
Contents
- 概要
- 1. はじめに
- 2. ホルミシスの起源:Hugo Schulz
- 3. 伝統医学は閾値用量反応モデルを選択する:論争
- 4. 論争から用量反応革命へ
- 5. 確信的な複製の期間だが、共同の科学的活動はない
- 6. 1970年代後半の重大な転機
- 7. 1985年の次の重要なステップ
- 8. 毒性学におけるホルミシスの発展
- 9. これまでに得られた教訓とは?
- 10. ホルミシスの発生/頻度はどの程度か
- 11. 閾値やLNTよりも頻度は高いが、まだ限界がある
- 12. ホルミシスはなぜ生態学的・進化学的に重要なのか?
- 13. ホルミシスは発生に影響を与えるか?
- 14. 構成的なプロセスと適応的なプロセス:進化的、発達的に関連するもの
- 15. ホルミシスの用量反応研究を進化と発展の枠組みに統合する
- 16. 最後に
Theodosius Dobzhansky’s view on biology and evolution v.2.0: “Nothing in biology makes sense except in light of evolution and evolution’s dependence on hormesis-mediated acquired resilience that optimizes biological performance and numerous diverse short and longer term protective strategies”
Edward J. Calabresea,*, Evgenios Agathokleousb
a Environmental Health Sciences, Morrill I, N344, University of Massachusetts, Amherst, MA 01003, USA
b 生態学研究所,江蘇省農業気象重点実験室,南京情報科学技術大学応用気象学部(中国江蘇省南京市
概要
ホルミシスと呼ばれる二相性の用量反応は、生物学的モデル、生物学的組織のレベル、エンドポイント、誘発物質、メカニズムに依存せず、非常に一般化可能である。ホルメット反応は、基本的にすべての細胞や生物において、構成的な反応と適応的な反応の両方を媒介する重要な役割を果たしている。本論文では、生物学および生物医学におけるホルミシス概念の起源の歴史的概観と、生態学、進化、発達におけるホルミシス概念の潜在的役割の両方を提供している。これらの統合的な知見は、生存、寿命、繁殖、学習・記憶、組織修復、生殖・協力、発生プロセスに影響を与える複雑な進化に基づく選択戦略をよりよく理解するための広範な科学的枠組みを提供し、多くの課題の存在下での回復力を提供する。
1. はじめに
ホルミシスの枠組みの中では、低用量のストレスは生物の健康を増進し、高用量のストレスは生物を抑制するという、二相性の用量反応関係が見られる(図1)。ホルミシスは、局所的なスケールから地球規模のスケールまで発生する基本的な用量反応現象として浮上しており、惑星の健康に影響を及ぼす可能性がある(Agathokleous and Calabrese, 2019, 2020)。このようなホルミシスの理解の進展は、ホルミシスが持つ生態学的・進化学的な意味合いを再考することを促している(Agathokleous and Calabrese, 2019, 2020; Costantini, 2019; Costantini and Borremans,2019; Costantini et al 2010)。
この論文では、ホルミシスのトピックを、より広範な進化と発展の研究に紹介する。生物学および生物医学におけるホルミシスの概念の起源を歴史的に概観し、生態学およびヒトのリスク評価におけるホルミシスの潜在的な役割を探る。そして,ホルミシスの用量反応研究が,進化と発達の枠組みの中でどのように統合されているかを論じる。
2. ホルミシスの起源:Hugo Schulz
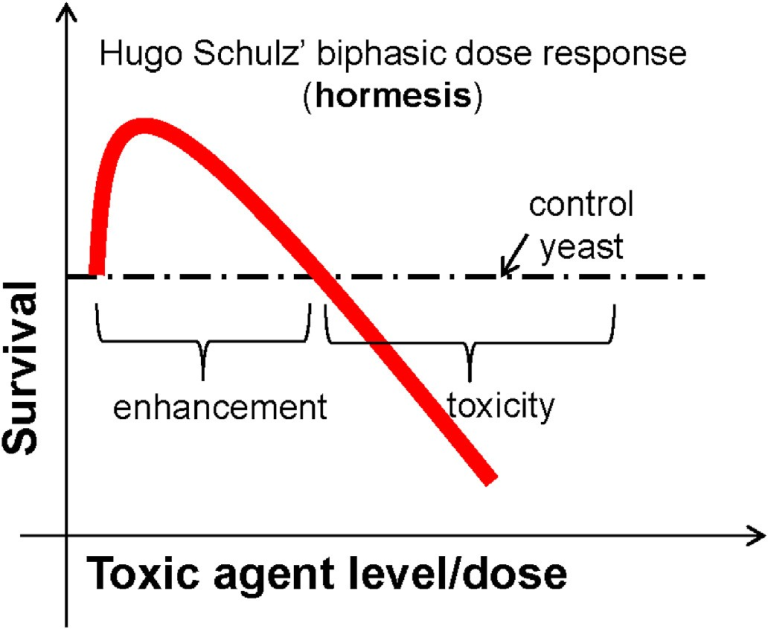
ホルミシスという概念は、1882年にドイツ北部のグライフスヴァルト大学で毒性学と薬理学を教えていた若い教授Hugo Schulzが、ほぼ10種類の「毒物」が低用量では酵母の代謝や生存を促進し、高用量では有害・毒性を示すことを予期せずに観察したことに端を発している(Fig.1)。Schulzとこの助手は、低用量での刺激反応が予想外であったため、実験手順に何らかのフラストレーションが生じたのではないかと考えた(Schulz, 1887, 1888)。薬品や実験系のテストを繰り返し行ったが、結果は同じで、二相性の用量反応であった。彼らは最終的にデータを追うことにし、その結果が非常に再現性の高いものであることを確信し、1884年に地元の医学会議でこの結果を初めて発表した(Crump, 2003)。初年度の若い教授にとって、この斬新な発見は多くの好機を生み出す可能性があった。しかし、シュルツにとって、この物語は、彼の長い職業人生全体に影響を与えるような、予想外の劇的な展開を見せることになった。
