給餌とマウスで改善された。また、一定の暗闇の下でマウスのHFDの消費はまた、彼らの運動量の活動を増加させ、概日運動量のリズムを変化させる[45]。ヒトでは、55%炭水化物-30%脂肪から40%炭水化物-45%脂肪に、食事の炭水化物と脂肪の割合の変化は、単球におけるコルチゾール概日振動、炎症性および代謝性遺伝子発現プロファイル、およびPER遺伝子発現リズムの変化につながった[49]。この研究は、食事の種類だけでなく、その構成成分の割合の重要性を強調している。高塩分(HS)食もまた、生体リズムに影響を与えることが示されている。このように、HS食は、腎内髄質におけるBmal1およびCry1の発現ピークの有意な遅延とPer2の発現抑制を伴う腎内概日リズムの変化を示した[50]。
最近、腸内細菌叢は代謝調節の重要な因子として浮上してきており、食物から得られるエネルギーを調節し、宿主および食事由来の産物のレベルを調節するため、概日リズムに対するその潜在的な影響は極めて重要である[51]。このように、食事によって誘導される腸内細菌叢の変化は腸内時計に影響を与え、生物の恒常性に影響を与えることができる。実際、腸内細菌叢の組成はマウスとヒトの両方で概日振動を受けている[51,52]。微生物相はまた、いくつかの神経細胞の機能にも影響を与えているようであり、したがって、腸内細菌叢の相互作用を通じて、体内時計や異なる脳領域が周期的に「栄養」情報を受け取っている可能性がある[51,53,54]。
食品に含まれる個々の栄養素については、いくつかのアミノ酸残基が睡眠/覚醒サイクルの変化に及ぼす潜在的な影響が評価されている。このように、就寝30分前に3gのL-セリンを摂取することと、朝の明るい光への曝露を組み合わせることは、ヒトにおける光誘発性の位相リセットを促進するのに有用であることが示されている[55]。さらに、トリプトファンはセロトニンとメラトニンの前駆体であり、それぞれ睡眠潜時と睡眠の質に関与するホルモンであることから、トリプトファンが睡眠調節に関与していることが知られている[56]。このように、トリプトファンを濃縮した穀類(トリプトファン60mgを朝と夜の1日2回摂取)の摂取は、55~75歳の高齢者において、睡眠効率、実際の睡眠時間、不動時間を増加させることが示されている[56]。さらに、トリプトファン、アデノシン-5′-リン酸、ウリジン-5′-リン酸を豊富に含む穀類を夜間に摂取することで、睡眠障害のある生後8~16ヶ月の乳児の睡眠効率が向上することが示された[57]。さらに、トリプトファン、セロトニン、およびメラトニンを高濃度に含むチェリーまたはチェリージュースを豊富に含む食事もまた、中高年および高齢者の睡眠/覚醒リズムに有益な効果をもたらし、例えば、不眠症を減少させたり、睡眠時間および睡眠効率を増加させたりすることができる[58,59,60,61]。しかしながら、それらの有益な効果は、チェリーの栽培品種の種類に依存する[58]。トリプトファン以外にも、植物本来のメラトニンであるフィトメラトニンやポリフェノールなど、チェリーに含まれる他の化合物がその効果に関与している可能性がある[59,60]。さらに、セロトニンを豊富に含む果物は、睡眠の問題にも役立つ可能性がある。このように、寝る1時間前にキウイ2個を4週間摂取することで、睡眠障害を持つ成人の睡眠の質を改善することが示されている[62]。
2.3. ゼノホルメシス
従属栄養生物は、栄養分に加えて、劣悪な環境下で植物が合成した非栄養分を媒介とする化学的手がかり(ファイトケミカルとも呼ばれる)を感知することができ、これらの生物の適応能力を促進している。このプロセスは、異性ホルメシスとして知られている[63,64]。実際、ポリフェノールなどのこれらのファイトケミカルの一部は、高血圧、癌、糖尿病、肥満、およびその他の医学的状態などの慢性疾患の予防および/または治療に関連している[65]。
3. フェノール化合物. 食事のパターンと疾患
