Contents
www.sciencedirect.com/science/article/pii/B9780128164105000050
Rethinking Food and Agriculture: New Ways Forward
2020年10月 ジョナサン・ラッセル・レイサム
バイオサイエンス・リソースプロジェクト、イサカ、ニューヨーク州、アメリカ合衆国

食料と農業の再考 新たな前進のための方法
Woodhead Publishing Series in Food Science, Technology and Nutrition(ウッドヘッド・パブリッシング・シリーズ・イン・フードサイエンス、テクノロジー、ニュートリション)。
ニューウェイズフォワード
編集:Amir Kassam
レディング大学、イギリス、レディング
ライラ・カッサム
お知らせ
この分野の知識とベストプラクティスは、常に変化している。新しい研究や経験によって理解が深まるにつれ、研究方法、専門的実践、または医療行為の変更が必要となる場合がある。
実務者や研究者は、本書に記載された情報、方法、化合物、または実験を評価し使用する際に、常に自身の経験と知識に頼らなければならない。このような情報または方法を使用する際には、自分自身の安全、および専門的な責任を有する関係者を含む他者の安全に留意する必要がある。
法律の及ぶ最大限の範囲において、出版社、著者、寄稿者、編集者は、製造物責任、過失、その他の問題、または本書に含まれる方法、製品、指示、アイデアの使用や操作による人または財産の損傷や損害に対して一切の責任を負わない。
目次
- 投稿者
- 感謝の気持ちと謝辞
- はじめに
- 1 農業におけるイノベーションの自由化 ルパート・シェルドレイク(Rupert Sheldrake)
- 1.1 はじめに
- 1.2 科学の聖職者
- 1.3 全知全能というファンタジー
- 1.4 唯物論の信憑性クランチ
- 1.5 分子生物学の果たせなかった約束
- 1.6 よりホリスティックなアプローチに向けて
- 1.7 イノベーションを自由にする
- 参考文献
- 2 農業は自然からの疎外感の種を植えた ジム・メイソン、ライラ・カッサム
- 2.1 はじめに
- 2.2 ドミニオニズム自然を支配する私たちの世界観とその起源
- 2.3 農耕の前に。生きている世界、包まれている世界
- 2.4 その他の動物人間の心や世界観を動かすもの
- 2.5 農業自然との新しい関係、新しい社会秩序
- 2.6 ミソザリー 動物や自然を減らすこと
- 2.7 牧場主の遺産。西洋の世界観の形成
- 2.8 戦争と植民地主義。国土の支配と他者への配慮
- 2.9 財産、貨幣、資本主義のルーツは牧民である。
- 2.10 ドミニオニズムとミソテリーの帰結
- 2.11 まとめ
- 参考文献
- 3 世界の食料・農業システムの政治経済学 フィリップ・マクマイケル
- 3.1 はじめに
- 3.2 現在の危機の前触れ
- 3.3 食のレジームの政治経済学
- 3.4 食品レジームコンプレックス
- 3.5 フードシステムの変遷
- 3.6 危機と解決
- 謝辞
- 参考文献
- 4 新植民地主義と食料安全保障と栄養のための新同盟。アフリカにおける開発の帰結に関するジェンダーの分析 マーク・ランガン、ソフィア・プライス
- 4.1 はじめに
- 4.2 南半球のドナー援助政策に対する批判としての新植民地主義
- 4.3 NAFSN:持続可能な開発か、それとも「新植民地」の押し付けの一形態か?
- 4.4 「開発」に対するジェンダーのレンズ。新植民地主義的なNAFSNがアフリカの女性たちに与えた影響
- 4.5 まとめ
- 参考文献
- 5 食料危機の神話 ジョナサン・レイサム
- 5.1 はじめに
- 5.2 GAPSの前提条件
- 5.3 モデルは何のためにあるのか?
- 謝辞
- 参考文献
- さらに読む
- 6 畜産批判としての動物倫理、環境主義、フーディズム、ロカボリズム、クリーンミート ロバート・C・ジョーンズ
- 6.1 はじめに
- 6.2 倫理学の基本。道徳的地位、道徳的価値、人間中心主義
- 6.3 現代の動物愛護運動の倫理的基盤
- 6.4 動物解放は決して三角関係ではなかった
- 6.5 フーディズムとロカバリズム。「人道的」な搾取の祭典
- 6.6 「クリーンミート」は畜産を終わらせるだろうか?
- 6.7 種差別の再定義
- 6.8 まとめ
- 参考文献
- 7 未来にふさわしいフードシステム トニー・ジュニパー
- 7.1 転換、集約化、劣化
- 7.2 インプットの影響
- 7.3 気候変動とより広範な食料システムの脆弱性
- 7.4 現行システムの推進要因
- 7.5 食料不足って?
- 7.6 私たちの食べ物は本当に安いのか?
- 7.7 偽のフレームに気をつけよう
- 7.8 役者
- 7.9 持続可能な食料システムのための優先事項
- 7.10 ガバメント
- 7.11 民間企業
- 7.12 コンシューマー
- 7.13 大きな問題
- 参考文献
- 8 なぜ、食料の栽培、加工、消費の方法を変えるのか? ハンス・R・ヘレン
- 8.1 はじめに
- 8.2 破綻したシステム。原因ではなく、症状に対処する
- 8.3 フードシステムの新しいパラダイム。軌道修正する方法
- 8.4 変化のためのフレームワーク
- 参考文献
- 9 科学の2つのパラダイム-そして科学に基づく農業の2つのモデル コリン・タッジ
- 9.1 パラダイムI:ハイテク産業ルート
- 9.2 パラダイムII:啓蒙的農業への道
- 9.3 アグロエコロジーの絶対的重要性
- 9.4 基盤を超えた再考察
- 9.5 補遺 I: 家畜の値段は?農場の「混農」は本当に必要なのか?