シュルツにとって状況が変わったのは、1885年に同僚のルドルフ・アーントとの出会いがきっかけであった。彼らは、Schulzの用量反応性の結果だけでなく、他の2つの観察結果も検討した。1つ目は、1884年にホメオパシーの文献に掲載された、化学物質ベラトリンが胃腸炎の患者の治療に成功したという論文である(Bloedau, 1884)。次に、このベラトリンの論文を読んだシュルツは、ロバート・コッホがこの病気の原因菌であると示したばかりの細菌のコロニーを手に入れた。シュルツはベラトリンが実際に胃腸炎の原因菌を殺すことができるかどうかを実験し、ベラトリンの臨床的観察結果を実験的に確認しようとした。その結果、ベラトリンは投与量にかかわらず、細菌に作用しないことがわかった(Bohme, 1986; Schulz, 1885)。このようにベラトリンが胃腸炎の原因菌を殺すことができなかったのは、この臨床研究のサンプル数が限られていたことから、臨床観察が偶然に起こったのではないかという疑問が生じたかもしれない。しかし、シュルツの場合はそうではなく、科学的な直感で別の道を歩んだのである。
図1 H. Schulzの発見を例にしたホルミシスの仮説例
SchulzとArndtは、これらのバラバラな知見を統合して、新たな用量反応仮説を構築した。まず、彼らは、たとえ原因物質を直接殺すことができなくても、ベラトリンの治癒効果は本物であるという信念を主張した。そうであれば、なぜ患者が治ったのか?そこで2人の科学者は、ベラトリンは細菌を殺すのではなく、生物の感染症に対する適応能力を高めることで作用すると推論し、Schulzの低用量酵母の二相性用量反応データを独創的に利用して、観察結果を新しい概念で統合し、二相性用量反応モデルを作成した(Crump, 2003)。彼らは、この説明で胃腸炎患者に対するバートリンの作用が説明できるだけでなく、ほとんどのホメオパシー薬が同じような二相性の用量反応で作用する可能性が高いと主張した。
この用量反応仮説の一般性は推測の域を出ないものであったが、シュルツとアーントは、当時、科学的、医学的、経済的に大きな対立を抱えていたホメオパシーの説明原理を発見したと主張し、この仮説は非常に大きな議論を呼んだ(Calabrese, 2005, 2011)。この時代、医学とホメオパシーの間の敵対関係が非常に激しかったため、シュルツはすぐに自分が大学や専門家の中で疎外されていることに気付いた。新しい用量反応理論は仲間から否定され、嘲笑と多面的な攻撃の対象となった。Schulzのキャリアは一夜にして変わり、彼の新しい理論は、非常に影響力のある科学者たちからかなりの反対を受けた。シュルツと彼の二相性用量反応モデルは、ホメオパシー運動の高希釈派と不当に結びつけられ、シュルツを過激派に仕立て上げ、その信頼性を失わせようとした(Bohme, 1986;Calabrese, 2011)。エディンバラ大学のアルフレッド・J・クラーク(Alfred J. Clark)のような非常に著名な学術研究者や学者は、彼の主要な薬理学や毒性学の教科書のいくつかで、シュルツと彼の用量反応理論についてかなりのスペースを割いており、非常に批判的であった(Clark, 1927a, 1927b, 1933, 1937)。普段は非常にバランスのとれた徹底したクラークの文章を詳しく調べてみると、彼のコメントは例年になく不公平であり、用量反応研究の記録を不均衡に表現していることがわかった(Calabrese, 2009, 2011)。
3. 伝統医学は閾値用量反応モデルを選択する:論争
ホメオパシーがライバルに勝って用量反応の概念を獲得したため、このような一連の出来事はすぐに伝統医学に独自の用量反応の信念を採用させることになる。ホメオパシーがライバルに勝って用量反応の概念を獲得したからである。自分たちの用量反応がライバルの二相性モデルであるはずがないので、伝統医学は最終的に、個人的な経験と実験的な裏付けがよく一致する閾値用量反応に落ち着いた(Calabrese, 2009, 2011)。このように、用量反応の概念は、一方の用量反応モデル(すなわち、閾値モデル)が生き残り、成功する機会を与えられた一方で、他方(すなわち、Schulzの二相性の用量反応)は、疎外され、否定され、嘲笑されるという論争の中で生まれた。シュルツ氏と彼のモデルを支持する傾向にあった人々は、権力者から専門的に脅かされ、他の人々が二相性の用量反応モデルを探求し、広めるのを妨げてた。シュルツにとって、二相性用量反応モデルが採用され、ホメオパシーと結びついたことで、グライフスヴァルトからドイツの他の著名な研究センターに移ろうとしていたことが頓挫してしまった。実際、シュルツは50年間の研究生活のすべてを、彼が始めた場所で過ごすことになる。長年の同僚は、シュルツの思い出として、用量反応モデルをホメオパシーに結びつけた結果、シュルツの職業生活がいかに困難なものになったか、また、明らかに学問的才能があったにもかかわらず、いかに待遇が悪かったかを、非常に興味深く、また、支持してくれている(Wels, 1933)。
4. 論争から用量反応革命へ