異性ホルミシスに関しては、ファイトケミカルの最も重要なグループの一つはフェノール化合物からなる。これらは、干ばつ、高温または低温、微生物感染、または草食動物による消費など、いくつかのタイプのストレスに応答して植物によって生成される二次代謝物である[66,67,68,69,70]。
植物には、共通して少なくとも1つのフェノール環の存在を有する数千のフェノール化合物が記載されている(8000以上の構造が記載されている)[71]。これらの化合物は、その化学構造に基づいて2つの主要なグループに分けることができる:フラボノイドおよび非フラボノイド。フラボノイドは最も広く分布する化合物である[72]。
図S1は、これら2つのカテゴリー内のフェノール化合物の主なクラスを示している。一般に、これらの化合物は、植物中に高分子形態および/または糖に連結された形態として存在することができる[73]。例えば、プロアントシアニジンとしても知られる高分子量縮合タンニンは、フラバノール単位の繰り返しからなるポリマーである[74]。
フェノール化合物を豊富に含む果物や野菜の消費から得られるいくつかの健康上の利点がある[72,75]。さらに、これらの化合物は、果物や野菜に苦味、渋み、風味、色、酸化安定性を与える。この意味で、ポリフェノールはワインの赤色や苦味の主な原因となっている。
3.1. 野菜に含まれるフェノール類の含有量
果物、ココア製品、およびお茶やワインなどの飲料は、ヒトの食事におけるフェノール化合物の主な供給源である[72,76]。これらの化合物は、根、葉、および果実を含む植物全体に存在する。このように、葉や茎にはこれらの化合物が多く含まれており、主に単量体形態で存在し、高分子ポリフェノールは液胞、葉、表皮、花、果実に存在している。
表S1は、Folin-Ciocalteauアッセイにより決定され、Phenol-Explorerデータベース[77,78,79]から得られた総ポリフェノール含有量を示している。表S1に含まれる果物および野菜の中で、ココア、豆類およびクルミが最も高いポリフェノール含有量を有していた(それぞれ5624.23,1234.38および1574.82 mg/100 g FW)。その他のフェノール化合物を豊富に含んでいるのはザクロ、プラム、イチゴ、オレンジであり、これらのポリフェノール含量は約400~280 mg/100 g FWの範囲である。これらの値は、いくつかの独立した研究で報告された平均値であり、いくつかの要因により異なる可能性があることを考慮に入れなければならない。
植物に含まれるフェノール化合物のプロファイルは種によって異なる。この点、柑橘類にはナリンゲニンやヘスペリジンなどのフラバノン類やフラボン類が豊富に含まれており[80]、さくらんぼ、ぶどう、ベリー類などの色の濃い果物にはシアニジンやマルビジンなどのアントシアニン類が豊富に含まれており[81]、さらにワイン、コーヒー、紅茶などの飲料にはエピガロカテキンガレート、カテキン、エピカテキンなどのフラバノール類が多く含まれている[82]。これらの最後の化合物は通常、液胞液中と表皮[72,83]で発見される。非フラボノイド類の中では、リグナン(ピノレシノール、マタイレシノールなど)が種子やナッツ類に多く含まれている[83]。
3.2. ポリフェノール組成に影響を与える要因
前に説明したように、いくつかの要因は、同じ種内のフェノール化合物の含有量に影響を与え得る。実際、農業的要因と技術的要因の両方が植物のフェノールプロファイルに影響を与えることが示されている。これら2つのグループの中では、品種、環境、土壌施肥、灌漑システム、および成熟段階が主な農業的要因である。一方、技術的要因の中にはポストハーベスト処理がある。
これらの要因の中でも、品種はフェノール化合物のプロファイルを強く左右するため、最も重要な要素の一つである。例えば、文献には7500以上の異なるリンゴ品種が記載されており[84]、Kalinowskaらが記載しているように[85]、そのフェノール化合物の含有量には大きな差がある。例えば、総フェノール含有量は、ガラリンゴの56mgGAE/100gFWからパナイアリンゴの221mgGAE/100gFWまでの範囲であることがある[86]。興味深いことに、これら2つのリンゴは赤色の品種である。FranciniとSebastiani(2013)は、異なるリンゴ間の遺伝的変異がフェノールプロファイルに重要な影響を与えることを示唆している[87]。これらの変化のいくつかは、品種間で異なるフェノール化合物の生合成経路に関与する遺伝子の発現によって説明できる。例えば、ブドウでは、アントシアニンの合成に重要な酵素であるアントシアニジン3-O-グルコシルトランスフェラーゼ2(UFGT)遺伝子の発現不足が、白品種では赤品種に比べて観察されている[88]。