- 9.6 補遺Ⅱ:本物の農業と食文化が学べる大学
- 参考文献
- 10 農業のパラダイム アミール・カッサム、ライラ・カッサム
- 10.1 はじめに
- 10.2 産業的なグリーン革命農業のパラダイム
- 10.3 農業のオルタナティブ・パラダイム
- 10.4 代替農業のパラダイムは、生態学的にどの程度持続可能か?
- 10.5 構造的な問題についての考察
- 10.6 オルタナティブ・パラダイムへの道筋
- 10.7 包括的責任ある食料・農業システムという文脈における代替農業パラダイムの将来展望
- 参考文献
- 11 土壌の健全性とコンサベーションの革命的可能性 アグリカルチャー デビッド・R・モンゴメリー
- 11.1 はじめに
- 11.2 世界の土地劣化の程度
- 11.3 慣行農業の副作用
- 11.4 保全農業による土壌の健康づくり
- 11.5 第5の革命
- 参考文献
- 12 コンサベーションによる気候変動への適応と緩和 アグリカルチャー エミリオ・J・ゴンザレス=サンチェス、オスカー・ベロス=ゴンザレス、マヌエル・モレノ=ガルシア、マヌエル・R・ゴメス=アリサ、ラファエラ・オルドニェス=フェルナンデス、パウラ・トリヴィノ・タラダス、アミール・カッサム、ヘスス・A・ジル・リベス、ゴットリープ・バッシュ、ロサ・カーボーネル・ボジョロ
- 12.1 はじめに
- 12.2 気候変動と農業。なぜ農業のパラダイムを変える必要があるのか?
- 12.3 保全農業。気候変動を緩和し、適応する持続可能な農業システム
- 12.4 結論
- 謝辞
- 参考文献
- 13 遺伝子編集作物などのGM作物は持続可能な食料システムを破綻させるだろうか? アリソン・K・ウィルソン
- 13.1 はじめに
- 13.2 HT作物およびBt作物の影響
- 13.3 遺伝子組み換え作物における意図しない形質
- 13.4 新しいGM形質と技術
- 13.5 持続可能な農業と植物育種
- 13.6 結論障害と機会
- 参考文献
- 14 農業の生物多様性と異質な種子の持続可能性 パトリック・マルバニー
- 14.1 農業の生物多様性は意図的に異質なものである
- 14.2 農業の生物多様性の特徴的な特徴
- 14.3 農業の生物多様性への脅威
- 14.4 「世界を養う」作物は少ない?
- 14.5 農業生物多様性と種子の国際的ガバナンス
- 14.6 農民の種。より多く、より異質である
- 14.7 生物多様性と異質な農民アグロエコロジーの優先順位付け
- 14.8 農業生物多様性と種子システムの支配を争う3つの連合体
- 14.9 異質な種子と農業の生物多様性-責任ある農業と食料システムの基盤
- 参考文献
- 15 責任あるフードシステムの指針としての健康的な食生活 Shireen Kassam、David Jenkins、Doug Bristor、Zahra Kassam。
- 15.1 はじめに
- 15.2 現在の疾患傾向
- 15.3 健康・長寿に関連する食事パターン
- 15.4 セラピーとしての栄養の利用
- 15.5 間違った推奨をもたらした栄養科学へのアプローチ
- 15.6 栄養研究およびガイドラインに対する業界の影響力
- 15.7 健康的な食事と栄養を促進するための政府の介入
- 15.8 現在の食生活のトレンドが環境に与える影響を懸念している。
- 15.9 結論と提言
- 参考文献
- 16 包括的に責任ある食品と農業のための知識システム ロバート・チェンバース
- 16.1 はじめに
- 16.2 知識体系、言葉、意味
- 16.3 凝り固まった知識へのパラダイム的挑戦からの学び
- 16.4 農家とそのコミュニティ、自然資源を第一に考える
- 16.5 コーダ
- 参考文献
- 17 食と農のシステム変革における社会運動 ナッシム・ノバリ
- 17.1 はじめに
- 17.2 少数者のための企業の食糧体制
- 17.3 問題を知る:アグロエコロジーと欠落の物語を見抜くこと
- 17.4 都市活動家、動物解放運動家として踏み込む。 シード・ザ・コモンズの誕生
- 17.5 フード・ムーブメントにおける畜産の正常化と誤った二項対立の創出
- 17.6 層を剥がす。再生放牧と植民地物語の再生産
- 17.7 進むべき道ヴィーガンフードシステムを育てる
- 17.8 結論
- 参考文献
- 18 世界の食糧体制への代替案。システム変革への一歩 ヘレナ・ノーバーグ=ホッジ
- 18.1 グローバリゼーション 問題の根源
- 18.2 グローバリゼーションのコスト
- 18.3 ローカルへのシフト
- 18.4 未来のために、地元の食を
- 18.5 シフトを作る
- 18.6 貿易条約
- 18.7 補助金
- 18.8 課税
- 18.9 健康および安全に関する規定
- 18.10 幸福の経済学に向けて
- 参考文献
- 19 責任ある食品と農業のシステムを共創する ヴァンダナ・シヴァ
- 19.1 未来への2つの道
- 19.2 生きている土の再発見
- 19.3 インダストリアルパス
- 19.4 生命の道への回帰
- 19.5 未来の種を蒔く
- 参考文献
- 20 包括的な責任を果たすために ライラ・カッサム、アミール・カッサム
- 20.1 全体論的なパラダイムを目指して
- 20.2 豊かさの物語に向けて
- 20.3 農業の生態学的・多機能的パラダイムへ向けて
- 20.4 食と経済のシステムにおける権力の分散化に向けて
- 20.5 人と地球の健康を促進する食生活の実現に向けて
- 20.6 強力な社会運動と市民社会へ向けて
- 20.7 包括的責任のための倫理的枠組みに向けて
- 参考文献
- インデックス
投稿者
- Gottlieb Basch 欧州保全農業連盟(ECAF)、ブリュッセル、ベルギー;Instituto Ciências Agrárias e Ambientais Mediterrânicas, Universidad. Evora, Évora, Portugal
- Doug Bristor 独立研究員、ケンブリッジ、イギリス
- ロサ・カルボネル・ボジョロ IFAPAセンター・アラメダ・デル・オビスポ(スペイン・コルドバ)生態系生産・天然資源地域ロバート・チェンバース開発研究所(イギリス・ブライトン、サセックス州