歴史は長く続くものだということを認識しておくことは重要である。しかし、医学とホメオパシーの間のこの長い間離れていて忘れられていた論争は、20世紀を通じて科学者が用量反応の性質について教育を受ける方法を変えたのである(Calabrese, 2011)。シュルツはそれを目にすることなく生きていたが、彼の苦闘した二相性線量反応理論は1970年代半ばに再発見され、1980年代にはホメオパシーとは関係のない形でさらに関心を集め、現在では低線量域での線量反応の性質をよりよく理解することを中心に、一種の線量反応革命を起こし、この二相性の概念が生物学、医学、公衆衛生の様々な側面を変革する可能性を秘めている(Calabrese and Mattson, 2017; Agathokleous and Calabrese, 2020)。) この用量反応革命の中心にあるのは、細胞、臓器、生物がどのようにして適応メカニズムをアップレギュレートするかであり、その結果、時間的にフレーム化された後天的な回復力が得られる。過去30年間、生物学および生物医学の文献にホルミシスが盛んに記録され、微生物、植物、動物で広く見られるようになり(Calabrese and Baldwin, 2000a,b,c,d,e)毒性学および薬理学の文献にも頻繁に登場し(Calabrese and Blain, 2005, 2009, 2011)その定量的特徴(Calabrese er al)。 このような発見は、進化におけるホルミシスの役割を検討するための健全な基盤となり、最近のかなりの努力を刺激している(Agathokleous and Calabrese, 2020; Costantini, 2019; Costantini er al)。
5. 確信的な複製の期間だが、共同の科学的活動はない
Schulzが直面した課題にもかかわらず、彼の線量反応のアイデアは影響力を持ち、多数の学位論文や学術誌への掲載など、かなりの研究を刺激した(Calabrese and Baldwin, 2000a,b,c,d,e参照)。ロチェスター大学の細菌学者であるSarah Branham (1929)は、実験室の条件を大幅に改善してSchulzの発見のすべてを再現し、Schulzのデータを確認するとともに、これらのケースでの二相性用量反応は、最初の控えめな用量依存性毒性反応に対する過剰補償として起こるという以前の提案をはるかに明確にした。この時代、二相性の用量反応研究は、重金属や電離放射線の植物、微生物、昆虫への影響に関する研究に強く支配されていた。これらのモデルでは、この二相性現象の用量反応の特徴をよりよく研究するために、より多くの用量を使用することができたからである。このような歴史的文献の評価は、当時の経験豊富な著名な研究者が幅広く参加してかなり広範囲に行われたが、これらの情報は批判的に検討されたり、十分に統合されたりしなかった。また、このような研究成果を支援し、今後の進め方を検討するための会議、議事録、専門的な社会活動など、二相性の用量反応関係に関する活動を組織しようとする試みもなかった。実際、この初期のホルミシス研究の時代に最も生産性の高かった20数名の研究者の職業生活を詳細に評価したところ、ほぼ全員が他の職業活動に方向転換し、特に様々な連邦機関での学術的なアドミニストレーションや同様の活動に重点を置いていたことが明らかになった(Calabrese, 2009)。研究リーダーの継続性がなければ、ホルミシスの概念は発展しないであった。1943年に「ホルミシス」という言葉を生み出したChester SouthamとJohn Ehrlichでさえ、Southamはコロンビア大学の医学部に進学し、Ehrlichはミネソタ大学でペニシリンの大量生産の方法を模索するために戦地に赴いたため、すぐにこの再探索領域を離れてしまった。
6. 1970年代後半の重大な転機
ホルミシス線量反応は、1970年代後半に3つの独立した研究活動によって大きな転換点を迎える。電離放射線の分野では、Thomas Luckeyが放射線ホルミシスに関する膨大な文献をまとめた2冊のモノグラフを執筆した(Luckey, 1980, 1991)。毒性学の分野では、Tony Stebbing(1982)が同様の道をたどっていたが、環境/海洋毒性学に焦点を当てていた。この2人の研究者に加えて、Szabadi(1977)やDe Leanら(1979)などの薬理学のチームが、二相性の線量反応を広範囲に記録しただけでなく、そのような反応の力学的な根拠を示した。唯一の違いは、LuckeyとStebbingがホルミシスという用語を使ったのに対し、phar macologyグループはまだホルミシスという用語を使っていなかったことである。これらの研究者は、二相性用量反応の現代的な変化をもたらし、次の変化のステップに必要な科学的根拠を与えたのである。
7. 1985年の次の重要なステップ
