www.nature.com/articles/s41559-020-1273-8
4,600件以上の研究から得られた新たな証拠は、環境ストレスの勾配に沿って臨界閾値あるいはティッピングポイントを普遍的に適用することに疑問を投げかけている。環境政策や環境管理において、決して超過することのない環境目標を特定することは困難であることが明らかになった。
ジョーン・ダドニーとキャサリン・N・スディングは、「世界は不動の不動の場所のように見えるかもしれない。しかし、そうではない」と、Malcolm Gladwellは『Tipping Point』の中で書いている1。ティッピング・ポイントという考え方は、長い間、学者、作家、識者の想像力をかきたててきた。著名な論文では、地球規模のティッピングポイント2、社会の崩壊3、生物多様性の急速な減少4が差し迫っていることを警告している。これらの差し迫った崩壊の根底にあるのは、ティッピングポイント(一旦通過すると戻ることが極めて困難な閾値)の越境である。この閾値の超過は、天然資源管理に大きな影響を与え5、人新世において加速する変化の背景を作り出してきた。しかし、環境勾配に沿った転換点の位置を定量的に把握することは、あまり得意ではない、という厄介な現実が浮かび上がってきた。
Nature Ecology&Evolution誌に掲載された新しい論文では、臨界閾値の普遍的な適用に疑問を投げかけている。Hillebrandら6人は、環境要因に対する生態系の反応を、実験と観察の両方で調べた36のメタアナリシスを包括的に統合して発表している。環境ドライバーは多様で、富栄養化、気候温暖化、火災、窒素肥料などが含まれる。また、陸上、水中、海洋の各システムを対象に、独立栄養生物、従属栄養生物、微生物を対象とした4,600件に及ぶ研究が含まれている。
1つ目は、環境要因の強さが増すにつれて、特に環境ストレスのレベルが中間の場合、応答変数の分散が大きくなること、2つ目は、応答変数の効果量が多峰性分布に従うことで、状態遷移の典型的な兆候であることである。Hillebrandらは、環境ストレスの増加によって生態系が急速に変化する可能性があるという豊富な証拠を発見したが、反応は転換点付近で一直線ではなく、緩やかであった(図 1)。メタアナリシスの4分の1以下(36件中8件)が効果量のばらつきを示し、確認された増加は中程度の環境ストレスレベルではなく、極端なストレスレベルで発生していた。Hillebrandらは、さらに、Hartiganのdip検定法を用いて、双安定性を探った。36のメタアナリシスのうち、効果量の多峰性を示したものは1つもなかった。例えば、土壌呼吸に対する温暖化の影響に関するメタアナリシスでは、研究間で温暖化に対する反応が小さいことも大きいこともほとんど証明されていない。
図1|普遍的なティッピングポイントを示す証拠はほとんどない
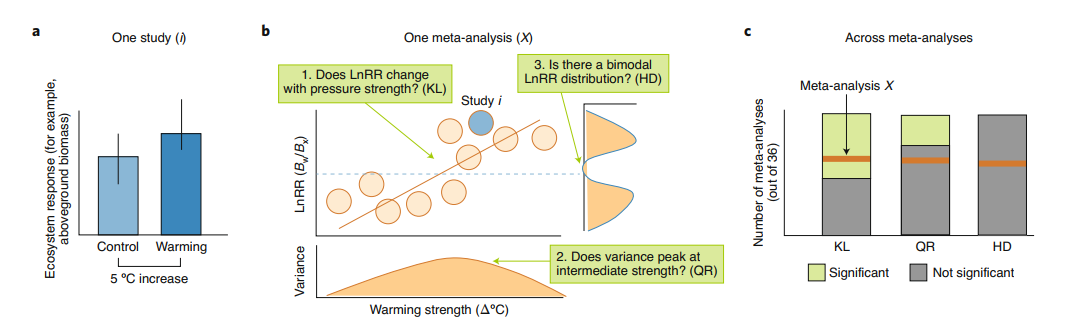
Hillebrandらは、広範な環境ドライバーに対する生態系の反応に焦点を当てた4,600件の研究の36のメタアナリシスを分析した。
a,各研究(i)について、ドライバーの圧力強度(例えば、温暖化度)に対する応答効果量(例えば、温暖化処理BwとコントロールBx間のバイオマスの自然対数変換応答比(LnRR)を抽出した。
b,次に、共通の反応軸と駆動軸で配列できる研究を組み合わせ(1つのメタ分析、X)、圧力の強さが増すにつれて閾値反応が発生するかどうかを示すかもしれない3つの質問を検証した。c,36のメタ分析にわたってこれらの検証結果を組み合わせたところ、著者らは仮説の閾値力学を裏付ける証拠は限られていることを発見した。Kullback-Leibler(KL)ダイバージェンス検定は、あるストレス因子値に対する反応分布と、ストレス因子値に差がないと仮定した場合の限界反応分布の間の全体的な偏差を定量化するものである。分位間範囲(5-95%)の重み付き分位比(QR)は、観察されたストレッサーの範囲にわたる反応分布の分散の実質的な非一様性を定量化するものである。Hartigan’s dip test method(HD)は、効果量の多峰性を評価するものである。圧力の強さの勾配に沿って反応を評価し、すべての生態系に普遍的な臨界閾値を仮定するこの手法は、本来この種の調査を意図していない研究デザインにおいても、転換点を調査する前例のない機会を提供するものである。
統計的検定の検出力を評価するために、Hillebrandらは、経験的データと、異なる応答-ストレス関係を自然変動を表すランダムな変動と組み合わせたシミュレーションデータとを比較した。閾値を検出する技術は、少量のノイズ(つまり、環境データに極めて一般的な系統的分散と確率的分散)を含むと性能が低下し、閾値を検出することができなくなった。
著者らは、分析したデータセットに閾値が存在したとしても、生態学的データに固有のノイズのため、検出は非常に困難であると結論付けている。