- ヘスス A. ギル=リベスコルドバ大学農林工学部農村工学科、スペイン・コルドバ市、Asociación Española Agricultura de Conservación, Suelos Vivos (AEAC.SV), Spain マヌエル R. ゴームス=アリサスペイン・コルドバ市、Emilio J. ゴンザレス=サンチェス農林工学部、Asociación Española Agricultura de Conservación,Suelos Vivos (AEAC.SV),Spain ゴンザレス-サンチェスコルドバ大学農林工学部農村工学科、欧州保全農業連盟(ECAF)、ブリュッセル、ベルギー、スペイン、コルドバハンス-R-ヘレンミレニアム研究所、米国、ワシントンDC、米国
- David Jenkins トロント大学医学部栄養科学科、同医学部医学科、臨床栄養学・危険因子修正センター、Li Ka Shing Knowledge Institute、セント・マイケルズ病院内分泌・代謝科(カナダ、ON、トロント)。
- ロバート・C・ジョーンズカリフォルニア州立大学ドミンゲスヒルズ校(米国カリフォルニア州カーソン市)
- Tony Juniper Natural England(ロンドン)、Institute for Sustainability Leadership, University of Cambridge(ケンブリッジ)(英国)。
- Amir Kassam レディング大学(英国、レディング市
- Laila Kassam Animal Think Tank(ランカスター、イギリス
- Shireen Kassam キングス・カレッジ病院(ロンドン)、ウィンチェスター大学(ウィンチェスター、イギリス
- Zahra Kassam Stronach Regional Cancer Centre および Princess Margaret Cancer Centre; Radiation Oncology 部門(トロント大学、トロント、ON、カナダ
- Mark Langan Newcastle University, Newcastle upon Tyne, United Kingdom Jonathan Latham The Bioscience Resource Project, Ithaca, NY, United States Jim Mason Independent Researcher, Washington, DC, United States
- フィリップ・マクマイケルコーネル大学グローバル開発学部(米国ニューヨーク州イサカ市)
- David R. Montgomery ワシントン大学地球宇宙科学部(米国ワシントン州シアトル市)
- Manuel Moreno-Garcia IFAPA Centro Alameda del Obispo, Cordoba, Spain Patrick Mulvany Centre for Agroecology, Water and Resilience, Coventry University, Coventry, United Kingdom
- ナッシム・ノバリシード・ザ・コモンズ(アメリカ・カリフォルニア州・サンフランシスコ
- ヘレナ・ノーバーグ・ホッジエネルギー・資源グループカリフォルニア大学バークレー校(アメリカ合衆国)
- Rafaela Ordoñez-Fernandez スペイン農業保護協会(AEAC.SV)、IFAPA生態系生産・天然資源領域 Centro Alameda del Obispo, Cordoba, Spain Sophia Price Leeds Beckett University, Leeds, United Kingdom
- Rupert Sheldrake Schumacher College, Dartington, Devon, United Kingdom(英国)。
- ヴァンダナ・シヴァ科学技術とエコロジーのための研究財団とナヴダニア、ニューデリー、インド
- Paula Trivino-Tarradas コルドバ大学農林工学部農村工学科(スペイン・コルドバ) Colin Tudge オックスフォード・リアルファーミング会議およびリアルファーミングと食文化のためのカレッジ(イギリス・オックスフォード
- Oscar Veroz-Gonzalez Asociación Española Agricultura de Conservación, Suelos Vivos (AEAC.SV), Cordoba, Spain Allison K Wilson The Bioscience Resource Project, Ithaca, NY, United States
感謝の気持ちと謝辞
世界が直面している複数の危機の間にある点と点を結びつけ始めるのに、私たち二人は長い年月を費やした。これらの危機がなぜ、どのようにして発生したのかを問い、単なる症状ではなく、根本的な原因に対処するための可能な方法を見いだすために。この数年間は、農家や同僚、市民社会組織から多くのことを学び、批判的に考察し、学習していた。私たちは、さまざまな知識、専門技術、知恵、そして自然や動物の仲間たちから、互いに学び続けているのである。
この間、私たちは、より良い世界の創造に貢献するという目的意識、責任感、主体性を着実に高めていた。この感覚は、特に食料と農業の分野で、より持続可能で公正な方法を見つけるという私たちのコミットメントの根拠となり、それらが組み込まれ、絡まり合っているより広いシステムにおいても同様である。
世界の複雑さと私たちが直面している危機を考えると、私たちは、代替的な前進の道を垣間見ることができたことを幸運に思っている。この本を作る過程で、父と娘のチームとして、私たちの考えを探求し、統合する機会を与えられたことは、特に幸運なことだと感じている。また、本書に寄稿し、すべての人のためのより良い未来に向けた説得力のあるビジョンを提示してくれた著者たちによって、私たちの探求とアイディアに挑戦し、強化し、発展させることができたことに、心から感謝している。