ホルミシス概念発展のための重要な次のステップは1985年に起こった。放射線ホルミシスに関する会議(カリフォルニア州オークランド)と1987年にHealth Physicsに査読付き論文を発表した。これがきっかけとなって、会議のリーダーであった二人が雑誌「Science」でポイント・カウンター・ポイントの議論を行った(Sagan, 1989; Wolff, 1989)。これらの出来事は、その後、ホルミシスの科学的基礎を探求するためのマサチューセッツ大学/アマースト校のBiological Effects of Low Level Exposure (BELLE)という活動/グループの創設に直結し、その後、多数の出版物、ワークショップ、会議、複数の大規模なホルミシスの線量反応データベースにつながり、最終的には、いくつかの国際的な科学的ホルミシス学会や学術誌の創設につながっている。この分野の急速な発展は、1980年代のWeb of Scienceデータベースでは、hormesisやhormeticという用語が年平均17回引用されていたのに対し 2010年代では年平均7627回引用されていることからも推察される(Web of Science; search method: Topic; 14 April, 2020)。2019年だけでも、これらの用語は、広く多様な科学分野の研究領域から約12400回引用されていた(図2)。また 2020年4月14日時点で、「ホルミシス」または「ホルメティック」という用語の被引用回数の合計は93 762回で、58 687件の報告に引用されている(Web of Science;検索方法Topic)。このような構造的変化により、低線量域での線量反応の性質に関する情報が広範囲に展開されている。
図 2. 年ごとのホルミシスの引用数の合計
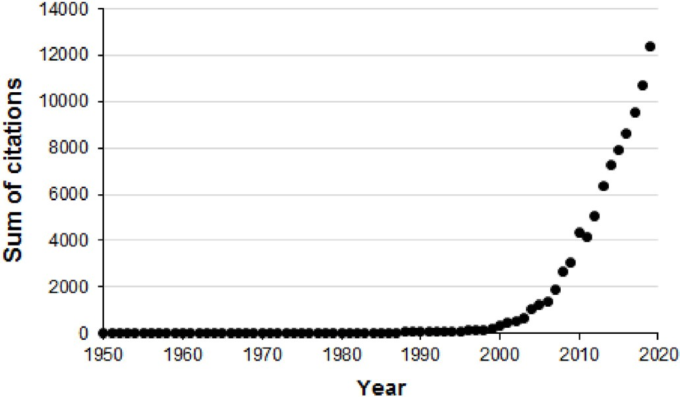
文献の評価は、Web of Science(2020年4月14日アクセス)で行った。キーワードは “horm- esis “または “hormetic”(検索方法Topic)を使用している。注:1980年代までの引用数がゼロなのは、当時の論文のテキストがWeb of Scienceに掲載されていないためと思われる。ホルミシスという言葉は1943年にSouthamとEhrlichによって提案されたもので(Southam and Ehrlich, 1943)、例えば1981年には3つの論文がホルミシスを引用しているが(Laughlin et al 1981; Stebbing, 1981a,b)、Web of Scienceでは1つの引用しか辿れない。したがって、実際の総被引用数は過小評価されている可能性が高い。
8. 毒性学におけるホルミシスの発展
ホルミシス分野の課題は、ホルミシスが毒性学の中でかなりの範囲で発展したことである。そもそも毒性学は、高線量-低線量の危険性評価を目的とした研究分野であった。このため、毒性学的な理解は、適応反応の領域にはほとんど関係のない線量域で行われてた。このような高線量試験は、低線量の影響に対する生物学的な誤解を招き、有害物質にさらされたとき、生体システムはしばしば受動的な受容者のように見え、誘発された損傷を修復することができなかったり、失敗したりするという意味合いを持ってた。しかし、1970年代初頭には、低用量の有害物質を事前に投与しておくと、その後10~14日程度の限られた時間内に同種または類似の有害物質を大量に投与した場合に防御されるという研究結果が発表され、毒性学における適応反応の分野が浮上していた(Gerhard et al 1972年、Glende、1972)。このような現象は、その後、化学的に誘発された突然変異反応についても報告された(Samson and Cairns, 1977)。電離放射線についても1920年代後半から同様の開発が行われており(詳細なレビューはCalabrese, 2016aを参照)1980年代前半には突然変異反応にも応用されるようになった(Olivieri et al, 1984)。プリコンディショニング効果の分野は、Murryら(1986)の報告により、低用量の心臓虚血ストレスを繰り返すことで、イヌが誘発した筋心筋梗塞によるその後の損傷や死から心臓を保護するという、臨床的に重要な意味を持つことから、より深く注目されるようになった。この結果は再現され、他の多くの生物学的モデルにも一般化された。このような低用量ストレッサーは、すぐにすべての細胞タイプと器官に保護を与えることが再確認され、定められた時間内に後天的な回復力を誘導する手段を提供した(Leak et al 2018,Stone et al 2018)。本論文で特に重要なのは、実験プロトコルで複数のプレコンディショニング用量を使用した場合に発生した。このような場合、結果は必ずホルミシスのような二相性の用量反応の発生を明らかにし、ホルミシスの概念をさらに一般化し、適応反応やプロサバイバルパスウェイやメカニズムと密接に結びつけることで(Calabrese, 2016b,c)ホルミシスに対する進化生物学的な関心をさらに高めることができた。
ホルミシスは、20世紀のほぼ全期間にわたって科学界から無視され、生物学、生態学、毒性学、生理学などの教科書には基本的に存在せず、生物学の中心的な概念であるにもかかわらず、教えられることがなかったことを考えると、重要な意味がある。今日でも、ホルミシスの概念やその生物学的、生物医学的な意義についての知識は、科学界では十分に評価されていない。例えば、プレコンディショニングの概念は1920年代から知られており、1980年代からはかなりの関心が寄せられていたにもかかわらず、ホルミシスの文脈の中でプレコンディショニングを統合したのは、数年前のことであった(Calabrese, 2017a,b)。
9. これまでに得られた教訓とは?