環境要因もまた、果物や野菜のフェノール組成を調節する。例えば、水の利用可能性、温度、光への曝露、土壌の塩分は、フェノール合成の重要な調節因子として記述されている[66,67,68,69,70]。実際、異なる地域で栽培された同じ品種の果実は、異なるフェノール化合物の含有量を示す[89]。例えば、Häkkinen と Törrönen は、フィンランドで栽培された果実(49.3 mg/100 g FW)とポーランドで栽培された果実(36.1 mg/100 g FW)のフェノール含量に有意な差を認めた [89]。これらの結果は、植物のフェノール化合物の含有量における環境要因の重要性を浮き彫りにしている。
最後に、農学的要因については、有機栽培ではフェノール含有量について議論の余地のある結果が観察されている。したがって、いくつかの研究では、この農法が栽培品種中のフェノールの量を増加させることが示されている。例えば、Strackeらは、有機栽培品種の「ゴールデンデリシャス」リンゴは、非有機栽培品種のリンゴよりもフェノール含有量が14%〜19%多いことを示した[90]。しかし、Winterらの研究[91]では、著者らは、有機栽培品種と慣行栽培品種の間には一貫性のない違いが存在すると結論づけている。別の研究では、Mulero ら [92] は、未熟な有機栽培品種と非有機栽培品種の間には有意な差が見られたが、その差はブドウが成熟期に達すると消失することを述べている。したがって、この分野ではさらなる研究が必要である。
技術的な要因としては、洗浄、貯蔵、抗菌処理、最小限の処理など、収穫後のすべての処理が挙げられる。この意味で、貯蔵中に使用されるガスの混合物は、フェノール化合物の合成を調節することができる。また、収穫時に植物を傷めると、防御機構としてフェノール化合物の産生が誘導される[93]。
これらすべての事実を考慮すると、果物や野菜のフェノール化合物のプロファイルは非常に可変的であることが明らかである。したがって、特定の地域で栽培され、決められた農法に従って栽培された地域の果物は、フェノール化合物のプロファイルが特異的であると考えるのが妥当である。異性ホルミシス理論に関連して、これらの植物ポリフェノールによって付与された外部シグナルは、ローカルな次期プロセスのために体を準備する可能性がある。これはすべて、他の果物と比較して生物にいくつかの利点を提供する可能性があるので、地元の製品の消費を強化する可能性がある。
3.3. ポリフェノールと病気
表S1に示すように、フェノール化合物を豊富に含む果物や野菜の有益な効果は多岐にわたっており、これらのファイトケミカルは抗炎症作用、抗酸化作用、抗がん作用、心保護作用、降圧作用などを発揮する。これらの効果の根底にあるメカニズムは、これらの化合物が炎症や脂質・エネルギー代謝に関連する基本的な生化学的経路を調節する能力に基づいている[94]。さらに、これらの化合物は、エピジェネティックなメカニズムを介してその作用を発揮することができる。例えば、これらの化合物はmiRNAの発現を調節することが観察されている[95]。