本書の土台となった何世代にもわたる知的エネルギーと実践的な行動を十分に理解することはできない。私たちは、過去から現在に至るまで、世界の食糧と農業のシステムをより透明で、責任ある、包括的で、公正で、持続可能なものにするために、それぞれの方法で取り組んできたすべての人々に、特別かつ心からの感謝を表す。
また、私たちは、世界中の様々な進歩的な社会的・環境的正義運動に携わる人々にも深い感謝の意を表す。これらの運動は、個人の変革からより広範な構造的変革まで、さまざまな戦略を包含しており、私たちにとって大きな刺激と希望の源となっている。これらの運動と、それらが反映する感情の深さは、無視することはできないし、してはならない。
本書の作成にあたり、多くの同僚や組織から多大な援助と協力を得たことを認識したい。本書の編集を依頼した出版社エルゼビア社、特に本書の企画を考え、本書の範囲を明確にするために最初に協力してくれたナンシー・マラギオに心から感謝する。また、プロジェクトの様々な段階を積極的に管理し、様々な契約、スケジュール、フォーマット、システム管理に関して指導してくれたElsevierのMichael Lutz、Emerald Li、Vignesh Tamilselvvanに感謝する。
最も重要なことは、貴重な貢献をしてくれた主著者と共著者全員に深い感謝の意を表することである。皆さん一人ひとりが、私たちに多くのことを教えてくれた。本書は、そのような皆さんの意欲的な研究をまとめたものであり、大変光栄であるとともに、身の引き締まる思いがします。
また、本書の企画書および各章の外部査読者の方々には、本書が最高品質の資料で構成されるよう尽力いただき、心より感謝述べる。これらの査読者は以下の通りである。Raj Patel, Alain de Janvry, Uma Lele, Tim Wise, Per Pinstrup-Andersen, Nikos Alexandratos, Philip McMichael, Andrew MacMillan, the late Dick Harwood, Hans Gregersen, Norman Uphoff, Francis Shaxson, Thomas Welsh, Zahra Kassam, Shireen Kassam そしてParin Kassam。
最後に、私たちの愛する家族、愛する人たちに感謝する。この本のプロジェクト、そして世界における私たちのより広い活動に対する皆さんの支援と励ましは、私たちを希望と愛と感謝の気持ちでいっぱいにしてくれる。そして、より公平で、より優しく、より持続可能な世界は可能であり、私たち全員が果たすべき役割を担っているという確信を強めてくれる。
はじめに
Laila Kassama and Amir Kassamb aAnimal Think Tank, Lancaster, United Kingdom bUniversity of Reading, Reading, United Kingdom
私たちは、前例のない時代に生きている。私たちは、かつてないほど、自分自身や他の種の存在を脅かす重大な脅威に直面している。これらの脅威は、私たち自身が作り出したものである。1970年以降、人間の活動によって野生生物の個体数の60%が一掃された(Barrett et al.、2018)。現在、推定100万種の動植物が絶滅の危機に瀕している(IPBES、2019)。私たちは、絶滅率がそうでない場合の1000倍と推定されるほど、自然界に影響を与えている(Pimm et al.、2014)。生態系の90%近くが劣化または深刻な状態にあり、妥当な状態にあると推定されるのはわずか11%である(MEA, 2005)。人間の行動は、「第6の大量絶滅」事象を推進している「第1の大量絶滅」と呼ぶ方が適切な人もいる。「生物多様性の」消滅”は、地球の生命維持システムと私たち全員が依存している生態系を脅かす緊急事態である。
生物多様性と生態系機能の喪失は、気候の破壊と密接な関係がある。両者は互いに影響し合い、悪化させる。例えば、気候の破壊は生物多様性の損失の要因として浮上しているが、この損失は、気候の破壊によってますます一般的になっている異常気象に対する私たちの回復力を低下させている。両者がもたらすリスクは、同様に深刻である(IPBES, 2019)。1.5℃を超えるわずか半度の地球温暖化によって、何億人もの人々の干ばつ、洪水、火災、ハリケーン、猛暑、作物不作、貧困、避難、死亡のリスクが著しく悪化すると推定されている(IPCC、2018)。これらのリスクは、グローバルサウスの低工業国、低所得国には長年存在しており、グローバルノースの高工業国、高所得国と比較して、今後も不釣り合いに影響を及ぼすだろう。工業化の進んでいない国々は、この問題を引き起こしている責任が最も軽い国である傾向があり、過去数十年間、一人当たりの温室効果ガスの排出量は、豊かな国々に比べてはるかに少なかった。
最近の研究では、地球温暖化が世界経済の不平等を悪化させている可能性が非常に高く、気候変動の経済的不公正は少なくとも60年前から進行していることが示唆されている(Diffenbaugh & Burke, 2019)。私たちは、気候破壊という存亡の危機を好転させるためには、あと10年あまりしかないと言われている(IPPC, 2018)。したがって、私たちは、地球、私たち自身、そして地球を共有するすべての人々に対するこの破滅的な損害を防ぐことができる最後の世代なのである。しかし、一部の人によれば、私たちはすでに帰れないポイントを過ぎ、一連のティッピングポイントの閾値を越え、「惑星の非常事態」の状態にあるのかもしれない(Lenton et al.、2019)。