ホルミシスに関する過去数十年の研究で学んだことは、この現象は二相性の用量反応再関係であり、その振幅に関しては特定の定量的特徴を持つということである(Calabrese and Blain, 2005, 2009, 2011)。何千もの例を含む複数のホルミシス用量反応データベースが作成されると、ホルミシス刺激の振幅は一般的に控えめであることが明らかになった。これは、生物学的モデル、測定されたエンドポイント、誘導剤、誘導剤の効力、生物の年齢や性別、生物学的組織のレベル(すなわち、細胞、器官、生物)メカニズムにかかわらず、同様である(Calabrese and Mattson, 2017)。最大刺激は、対照群の反応に比べて約30~60%大きいことが一般的に観察されている。一般的に、最大のホルミシス反応の約80%は、対照群と比べて100%以下の大きさである。このような控えめな反応は、ホルミシス刺激の検出をバックグラウンドの変動と区別することを困難にする。このためには、低用量域での十分な数の用量を使用し、統計的検出力を高め、より多くの再現実験を行う必要があることが多い。現実的には、バックグラウンドの変動が比較的少ない対照群を使用することが好ましい。刺激量とは対照的に、ホルミシス刺激範囲ははるかに変動しやすいことが示されている。一般に、ホルミシス刺激量の範囲は、毒性学的/薬理学的に推定される閾値を起点として、約50倍以下であるが、刺激量の範囲が100倍を超えるケースも数多く存在し、ホルミシスデータベース(Calabrese and Blain, 2011)に登録されているよく特徴づけられたケースの約5%では、1000倍を超える可能性があるという。刺激範囲の変動が大きい理由は定かではないが、少なくとも部分的には、研究集団内の遺伝的異質性の程度に関連していると考えられる。つまり、刺激範囲の広さは、遺伝子の変異だけでなく、発達条件にも起因している可能性があるのである。集団内の異なる個体群は,異なる種類のストレスに対する感度や感受性の度合いが異なると考えられ(Agathokleous and Saitanis, 2020; Belz er al)。 個体群の刺激範囲の幅は、感受性の程度(ditogenic stage; Visser et al 2018)やその他の相互作用する環境因子によって異なるだろう。しかし、生物の競争力、防御力、生存力、ひいては環境での成功を後押しするであろうこの重要な側面は、実験的な考察を欠いている。したがって、刺激範囲の幅が、異なる個体群(遺伝子型、生態型、栽培品種、系統、種など)や、相互作用する環境条件・要因によってどのように変化するかを評価するための新たな実験研究が必要である。
ホルミシスによる刺激は,直接的な刺激反応によるものと,恒常性の乱れや軽度の毒性に対する適度な過剰補償によるものとがある(Calabrese er al 2007)。ホルミシスはホメオスタシスの乱れに対する過剰補償の結果として起こりうるので、ホルミシスを用量-時間-相関関係の中で研究することが必要である(Calabrese, 2001)。ホルミシスの用量反応がどのような過程で起こるかにかかわらず、最大の刺激は同様である。このような用量-時間反応現象は、ホルミシス研究における実験計画に時間と資源の両方の課題をもたらし、いくつかの重要な考慮事項を生み出する。第一に、潜在的なホルミシス反応は従来の閾値用量以下で起こるため、用量反応閾値を確実に推定することが必要である。このためには、一般的には、閾値の信頼できる推定値を得るための十分な予備的な用量反応実験が必要である。また、ホルミシス評価では、従来の毒性学的閾値以下の用量と、適切な用量間隔で閾値以上の用量を十分に設定する必要がある。最近、閾値以下の線量数とその間隔が、ホルミシスゾーンにおける最大刺激の推定値の正確性を大きく左右することが示された(Calabrese er al)。 これらの余分なステップは、ホルミシス性線量反応を研究するためには、より大きな再ソースと時間を必要とするため、よりDfficultになる。
10. ホルミシスの発生/頻度はどの程度か
ホルミシスが実験的にどのくらいの頻度で起こるのかという問題は、1990年代によく取り上げられてた。ホルミシスの例は、すぐに特定でき、再現性があることが示されているものが多くあった。これは貴重な情報であるが、頻度の問題には対処できなかった。頻度の問題を解決するためには、線量反応の客観的な先験的入力基準と評価基準を開発する必要があった(Calabrese and Baldwin, 2003a)。これにより、毒性学的な打率のような、文献中の頻度推定値を推定することができる。ホルミシスフリークエンシーを算出するための最初の試みは、3つの主要な毒性学・生命科学ジャーナルに掲載されたすべての論文に、先験的な入力基準を適用することであった。エントリークライテリアには、投与回数、推定閾値の発生、閾値以下のN個の投与量の存在などが含まれていた。そして、これらの線量反応は、厳格な統計的基準を適用した評価基準にかけられた。このプロセスでは、約21,000の論文が先験的な入力基準でスクリーニングされた。応募基準を満たした線量反応のうち、37%が評価基準を満たした。この研究により、文献上で初めてホルミシス頻度の推定値が得られた(Calabrese and Baldwin, 2001, 2003a)。その後、同様の戦略が他の大規模データセットに適用され、概ね同様の結果が得られた(Calabrese et al 2006,2010,2019b)。 これらの補完的なアプローチにより、ホルメチックな線量反応は稀な現象ではなく、ありふれたものであることが明らかになった。閾値用量反応モデルと比較すると(図3)ホルメティックモデルは閾値モデルに比べて2.5倍の頻度で評価基準を満たしていた(Calabrese and Baldwin, 2003b)。 線形非閾値(LNT)用量反応モデルは、非常に遠い3番目であった。
11. 閾値やLNTよりも頻度は高いが、まだ限界がある
上記の研究は、生物学および生物医学の文献において、ホルミシスの頻度がかなり高いことを確立した点で重要である。ホルミシスモデルがその2つの著名なモデルよりもはるかに高い頻度で出現したことは、皮肉なことではあるが、20世紀を通じて生物医学界で意図的に疎外されてきたのがホルミシスの用量反応モデルだったからである。ホルミシスの頻度が文献で確立される一方で、閾値用量反応モデルが環境・労働衛生基準、食品毒性基準、水生生物の水質基準などのリスク評価のために規制当局に採用される前に、科学界が閾値用量反応モデルをどのように検証しようとしていたかを評価する努力がなされた。しかし、どの国の政府内外の個人や科学者団体でも、低線量域(すなわち閾値以下)で正確な予測を行う閾値線量反応モデルの能力を検証する試みを発表した証拠は見当たらない。そのため、世界中の閾値モデル規制リスク評価プログラムは、このモデルが低線量域(すなわち閾値以下)における反応を正確に予測する能力を評価されることなく開発・採用された。人間のリスク評価のためのLNTモデルの使用についても、同様に詳細な再構築が行われたが、重大な脱落があり、現在では誤りがあることが明らかになっている(Calabrese, 2019)。LNTモデルは、ヘルマン・J・ミュラーが極めて高い線量のX線で遺伝子の突然変異を誘発したという仮定に基づいていた(図3)。しかし、現在では、ミュラーが誘発した遺伝子変異の証拠はほとんどなく、ほとんどがささやかなものから大規模な遺伝子欠失を誘発したと認識されている(Calabrese, 2017c)。しかし、シングルヒットのLNTモデルは、遺伝子変異が誘発されたというMullerの誤った判断に基づいていたのである。LNTモデルの歴史と規制機関による採用についての詳細な説明は(Calabrese, 2015, 2016a, 2019)を参照のこと。
第3図 H.J.Mullerの発見を例にした、閾値と線形非閾値(LNT)の用量反応モデルの仮想的な例

12. ホルミシスはなぜ生態学的・進化学的に重要なのか?