果物や野菜の摂取による健康効果は、生理活性化合物として作用することができる各植物に含まれる特定のポリフェノールに部分的に起因している。例えば、フラバノールの(+)-カテキンおよび(-)-エピカテキンは、リンゴ、ココア、ブドウなどの非常に一般的な果物に存在する主な生理活性物質の一部である[72,82,94,96,97,98,99,100,101,102,103,104,105,106,107,108]。しかしながら、オレンジやレモンなどの柑橘類においては、これらの活性は、フラバノン類のヘスペリジンおよびナリンゲニンに関連している[72,97,109,110,110,111]。ポリフェノールの健康効果は、主にその代謝産物に起因することを強調することが重要である。この意味で、ポリフェノールは、特定の酵素によって小腸および肝臓で広範囲に共役し、グルクロニド、硫酸塩、およびメチル化誘導体を生成する。ポリフェノールのほとんどは小腸では吸収されずに大腸に到達し、そこで腸内細菌叢の活動にさらされる[112,113]。したがって、植物中のポリフェノールの含有量に影響を与える農業的・技術的要因に加えて、微生物相や酵素活性などの宿主の内的要因も、これらの化合物の効果に影響を与えている可能性がある。
3.4. 摂食パターン
従来、果物や野菜は、それぞれの品種の気候条件により、限られた季節に生産・消費されていた。例えば、さくらんぼやイチゴの完熟は春に限定され、オレンジの完熟は秋冬に限定されていた。しかし、現在では、野菜や果物が一年を通して消費されるようになったグローバル化により、これらのパターンは変化しつつある。例えば、スペインは欧州連合(EU)のオレンジの主要生産国であり、推定55%を生産している。しかし、この生産量は秋から冬にかけての季節に限られている。そのため、それ以外の期間、EU内のオレンジは、南アフリカ、アルゼンチン、ウルグアイなどの南半球の国々から輸入されている[114]。これにより、年間を通じて市場でこの製品を維持することができる。さらに、温室での果物や野菜の生産は、一部の製品の季節性を避けることにも貢献している。例えば、トマトやキュウリは、この農業慣行により、一年中市場に出回ることができる[115]。しかし、輸入されるこれらの果物や野菜の中には、成熟する前に収穫されるものもあることを考慮しなければならない。また、温室で生産されるものは、伝統的に栽培されているものとは異なるポストハーベスト処理が施されている。このように、これらがフェノール合成に影響を与える重要な要因であることを考えると、フェノール化合物の含有量が変化し、その結果、それらを摂取することによる健康効果にも影響を及ぼす可能性がある。
4. ポリフェノールを多く含む果実の季節的摂取による代謝調節機構の解明
以上のように、それぞれの植物が収穫された環境によって(ポリ)フェノールの組成が異なることを考慮すると、ポリフェノールを多く含む果実を季節的に摂取することで、摂取時期によって生理や代謝の調節が大きく変化する可能性があると考えるのが妥当である。実際に、ポリフェノールの健康効果には、周年サーカディアンリズムとサーカディアンリズム(24時間周期)の両方が影響していることが確認されている。図3は、生体リズムの遺伝子制御、植物ポリフェノール組成の季節変動、健康季節効果の相互作用を模式化したものである。しかし、この事実についての情報はまだ不足しており、文献には乏しい研究が見られる。表1は、概日リズムと周年リズムによって変調された食事介入とその健康効果をまとめたものである。
図3 生体リズムの遺伝子制御、植物ポリフェノール組成物の季節変動、健康季節効果の相互作用
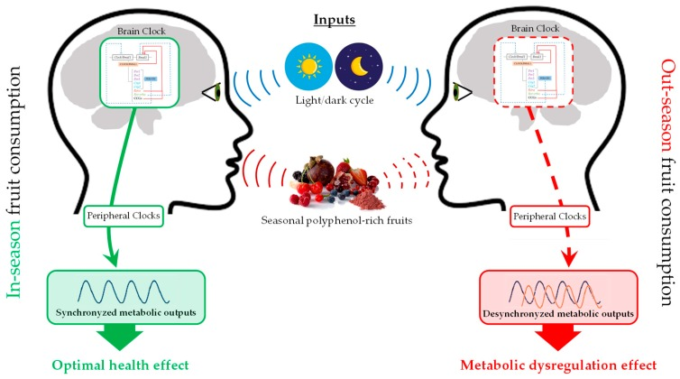
表1 食事介入とサーカディアンとサーカントリズムによって変調されたその健康アウトカム
| 食事介入 | 実験モデル | 健康成果 | タイムスケール | 参考文献 | |
|---|---|---|---|---|---|
| 概日リズムA | 年次リズムb | ||||
| 人間モデル | |||||
| カテキンが豊富な緑茶 | 健康な若い男性 | 食後の血漿グルコース濃度の低下 | 夕方(17:00 h) | na | [ 124 ] |
| ポリフェノールが豊富なブドウ-ワイン抽出物 | 軽度高血圧の男性と女性 | 歩行可能な収縮期血圧と拡張期血圧の低下 | 昼間 | na | [ 125 ] |
| 動物モデル | |||||