長期的な気候変動の影響は、様々な形で農業に大きな影響を与え、特に世界で最も脆弱な人々の食料安全保障に対するリスクを増大させると考えられる。熱帯・亜熱帯地域は、気候変動の影響を最も受けると予想される。先に指摘した嵐、洪水、干ばつなどの極端な気候現象の頻度と規模は今後も増加し、地球のあらゆる地域の農業生産性に深刻な影響を及ぼすと推定されている(Cline, 2007; González-Sánchez et al., 2017)。将来、居住可能な土地や天然資源をめぐる紛争が発生する深刻なリスクがあり、強制的な人間の移動が増加すると予想される。実際、近い将来の社会的崩壊はもはや避けられないと考えるアナリストもいる(Bendell, 2018)。Bendell (2018, p. 26)によれば。”最近の研究によると、人間社会は気候ストレスにより10年以内に基本的な機能に混乱が生じることが示唆されている。そのような混乱には、栄養失調、飢餓、病気、内紛、戦争のレベルの増加が含まれ、豊かな国を避けることはないだろう。”
気候変動は、人間活動がオーバーシュートした4つの惑星境界のうちの1つである。私たちの社会と生態系が依存している地球システムには、9つの境界線が存在する。オーバーシュートした境界のうち、気候変動と生物圏統合の2つは「中核的な境界」であり、大きく変化した場合、「地球システムを新しい状態に追い込み」、人類の幸福に深刻な影響を与える可能性がある(Rockström et al. 2009; Steffen et al.、2015)。
私たちの自然破壊は、相互に関連する多くの形態をとっている。私たちは過去150年間で地球上の表土の半分を失い(WWF, n.d.-a)、毎年240億トンを失っている(UNCCD, 2017)。表土は自然が補充できる速度の13~40倍の速さで浸食されており、現在の土地劣化の速度が続けば、60年以内に世界のすべての表土と現在の農業用作物がなくなるか、著しく浸食される恐れがある(Montgomery, 2007)。この表土が流水や風に乗って侵食されると、土壌の堆積物に加えて、肥料や農薬などの農薬や、水系や大気を汚染する微生物なども運ばれてくる(Juniper, 2015)。
また、森林も驚くべきスピードで消失している。人類の文明が始まって以来、私たちは46%の木を伐採し(Crowther et al., 2015)、過去50年間でアマゾンの熱帯雨林の約17%を破壊したと推定されている(WWF, n.d.-b)。しかし、これらの推定値には、最近のアマゾンの火災による破壊も、コンゴ盆地、米国、オーストラリアなどでの火災による破壊も含まれておらず、その荒廃の全容はまだわかっていない。
私たちは、食用や羊毛、毛皮、皮革などの製品のために、増え続ける陸上動物を殺している。1961年、私たちは食用として約70億頭の陸上動物を屠殺し、現在、年間約700億頭の陸上動物を殺している(卵産業で殺される雄雛は含まず)(Sanders, 2018)。また、養殖魚も毎年約800億匹殺している(Mood & Brooke, 2010)。
海の生命も、海洋酸性化や漁業など、人間の活動によって破壊されつつある。私たちは毎年1兆から3兆の野生の水生動物を食用として殺している(Mood & Brooke, 2010)。現在のペースで漁業を続ければ、2048年までに海から魚がいなくなると推定されている(Worm et al. 2006)。
人類が地球とそこに住むすべての人々に不釣り合いな影響を与えていることから、この時代(あるいは新しい地質学的時代)は「人新世」と呼ばれることが多くなっている。また、私たちが直面している危機における人間(アントロポス)や人間の本質とは対照的に、15世紀に始まった「安い自然」の創造に基づく資本蓄積の原動力を強調するために、「資本新世」と呼ぶ人もいる(Moore, 2015)。むしろ、アメリカ大陸、アフリカ、アジアを含むヨーロッパの植民地化をルーツとする西欧資本主義に連なる、ある種の破壊的な人間活動がその原因であると言える。果てしない資本蓄積が「地球全体を壊死させ…すべての生命を食い尽くし…その跡に種、言語、文化、民族の消滅を残す」(McBrien, 2016, p. 97)様子を反映して、「Necrocene」あるいは「New Death」と呼ぶべきとする意見もある。このように、資本の蓄積と消滅が同じプロセスであることから、「ネクロセン」はキャピタロセンの「影の二重人格」として考えられている(McBrien, 2016)。
しかし同時に、特にこの100年の間に、人間の努力の多くの分野で信じられないほどの物質的な進歩があったのも事実である。社会経済指標を見ると、識字率、長寿、所得の面で大きく向上していることがわかる。例えば、1917年の世界の識字率は23%だったが、現在では86%以上となっている。1900年以降、世界の平均寿命は2倍以上に伸び、現在では70歳を超えている。1870年から2016年の間に、世界の一人当たり平均GDPは10倍以上(それぞれ約1263国際ドルから14574国際ドル)になったと推定されている(Our World in Data, 2020)。しかし、IPBES(2019)の報告書にもあるように、この一人当たりGDPの増加が示す世界的な消費財の生産の増加と、それ以外の自然から人への貢献の低下は、直接関係している。
世界は、需要の増加に対応するために、自然からの物質的な貢献の流れを加速させるために、ますます管理されるようになっている。1970年以降、世界の人口は倍増し、一人当たりの消費量は45%増加し、国内総生産(GDP)で測定される世界の経済活動の価値は300%以上増加し、世界貿易は900%以上増加し、自然からの生物材料の採取は200%以上増加した。(p. 2)
さらに、こうした物質的な進歩は自然を犠牲にしてきただけでなく、暴力、戦争、土地や資源の収奪、人間や非人間の移動、搾取、殺害の増加を伴い、それらに関連しているのである。
全体として、良いニュースはあまりないと言えるだろう。ただし、オックスファムによって、地球の最貧困層の半分(約38億人)と同じ富を持つと推定される26人の億万長者については別だろう。これらの億万長者は 2000人以上の他の人々とともに、2018年にその富が12%増加し続けたのに対し、最も貧しい半数はほぼ同じ割合(11%)でその富が減少した(Lawson et al.、2019)。これらの推定値は議論を呼ぶかもしれないが、不平等が拡大している傾向はそれほどでもなく、経済成長の恩恵は富裕層によって不釣り合いに取り込まれていると推定される(1980年から2016年の間に、人口の最貧困層50%は世界の所得増加の1ドルあたり12セント、上位1%は1ドルあたり27セントだった)(Lawson et al.,2019).