しかし、しきい値や転換点という概念を捨て去る前に、Hillebrandらが明らかにしたパターンからいくつかの疑問が浮かんでくる。まず、生態系の転換点は、ある次元には存在するが、他の次元には存在しないのだろうか?生態系のタイプ(例えば、森林、草原、湿地)、種の構成、主要な種や機能グループが、変化する要因に対して閾値反応を示すかどうかは、この論文のスケールではまだ検証されていない。広範な証拠によると、種構成のシフトはより閾値力学を起こしやすく、その転換点は環境圧力の異なる臨界値で発生することが示唆されている7,8。ある次元での閾値シフトが、必ずしも別の次元での閾値シフトを引き起こさない理由をさらに解析することが不可欠である。
第二に、特異な環境勾配に沿ってティッピングポイントが発生すると考えるべきなのだろうか。
ほとんどの研究は、単一のドライバーの影響に焦点を当てているが(それゆえ、Hillebrand et al.が利用できるメタ分析)、パルス「トリガー」とゆっくりとした変化「プレス」ドライバーとの相互作用が急激な変化を引き起こすことを示す証拠が増えている9,10。生態系が単独で環境ドライバーに反応することはほとんどなく、複数のドライバーの相互作用(気候変動と撹乱など)が自然システムでますます一般的になってきている。相互作用する複数のドライバーが含まれていることは、Hillebrandらが示唆するよりも、より頻繁に閾値の変動が起こることを示しているのかもしれない。
最後に、生態学的な偶発性(例えば、過去の撹乱の頻度や強度、土壌条件、空間的状況)は、普遍的な閾値と地域システムに特有の閾値のどちらに期待すべきかにどのように影響するか?偶発性によって、あるシステムが閾値を超える限界値が変化する可能性があり11、Hillebrandらがすべてのシステムに共通する閾値を期待することには疑問が残る。今後、「ノイズの多い」空間的・時間的偶発性を考慮した解析を行えば、システム固有の閾値が普遍的な閾値よりも一般的であることが判明するかもしれない。
Hillebrandらは、自然システムにおける普遍的な閾値に疑問を投げかけているが、彼らの結果は、潜在的により大きな問題を指し示している。もし私たちが、社会に大きな影響を与える可能性のある転換点の測定方法を知らなければ、人新世の急激な変化に対する備えが、私たちが思っている以上に不十分なものになってしまうかもしれないのだ。Hillebrandらは、生態系の変化を予測するためには、おそらく限界しきい値の設定や安全な作業空間の定義よりも、機構的な理解が重要であることを思い起こさせてくれる。生態系の感受性の根本的な原因、例えば、環境要因のわずかな変化が、どのように人口動態の急激な変化や生物学的・同生物学的フィードバックを引き起こすのかを調査することは、次のステップとして非常に重要である12。今後の研究では、幅広い非線形ダイナミクスの高度で微妙な理解と定量化を進め、社会が今後待ち受ける急速な変化に対応できるようにする必要がある。
Joan Dudney1 and Katharine N. Suding2✉1Department of Plant Sciences,UC Davis,Davis,CA,USA. 2コロラド大学北極圏・高山研究所生態学・進化生物学、米国コロラド州ボルダー。
参考文献
- 1. Gladwell, M. The Tipping Point: How Little Things Can Make a Big Difference (Little, Brown and Company, 2000).
- 2. Barnosky, A. D. et al. Nature 486, 52–58 (2012).
- 3. Ehrlich, P. R. & Ehrlich, A. H. Proc. R. Soc. B 280, 20122845 (2013).
- 4. Rockström, J. et al. Nature 461, 472–475 (2009).
- 5. Dudney, J., Hobbs, R. J., Heilmayr, R., Battles, J. J. & Suding, K. N.
- Trends Ecol. Evol. 33, 863–873 (2018).
- 6. Hillebrand, H. et al. Nat. Ecol. Evol. doi.org/10.1038/ s41559-020-1256-9 (2020).
- 7. Hallett, L. M. et al. Ecology 95, 1693–1700 (2014).
- 8. Berdugo, M. et al. Science 367, 787–790 (2020).
- 9. Benedetti-Cecchi, L., Tamburello, L., Maggi, E. & Bulleri, F.
- Curr. Biol. 25, 1867–1872 (2015).
- 10. Millar, C. I. & Stephenson, N. L. Science 349, 823–826 (2015).
- 11. Monger, C. et al. Front. Ecol. Environ. 13, 13–19 (2015).
- 12. Turner, M. G. et al. Phil. Trans. R. Soc. B 375, 20190105 (2020).
競合する利益
著者らは競合する利害関係を宣言しない。