| エピガロカテキン-3-ガレート | C57BL / 6Jマウス | 肝臓および脂肪脂肪組織における概日時計遺伝子のリズミカルな発現を調節することにより、食事によって誘発される代謝の不整合を改善しました | 夜間 | na | [ 126 ] |
| ブドウ種子プロアントシアニジン抽出物 | オスのウィスターラット | 血漿メラトニンレベルの調節 | 昼間 | na | [ 121 ] |
| レスベラトロール | オスのウィスターラット | 酸化防止剤 | 夜間 | na | [ 127 ] |
| プロオキシダント | 昼間 | na | [ 127 ] | ||
| 赤ブドウ(伝統的な消費量:L6) | 標準(STD)給餌およびカフェテリア(CAF)給餌のオスのフィッシャー344ラット | 視床下部レプチン感受性の増加 | na | L6 | [ 118 ] |
| さくらんぼ(伝統的な消費量:L18) | STDを与えられたフィッシャー344雄ラット | 血中非エステル化遊離脂肪酸(NEFA)の減少 | na | L18 | [ 116 ] |
| 脂肪酸輸送、β酸化関連経路、循環グルコースおよびインスリンレベルの活性化の増加 | na | L6 | [ 116 ] | ||
| CAFを与えられたオスのフィッシャー344ラット | グルコース代謝に関連するCAF食の有害な影響の増大。 | na | L6 | [ 116 ] | |
| STD給餌およびCAF給餌のオスのフィッシャー344ラット | 視床下部レプチン感受性の増加 | na | L6 | [ 118 ] | |
a 昼間(光周期); 夜間(暗黒周期). b L6: 短日光周期(6時間/日); L18: 長日光周期(18時間/日) n.a. 情報なし.
4.1. 周年リズム
この意味で、食事誘発性肥満 F344 ラットを対象とした最近の研究では、スイートチェリー Prunus avium L.を 10 週間摂取すると、顕著な光周期依存効果があり、季節外れに摂取した場合には全身の脂肪酸化とグルコースとインスリンの循環レベルが増加することが示された [116,117]。これらの効果は、インスリンの下流ポストレセプター標的であるAkt2のリン酸化レベルのダウンレギュレーション、および骨格筋における脂肪酸輸送およびβ酸化関連経路の亢進によって部分的に説明された[116]。さらに、季節外れのPrunus avium L.の摂取は、白色脂肪組織の形態を変化させ、細胞面積を増加させ、脂肪細胞の数を減少させたが、これは主に脂肪組織の脂肪代謝に関与する主要遺伝子の発現のダウンレギュレーションに起因していた[117]。興味深いことに、褐色脂肪組織では、脂肪酸トランスポーターCd36もダウンレギュレーションされており、褐色脂肪細胞の脂肪吸収・酸化能力の低下(発熱活性の低下)を示唆している[117]。最後に、中枢神経系では、Prunus avium L.の消費もまた、季節外れの消費の場合にのみ、AgrpとPtp1B mRNAレベルを調節する視床下部レプチン系を調節した[118]。
同様に、同じ動物モデルでネベリナオレンジ(Citrus x sinsensis)を10週間摂取した場合も、脂質異常症やインスリン抵抗性、白色脂肪組織での脂肪酸合成の増加、褐色脂肪組織での脂質取り込みとβ酸化の調節の低下など、肥満誘発性の食事によって誘発される劇症的な効果を示した[119]。
関連して、いくつかの疫学研究では、生物学的リズムのずれが、肥満、脂質異常症、インスリン抵抗性、高血圧を含む多種多様な代謝障害にどのように寄与しうるかが示されている[120]。したがって、季節の果物の消費によって誘発される生理学的および分子的変化は、哺乳類の時計系の変調によって部分的に説明される可能性がある。実際、Prunus avium L.とCitrus x sinsensisの両方を季節外に消費すると、骨格筋のNr1d1,健康なラットの肝臓と白色脂肪組織のPer2とCry1などの異なる末梢時計関連遺伝子のmRNAレベルの変調と相関している[116,117,119]。