1990年以降、ミレニアム開発目標(MDGs)が貧困と飢餓を半減させることに成功したという良いニュースの語り口にもかかわらず(MDG, 2015)、独自の分析では、「現実には、今日も約40億人が貧困状態にあり、約20億人が空腹のまま-歴史上かつてないほど多く、国連が信じさせようとしているものの2倍から4倍」(ヒッケル、2016)だという。
世界は100億人以上を養うのに十分な食糧を生産しているにもかかわらずです(Holt-Giménez, Shattuck, Altieri, Herren, & Gliessman, 2012)。飢餓と栄養不良が増加する一方で、今やパンデミックと認識されている肥満の流行も、先進国だけに留まらない。2016年、約20億人の成人が太りすぎ、6億5千万人が肥満であった(WHO, 2018)。Lancet Commission on Obesityによると、肥満、栄養不足、気候変動のシンジテックは、人間、環境、地球にとって最も重要で緊急の課題である(Swinburn et al., 2019)。
食料と農業のシステム問題の一部と解決策
生態系や気候の非常事態、環境の悪化、不平等の拡大、貧困、飢餓、肥満などは、それぞれ別の問題ではない。それらは相互に関連しており、食料・農業システムの役割はそれらすべてにおいて中心的な役割を担っている。食料・農業システムとは、食料の生産、加工、流通、調理、消費に関連するすべての要素(環境、人間、非人間動物、投入物、プロセス、インフラ、制度など)と活動、および社会経済的・環境的成果を含むこれらの活動から得られる成果のことである。3つの中核的要素を持つ。(1)食料サプライチェーン、(2) 食料環境、(3) 消費者行動(CFS-HLPE, 2019)である。
IPBES(2019)報告書によると、土地利用変化(主に作物生産、飼育動物、プランテーションのための土地転換)は、私たちの自然破壊の最も強い推進力となっている。第一次世界大戦以降、食糧と農業のシステムは、現在支配的な産業的グリーン革命の農業のパラダイム(第10章)を開発した企業部門によってますます推進されてきた。この農業生産モデルは、集約的な耕作、近代的な種子、農薬と化石燃料の過剰使用、単一栽培、あるいは多様性に欠ける作付体系に基づいている。これらのシステムは、農業の生物多様性に乏しく、生態系の持続可能性を保証するものではない。また、生産性や投入資材の利用効率は最適とは言えず、生態系社会サービスはほとんど提供されず、気候変動に起因するものを含め、生物・生物学的ストレスやショックに対する適応力や回復力に乏しい。また、温室効果ガスを大量に排出し、気候変動を緩和するどころか助長している(Brisson et al., 2010; Kassam, 2020; Kassam et al., 2013, 2017; Kassam, Friedrich, Shaxson, & Pretty, 2009)。先に述べた破壊の多くは、工業的農業とそれを推進する企業の食糧体制に直接関連している(第3章)。
野生生物の損失の約60%は、農業と私たちが用いる工業的手法に関連していると推定されている(Barrett et al.、2018)。従来の工業的農業は、昆虫の80%、作物の多様性の93%を絶滅に追いやった(Shiva et al.、2017)。土地利用(土壌劣化や森林破壊の影響を含む)は、世界の温室効果ガス排出量の4分の1から3分の1に寄与すると推定されている(IPCC、2019)。飼料、農薬の製造、加工、輸送を含む食料システムは、世界の年間温室効果ガス総排出量の30%以上を占めると推定されている(Bajželj, Allwood, & Cullen, 2013; Herrero et al., 2016; Vermeulen, Campbell, & Ingram, 2012)そのうち60%以上は畜産から発生している。畜産は、すべての輸送手段を合わせたよりも多くの温室効果ガスを排出すると推定されている(Bailey, Froggatt, & Wellesley, 2014)。これは、世界最大の国民経済国である米国が排出する排出量よりもかなり多い。FAO(2013)によると、温室効果ガス総排出量の14.5%を養殖動物が占めている。また、畜産は食品関連の温室効果ガス排出量の少なくとも半分を占めている(Herrero et al., 2016; Vermeulen et al., 2012)。世界では、食肉・乳製品企業の上位20社は、ドイツ全体よりも多くの温室効果ガスを排出している(Heinrich Böll Stiftung, GRAIN, & Institute for Agriculture, & Trade Policy, 2017)。
従来の耕作ベースの農業は、土壌の破壊の75%に責任がある(Montgomery, 2007)。過去70年間で、土壌浸食と劣化、土壌の健全性と生態系機能の喪失により、約5億ヘクタール(年間700万~1200万ヘクタールの割合で)の土地が放棄された(Gibbs & Salmon, 2015; Juniper, 2015; MEA, 2005; Montgomery, 2007; Nkonya, Mirzabaev, & von Braun, 2016)。集中耕作による農業のため、浸食によって毎年約250億トンの表土が失われている(Nkonya et al.、2016)(第11章)。畜産による化学肥料や汚水の流出は、世界中で400以上の水生デッドゾーンを作り出し(Diaz & Rosenberg, 2008)、水路や地下水の汚染に寄与している。また、農業は世界の森林破壊の約80%を占めており、その中でも畜産(動物飼料の生産を含む)は大きな推進力となっている(Campbell et al., 2017; Hosonuma et al., 2012; Kissinger, Herold, & De Sy, 2012)。アマゾンでは、森林破壊された土地の70~80%が放牧用の牧草地に転換され、残りの土地の多くが大豆などの動物飼料の栽培に使われている(Machovina & Feeley, 2014; Steinfeld et al, 2006)。畜産は、地球上の生息地喪失の最も大きな要因であり(Machovina, Feeley, & Ripple, 2015)、世界の生物多様性喪失の最大の要因の一つである(Steinfeld et al. 2006)。肉、養殖魚、卵、乳製品の生産は、農地の約83%を使用する一方で、カロリーの18%しか提供していない(Poor & Nemecek, 2018)。さらに、集中的な工業的畜産は、飼料生産と野生動物取引のための関連する生息地の破壊とともに、新規の人獣共通感染症の拡大につながっている(Morse et al.、2012)。工業的畜産の継続は、さらなるパンデミックの発生を不可避とする(Ceballos, Ehrlich, & Raven, 2020)。