ホルミシスの最大値が、生物学的モデル、エンドポイント、薬剤などのあらゆる評価比較において類似しているという事実は、ホルミシスが進化の過程で高度に保存されているだけでなく、その基本的な量的特性と時間窓特性も同様であることを示唆している。刺激的な反応が控えめであることから、ホルミシスによる刺激は、重要なタイプの生物学的可塑性の限界を定量的に表しているのかもしれない(Calabrese and Mattson, 2011)。この反応の大きさの制限は,生物学的性能に影響を与えるプロセスの結果を制約する。このことは、記憶力を高める薬から、創傷治癒、骨折治癒、発毛などを促進する薬まで、製品開発を行っている製薬会社にとって、重要な意味を持っている。これらのケースでは、ホルミシスの用量反応の中で、薬剤がこれらの多様な基礎的なメカニズムのプロセスを向上させることができるという十分な証拠がある(Calabrese, 2013)。このタイムウィンドウの長さは、生体システムが危険信号を識別し、事前調整を開始し、限られた時間内に適応機構をアップレギュレートする能力を持っていることを示している。しかし、このような事前調整の時間的特徴の進化的基盤は、まだあまり注目されていない。
13. ホルミシスは発生に影響を与えるか?
発育は、遺伝性の遺伝子型と選択可能な表現型を結びつける基本的な役割を果たしている。前のセクションで述べた低レベルの応答戦略は,低レベルのストレスが発達に影響を与えることを示唆しており,この仮定は経験的な証拠によって裏付けられている。例えば,植物は,太陽からの光の取り込みや土壌からの栄養分の取り込みを有利にする生態戦略に応じて,地上部と地下部の発達に資源を配分するが,根の成長とシュートの成長(長さやバイオマス)の比率がホルメティックな反応を示すことが実証されている(Agathokleous er al 2019a)。このパターンは、低レベルのストレスが発達プロセスに作用して、地上部または地下部のどちらかの発達を有利にすることを示している。また,資源の枯渇を促す低レベルのストレスは,C-S-R戦略に依存して,植物の発達に異なる形で影響を与える可能性がある。例えば,競争戦略をもつ植物は,生産性が高く,かつ,ほとんど撹乱されていない系で生育した場合,低レベルのストレス下では,より多くの資源を競争に配分する(つまり,生産性が低下する)可能性があり,一方,ストレス耐性戦略をもつ植物は,生産性を維持または向上させる可能性がある。興味深いことに、成長の遅い植物は、成長の早い植物に比べて、ホルミシスをより一般的に、低用量で発揮する(Belz and Sinkkonen, 2019; Belz et al 2018b)。低レベルのストレスが、成長の遅い植物の成長と生存を促進することはあっても、成長の早い植物の成長と生存を促進することはないという事実は、低用量のストレスが、個体だけでなく、植物のコミュニティおよび/または集団の発達に影響を与えることを示唆している。この仮説は、さらに、異なる低線量のえさが、異なる影響を受けやすい集団のえさになることを示す研究によっても支持されている(Belz er al 2018b)。陸上の高等植物とは別に、藻類などの水生植物でもホルミシスが起こり、特に細胞の密度のホルミシス的な二相性の応答を示す形質がある(Zhang er al 2017)。
ホルミシスの発生学的な関連性は、動物についても実証されている。多くの研究者が示している顕著な例は、胎児が経験する低レベルのストレスが、ストレスに対するエネルギー消費と生殖との間のトレードオフを介して、後のライフステージや1世代または複数世代にわたって現れる世代間・世代間効果であり、これらはしばしば寿命の延長や生存優位性の向上につながり、その結果、自己および子孫のレジリエンスが向上する(明確なレビューはAgathokleous and Calabrese, 2020; Monaghan and Haussmann, 2015も参照)。 例えば,セレンが繁殖成功に及ぼす影響をアカシジミ(Agelaius phoeniceus)で評価したところ,セレン曝露地で餌を与えられた鳥は基準地で餌を与えられた鳥に比べて卵の孵化率が上昇した(Harding, 2008)。低レベルストレスの世代を超えた影響については,他にも多くの例があるが(Agathokleous and Calabrese, 2020),線虫のCaenorhabditis elegansでは,環境低レベルストレスが生殖細胞からソーマへのコミュニケーションを介して遺伝性の生存優位性を誘導することが明らかになった(Kishimoto er al)。 さらに最近では、除草剤メタミトロンの低用量投与が、特定の状況で望ましくないと判断された植物の、次世代における後続の処理に対する反応を変化させることが示された(Chenopodium album L.)(Belz, 2020)。また、親植物のプレコンディショニング時の刺激的な反応の大きさと、感受性やホルミシス発現の世代間変化の大きさとの間には正の相関関係が認められた(Belz, 2020)。このような世代間・世代間の影響については,例えば,一部の幼虫の段階で寿命が短くなるなど,負の影響と考えられるものもあるが,これらは,生存率を高めたり,後続の段階や世代でより高い生存確率を確保したりするための枠組みの中で,ある段階でのトレードオフを示しているに過ぎない可能性がある(Agathokleous and Calabrese, 2020; Feng er al)。 このような効果は、よりよく理解される必要がある。また、最近の研究では、イミダクロプリド抵抗性のシロイヌナズナBemisia tabaci Gennadius(Hemiptera: Aleyrodidae)の幼虫が除草剤を入れた土壌で栽培された植物を食べた場合、その幼虫から発育した雌の繁殖力が、条件付けされていない幼虫から発育した雌に比べて30~70%増加するような条件付けがなされていることが明らかになった(Rakotondravelo er al 2019)。