これまでの研究では、食事誘発性脂質異常症やインスリン抵抗性に対して顕著な保護効果を持つフラボノイドのサブクラスであるブドウ種子プロアントシアニジン(PA)が、雄性Wistarラットの中枢および末梢の両方の生体リズムを有意に修飾することが実証されている[94]。具体的には、PA を 250 mg/kg 体重で急性投与すると、光期の初めにメラトニンレベル(夜行性)の上昇が維持され、健康なラットのいくつかの重要な循環代謝物のリズム振動が変化した [121]。この表現型の変化は、Bmal1やNamptなどの時計遺伝子の視床下部の発現パターンの調節と同時に生じた [121]。さらに、食事誘発性肥満ラットにおける4週間のPAの異なる生理的用量はまた、これらの動物の肝臓、腸、および白色脂肪組織において正の用量依存的な方法でBmal1,Nampt、Sirt1,およびNAD+などの末梢クロック成分のレベルを変調した[8,122,123]、PAは肥満状態における末梢および中枢分子クロックシステムを調整することにより、生理および代謝を変調することができることを示唆している。
4.2. 概日リズム
他のいくつかの植物(ポリ)フェノールは、中枢および末梢の時計システムに影響を与えることが記載されている。緑茶(Camellia sinensis)に含まれる主要なカテキンであるエピガロカテキン-3-ガレート(EGCG)は、体重の減少、肝臓および血漿中のコレステロールおよびトリグリセリドレベルの低下、およびグルコースのホメオスタシスの改善など、肥満に関連するパラメータに対して有益な効果を有することが実証されている[128]。特筆すべきは、EGCGの作用機序の探索において、Miら[126]は最近、C57BL/6Jマウスの概日時計遺伝子のリズミカルな発現を調節することにより、EGCGがこれらの肥満に関連した代謝変化を改善する可能性があることを初めて示した(表1)。具体的には、EGCGは、肝臓と白色脂肪組織の両方でSirt1とPGC1αのレベルを調節することにより、Clock、Bmal1,Cry1などの時計遺伝子の概日性発現レベルを部分的に正常化した。さらに、ブドウや赤ワインに含まれる別の体脂肪低下ポリフェノールであるレスベラトロールは、白色脂肪組織における時計遺伝子(Clock、Bmal1,Per2,およびRev-Erbα)と時計制御脂質遺伝子(Sirt1,Pparα、Srebp-1c、Acc1,およびFas)の両方のリズミカルな発現の高脂肪食の改変によって誘導される脂質代謝のサーカディアン・デシンクロニーを回復させた[129,130]。また、脂肪肝疾患の予防効果を示すエキナセア・プルプルプレア由来のポリフェノール成分である桂皮酸は、Akt/GSK3βシグナル伝達経路を増強し、脂質代謝に関与する下流の遺伝子発現を調節することにより、脂肪肝蓄積に対するBmal1抵抗性を示している[131]。最後に、果物に多く含まれるもう一つのフェノール酸である桂皮酸は、抗酸化作用や抗糖尿病作用で近年注目されている[132]が、分化した神経細胞の分子時計の概日周期を短縮したり、マウスの行動リズムの自由行動周期を短縮することが報告されている[133]。
これらの研究はすべて、肥満に関連した行動によって誘発される生体リズムの非同期性を回復し、最終的には重要な代謝関連転写物および代謝物のほとんどのリズム発現を正常化することによって、肥満に関連した代謝変化を打ち消すために、ポリフェノールが豊富な果物の消費のための戦略を設計するための貴重なデータを提供する可能性がある。しかし、(ポリ)フェノールが中枢および末梢の時計機構を調節する基礎的なメカニズムを解明するためには、さらなる包括的な研究が必要である。
5. 結論
フェノール化合物と生物学的リズムとの間の双方向の相互作用は、それらの消費に由来する有益な効果に強く影響する。この意味で、植物におけるこれらの化合物の合成を調節する様々な因子は、この種の植物製品の健康への影響を評価する際に不可欠である。旬の製品の消費に基づいて、製品が含む特定のフェノールプロファイルに起因する代謝の異なる変化がヒトで観察される可能性がある。この事実は、これらの特定のフェノールプロファイルの健康への影響に焦点を当てた研究の必要性を強調している。