しかし、特に第二次世界大戦以降、自然破壊を推進してきたのは、農地利用の変化だけではない。これらの変化は、それに伴って発展してきた工業的農業生産システムとともに、消費者の食品と食生活への需要を喚起し、環境破壊だけでなく、肥満、がん、心臓病、糖尿病などの非感染性疾患、一般的な不健康の増加といった健康への悪影響をもたらしている。したがって、私たちの食品消費の選択は、土地利用の変化だけでなく、こうした環境や健康への影響を促進する上でも大きな役割を果たしている(Poor & Nemecek, 2018)。
それでも、これらの影響は、純粋に消費者の選択に起因するものではない。政府や国際機関によって促進される企業の食糧体制は、様々な形で消費者の選択に影響を与え、これらの環境と健康への影響を促進する上で中心的な役割を果たしている(3章)。例えば、食品システムは、すべての人に健康的で栄養価の高い食品を提供できないだけでなく、加工食品、砂糖、塩、油、動物性食品が多く、栄養価の高い植物性食品全体が少ない欧米の食事パターンを促進しているため、慢性的な飢餓の割合が高く、肥満が蔓延する両方の中心となっている(Willett et al., 2019)(15章)。それに伴い、人間の健康と農業の持続可能性の両方に必要であるとして産業や社会が推進する動物性食品の消費の増加により、食料穀物と土地の大半が畜産に割り当てられ、その多くが集約的な産業システムのもとで行われている。これらのシステムは、森林や生物多様性を破壊し、温室効果ガスの排出や気候・生態系の破壊を引き起こし、ある地域では社会から疎外された人々や先住民から土地や生計を奪い、ホームレスにしている。
結局のところ、天然資源基盤への圧力は、食糧と飼料に対する社会の需要の大きさと性質、そしてそれらが国内外においてどのように生産、加工、流通、利用、廃棄されているかに密接に関係している。同時に、一般的な食料・飼料システムの主要な政治的・経済的原動力は、農業システムを産業的・侵入的・技術指向の道へとさらに押し進め続けている。この道は、農業生態学的に指向され、再生可能で持続可能な農業生態系のプロセスと結果に基づくものではなく、環境的に劣化し、経済的に持続不可能になっている(第10章~12章)。
環境や健康への影響もさることながら、貧困に苦しむ人々の大半が農村部に住み、その大半が農業に従事していることを考えると、農業は貧困や飢餓にも直接関係している(Castaneda et al.) 世界の5億7000万以上の農場のほとんどは小規模で家族経営の農場であり、低工業国では25%の農地で最大80%の食料を供給していると推定されている(FAO, 2014; FAO-IFAD, 2019)。貧しい農民農家が世界の食料のほとんどを生産しているにもかかわらず、そのほとんどが飢えている。
現在の食料・農業システムが、私たちが直面している危機の重要な推進要因であることを考えると、それを持続可能で民主的なシステムに変革することも、解決策の重要な部分となりうることは当然である(第7章、第8章)。私たちの考えでは、多くの危機の最終的な原動力であるグローバルな新自由主義的企業資本主義経済システムを支え、促進する上で、企業の食糧体制が大きな役割を果たしていることから、その変革がシステム全体の変革につながる大きな可能性を秘めていると考えている。
持続可能な食料・農業システムは、「将来世代の食料安全保障と栄養を生み出す経済的、社会的、環境的基盤が損なわれないような方法で、すべての人の食料安全保障と栄養を確保する」ものと定義されている(CFS-HLPE、2019、p. 28)。ベースに生態学的に持続可能な農業の生産パラダイムがなければ、このようなシステムは不可能である。第二次世界大戦以降、産業的なグリーン革命の生産パラダイムが支配的であったが、いくつかの代替的でより持続可能なパラダイムが出現してきた。これらの代替パラダイムは、工業的農業が引き起こしてきた破壊と劣化、および資源に乏しい大多数の小規模農民のニーズに適さないことに一部対応して発展してきた(第10章)。これらの代替的なパラダイムは、世界各地で運用され、食糧と農業のシステムに貢献している。産業的なグリーン革命のパラダイムが息切れしているのに対し、これらのオルタナティブ・パラダイムは力をつけてきている。
しかし、生産パラダイムは、より広範な構造的変化を伴わない限り、転換することはできない。工業的農業のパラダイムとそれがもたらす破壊の主要な推進力は、「自由貿易」協定と大規模かつグローバルなものに偏った政策(第18章)、そしてそれに関連する企業による世界の食料・農業システムの乗っ取り(第3章と第14章)によって推進される経済のグローバル化である。グローバリゼーション、資本主義、そして企業の食糧体制は、持続可能な開発とは相容れないものである。
主流な反応と語り
持続可能な開発の追求は、国連(UN)とその専門機関、そして多くの国際・国内開発機関、政府、ドナーにとって、何十年にもわたって継続的な目標となっている。1972年、国連は「人間環境会議」を開催し、環境問題を初めて国際的な議題とした。1983年、国連は、ノルウェーの元首相グロ・ブルントランドを委員長とする「環境と開発に関する世界委員会」を設立した。ブルントラント委員会は、農業開発を含む世界的な経済発展に伴う環境と天然資源の劣化がもたらす課題を再検討することを目的としていた。そして、これらの課題に対処するための行動計画を策定し、各国が持続可能な開発に向けて共に努力することを奨励することを目的としていた。1987年に発表された「ブラントランド報告書」には、現在でもよく使われる「持続可能な開発」の定義が含まれている。