ホルミシスは、変態、繁殖力、平均世代時間、総繁殖率、その他の発達過程についても観察され、多くの生物の様々な発達段階を占めている(Ayyanath et al 2015;Bejarano et al 2006;Brunettiら。2020; Cutler, 2013; De Carvalho et al 2013; Eldridge et al 1977; Hashmi et al 2015; Iavicoli et al 2018; Monaghan and Haussmann, 2015; Nascarella et al 2003; Sial et al 2018; Visser et al 2018)低線量ストレスが動物の発達過程に多面的に影響を与えることを示している。これらのすべてのeffectは、一般的にコントロールよりも30~60%の範囲である(Calabrese er al)。 しかし、このような低線量のえさを誘発するのは、生物学的ストレスだけではないことを明らかにしておく必要がある。例えば、種のfiitsnpelsasyを測定する一人当たりの人口増加率(Saitanis and Agathokleous, 2019)は、人口密度に対して典型的な二相性の反応を示すことから、ホルミシスプロセスは人口動態によっても起こりうることが示唆される。したがって、ホルミシスは世代間および世代内の複数のレベルで集団に影響を与える可能性がある。
ホルミシスと発生の関連性は、ミミズクについても示されている。例えば、浄水場で課される低レベルのストレスは、細菌のEscherichia coliにおけるバイオフィルムのフォーメーションを高めることに寄与することが示されている(Cui er al 2018)。また、低用量の環境ストレスは、微生物の生物発光反応を刺激し(Sun et al 2020)、細菌細胞の成長を促進することができる(Sun et al 2018, 2019)。微生物に対する低レベルのストレスのこのような発生的な作用は、農業の実践だけでなく、病気の媒介者に対する人間の曝露に関しても重要であると考えられる(Agathokleous and Calabrese, 2020)。
ホルミシスの観点から見ると,発生過程が様々な生命体で影響を受けることは明らかである。ホルミシス研究は、これまで生理学に焦点を当てていたが、近年の進展により、低線量ストレスの生理学的な影響が、個体やそれ以上のレベルでの発生プロセスにどのように結びつくかに、より焦点を当てることが可能になっていた。
14. 構成的なプロセスと適応的なプロセス:進化的、発達的に関連するもの
ホルミシスの用量反応は、事前調整と後天的回復力のプロセスの基本であることに加えて、正常な生理的・生化学的活動を導く一連の構成的プロセスを支配する受容体ベースのプロセスの機能の中心的な特徴でもある。ホルミシスの用量反応を介して作用する数多くの受容体は、広範な必須プロセスとエンドポイントに影響を与えることができ(表1)ホルミシスの概念はさらに統合され、一般化され、生存と最適な機能の中心となっている(Calabrese and Baldwin, 2001)。
以上の考察から、ホルミシスは、高度に構造化された/制約された用量反応の枠組みの中で、構成的なプロセスと適応的なプロセスを仲介する非常に一般的な戦略であることがわかる。ホルミシスの用量反応は、割り当てられた生物学的資源のレベルを制御・管理する一種のレオスタットのように機能し、可塑性の制約を受けるように反応を制限する。これは、ホルミシスの概念が再認識されるまでは認識されていなかった一般的な戦略であり、あらゆる生物のあらゆるレベルの生物学的組織の作用に影響を与え、しかもメカニズムや誘発物質に依存しないという驚くべきものである。ホルミシスが高度に保存されていることから、この概念とその生物学的プロセス・メカニズムを進化論的な枠組みでよりよく理解する必要があることがわかる。このような観点から、では、ホルミシスの用量反応研究を進化と発展の枠組みの中で統合する方法を検討する。
表1 二相性の用量反応関係が観察された受容体系の例
(Calabrese et al 2013)から引用したもので、いくつかの参考文献がある。オートファジー受容体p62/SQST-1(Kumsta et al 2019)など、さらに多くの受容体があり、その多くは関連するストレス反応とともに文献レビュー研究で取り上げられている(Calabrese, 2016a,b; Carvalho et al 2020; Macias-Bobadilla et al 2020; Muszyńska and Labudda, 2019; Poschenrieder et al 2013)。
レセプター系
- アデノシン
- 神経ペプチド
- Adrenoceptor
- ニューロテンシン
- ブラディキニン 一酸化窒素
- コレシストキニン
- N-メチル-D-アスパラギン酸
- コルチコステロン
- オピオイド
- ドーパミン
- 血小板由来の成長因子
- エンドセリン
- プロラクチン
- 上皮成長因子
- プロスタグランジン
- エストロゲン
- ソマトスタチン
- 5-HT スペルミン
- ヒト絨毛性ゴナドトロピン
- サブスタンスP
- インスリン成長因子I テストステロン
- ムスカリン性アセチルコリン
15. ホルミシスの用量反応研究を進化と発展の枠組みに統合する
生命はダイナミックな適応現象であり、小さな/穏やかな摂動から生命を脅かす課題まで、幅広い生物学的ストレス要因にうまく対処するための高度に保存されたプロセスを持っている(Koike et al 2018,Körner 2016,Oziolor et al 2019,Poschenrieder et al 2013,Vitasse et al 2013)。これらのプロセスは、生物システムがダメージを受けたり、タイムリーにダメージを回復したりするための脆弱性を説明する、非常に選択された膨大な量-時間反応の再関係に反映される。また、重大なダメージを受ける可能性のある状況を予測し、そのダメージを修復する能力(プレコンディショニング)は、生存率を高める上で重要な意味を持つ。一般的に、細胞から複雑なレベルの生体論理組織に至るまで、すべての生体システムは、ダメージをEfficientlyに修復し、典型的には、元の対照群の設定値を一時的に30%~60%程度上回るような、控えめなホメオダイナミックな過剰補償またはリバウンド効果を示す(すなわち、薬理学におけるciling effectの概念に相当する)(Calabrese er al 2019a)。上記で示唆されているように、この用量反応プロセスは、初期のダメージは線形の用量反応を反映しているかもしれないが、時間の経過とともに、低用量でのEfficient repairや他の代償プロセスを反映した二相性-ホルメティックな用量反応に変化するという時間的なダイナミックさを持っている(Agathokleous, 2018)。このプロセスでは、起こりうるダメージから身を守る多数の適応・修復機構がアップレギュレーションされるだけでなく、それらのアップレギュレーションが数日から数週間にわたって持続し、生命を脅かす可能性のある大きさの後続の大きな課題に対して、新たに獲得した回復力を介して、被害を受けた生体システムが保護されるように事前準備/適応/下準備が行われる(Calabrese, 2016b,c; Calabrese et al 2019b; Leak er al)。 適応機構の初期のアップレギュレーションと、その後のより大きな侮辱からの保護の定量的特徴は、ホルミシスの用量反応の定量的特徴と驚くほど一致している(Calabrese and Mattson, 2017)。心臓発作や脳卒中による損傷を防ぐことから、草食性(Belsky, 1986; McNaughton, 1983)に対する修復的代償反応(Stone er al)。 ホルメティックな用量反応のこれらの多数の非常に多様な例は、進化の枠組みの中でその一般性を示している。これらの同じ定量的特徴は、すべての器官系のすべての細胞型に作用する多数の受容体および非受容体システムによって媒介される膨大な一連の直接的な刺激/ホルミシス作用に類似している。このように、直接的な刺激、過剰補償、適応過程のための進化に基づく基本的な戦略は、ホルミシス的な用量反応の定量的な特徴によって記述することができる(Calabrese and Mattson, 2011)。この用量反応戦略は非常に一般的であり、細胞から器官、個体に至るまで、あらゆる形態の生命と生物学的レベル・組織の中心となっている(Agathokleous er al 2019c; Calabrese er al 2019a; Calabrese and Mattson, 2017)。
この進化的に保存されたホルミシスの用量反応関係の中で、用量反応の連続体全体にわたって、生物学的ストレスの概念が評価される (Agathokleous and Calabrese, 2020)。この分析により、生物学的ストレスは潜在的な損傷源であるだけでなく、すべての生物学的システムがシグナル伝達機構を介して数日から数週間の限られた時間内に将来の脅威を予測することを可能にするシグナルとなり得ることが明らかになった(Govindan et al 2018,Hunt et al 2011,Kishimoto et al 2017,Kumsta et al 2017)。これは現在、レドックス生物学の分野で見られる。生命が進化し、環境の自然および人為的な変化に対処するための現代および将来の戦略を導くのは、この一般的な枠組みの中である(Costantini, 2019; Oziolor er al)。 ホルメティック-二相性-用量反応のダイナミックさは、競争、協力、生存、選択の文脈で見られる(Costantini, 2019; Costantini and Borremans, 2019; Saitanis and Agathokleous, 2019)。これらは、同一個体や個体間の異なる器官内の膨大な数のホルミシス的な生体システムに反映されている。これらは、同様の基本的な用量-時間反応パターンに従う傾向があるが、活性化、パフォーマンスのEfficiencies、時間的枠組みにはプロ発見の差異がある(Calabrese et al 2012,Stranahan and Mattson 2012,Wang et al 2018)。これらの適応的な反応戦略は、生物の競争姿勢の中心であり、1種類の競争的な生物のホルミシス戦略が他の個体のそれと競合する状況的な進化的対立のfierce「ゲーム」に劣らず動作する。
このような進化上の競争シナリオにもかかわらず、すべてのシステムは、動的で、適応的で、補償的で、適度に過剰補償的で、現在のために保護し、なおかつ様々な発達精神的な発生窓の中で、またそれを越えて動作する潜在的な将来の脅威を予測するプロセスを進化させていた(Agathokleous et al 2019b;Calabrese 2016b,c;Costantini et al 2010;Oziolor et al 2019)。すべてが同様のホルミシス学的青写真に従っているようである。このように、ホルミシス的な用量-時間反応の概念は、毒性学、薬理学、公衆衛生だけでなく、進化生物学においても中心的なものであり、今後の研究の取り組みを組み立てる上で、中心的で根拠のない文脈を提供している。
16. 最後に
進化生物学者のテオドシウス・ドブジャンスキーは、「進化の観点以外、生物学では何も意味をなさない」という有名な言葉を残している(Dobzhansky, 1973)。ここでは、ドブジャンスキーの見解の第二版を紹介する。生物学では、進化と、生物学的パフォーマンスを最適化するホルミシスを介した後天的回復力と、短期的および長期的な多数の多様な防御戦略への進化の依存性を考慮しなければ、意味をなさないのである。