しかし、人口の増加、都市化、ライフスタイルの変化、所得の増加、資源の消費と浪費、森林破壊、グリーン革命による農業など、環境と天然資源の劣化は深刻で、1992年に国連はブラジルのリオデジャネイロで「環境と開発に関する会議(地球サミット)」を開いた。気候変動、生物多様性、劣化に関する条約、アジェンダ21、「環境と開発に関するリオ宣言」、国連持続可能な開発委員会など、いくつかの条約が制定され、これらの課題に対処している。さらに最近のグローバルな取り組みとしては 2000年に策定されたMDGsや、20-30年までに達成するために2015年に設定されたSDGs(Sustainable Development Goals)がある。
しかし、これらの、そして他の多くの、国内外での「開発」(農業開発を含む)が生み出す環境破壊や不公平という課題への取り組みにもかかわらず、事態は悪化の一途を辿っている。私たちが見てきた「将来の世代が自らのニーズを満たす能力を損なうことなく、現在のニーズを満たす開発」として、気候や生態系の破壊、資源の過剰利用、貧困、飢餓はとどまるところを知らない(Bruntdland, 1987; Earth Summit, 1992; Foresight, 2011; IAASTD, 2009; MEA, 2005; Montgomery, 2007; SDG, 2018)。その理由の一つは、これらのイニシアチブの多くが、これらの危機の根本原因を明示的に特定できていないため、現実的かつ効果的な解決策を特定できていないことである。もう一つの理由は、主流の「開発」が実は植民地プロジェクトの継続に過ぎないという見方である。持続可能な開発や食料安全保障という言説が、企業によって貴重な資源へのアクセスに利用されており、「開発」は新植民地主義の一形態であるという意見もある(第4章)。
国連のSDGsは、貧困、不平等、気候変動、環境破壊、平和、正義など、私たちが直面するグローバルな課題に取り組むための現在の青写真である。国連のSDGsのすべての達成は、それ以前のMDGsと同様に、自然およびその直接的または間接的な人への貢献に依存している(IPBES, 2019)。自然破壊における食料・農業システム、特に従来の工業的農業のパラダイムの中心的な役割を考えると、また、進展が見られないことから、過去10年間、「従来通りのビジネス」からの脱却と食料・農業システムの根本的な変革の必要性を求める声が緊急かつ高まってきた(CFS-HLPE、2019;IAASTD 2009;IPBES、2019;WDR 2008)。しかし、政府、主流の開発組織、慈善活動家、企業の反応は、そもそも私たちをここに連れてきた産業農業と新自由主義資本主義経済学の同じパラダイムに基づき、同じことの繰り返しを提供しているように見える。
こうした還元主義的なパラダイムは、現在のシステムを維持・強化するいくつかの神話や物語を生み出してきた。これらの神話には、飢餓は食糧不足と人口過剰によるもの(第5章)、世界を養えるのは工業的農業と遺伝子組み換え生物(GMO)だけ(第13章と第14章)、有機農業とエコロジー農業では飢えた世界を養えない、自由市場で飢えをなくせる、自由貿易が答え、海外援助は飢えた人々を助ける最善の方法、権力は本当の変化をもたらすには集中しすぎている(第18章)、などがある。(Lappé & Collins, 2015). 他のすべての神話が依拠しているのは、欠乏や不足の神話である(飢餓や食糧生産に関連する欠乏だけでなく、資本主義経済システムが依拠し人為的に作り出した資源の欠乏の神話に関連する)(17章)。しかし、もし欠乏神話が真実だとしたら、20億人以上が飢えに苦しんでいるのに、どうして100億人以上を養うだけの食糧を生産できるのだろうか(Holt-Giménez et al.、2012)?
これらの神話や物語は、私たちが直面している複数の危機が相互に深く関連しているという事実を完全に見落としている。これらの相互関係は、歴史的、倫理的、経済的、社会的、文化的、政治的、構造的な要因とともに、システム全体の有意義な変革を可能にするために、よりよく理解される必要がある。しかし、資本主義経済システムは、食料・農業システムおよびこれらの相互関連する多くの危機の原動力のひとつですが、このシステム自体、一連の信念、価値観、倫理観に基づくより広範なパラダイムや世界観を反映している。このパラダイムや世界観は、科学、農業、知識システムなど、私たちのシステムの多くを形作っている(第1章、第2章、第9章、第16章)。したがって、私たちが直面している課題は、より深い根源にあると考える。それは、私たちが自然(人間のいない環境だけでなく、生命の網の目として理解される)から、ひいては私たち自身とお互いを分離し、切り離し、支配することに根ざしている。この断絶があるからこそ、私たちは他者の権利を無視し、抑圧し、搾取し、侵害し、自然を破壊することができるのである(第2章、第6章)。私たちの考えでは、抑圧と暴力の構造による人間や他の動物、自然に対するこのような膨大なレベルの不正と苦しみに対処することは、究極的には価値と倫理の問題である。責任ある食糧と農業のシステムは、利益によって、あるいは選択的な科学に基づいて駆動し、維持することはできない。倫理、公平性、生活の質、そして地域や国際的につながりのある市民社会の情報提供によって形成されなければならないのである。つまり、食料・農業システム、それが組み込まれた経済システム、そして私たちが活動しているパラダイム全体を支える倫理観、世界観、信念のあらゆるレベル、あらゆる領域での変革が必要なのである。
本書の範囲
現在、私たちが直面している生態系、社会、経済的な脅威や課題の多くを牽引しているのが食料・農業システムであることを踏まえ、本書は、現在の食料・農業システムとそれが機能している狭いパラダイムを「再考」(見直し、再評価、再想像を含む)する試みである。
この再考には、持続不可能性、農業環境の悪化、自然破壊、科学と知識システムの欠点、不平等、飢餓と食糧不安、不調和の歴史的、倫理的、経済的、社会的、文化的、政治的、構造的要因や根本原因を探り、明らかにすることが必要である。したがって、この再考は、食料安全保障と生産を含む「持続可能な開発」に向けた過去と現在の取り組みを見直し、これらの取り組みが、適切な文化的責任、許容できる社会的・環境的コスト、システム全体を通じて万人のための持続可能性、公平性、正義を確保する最適な関与をもって実施されているか、またはされているのかを再評価することも必要となる。
この再考は、最終的には、食糧と農業のシステムと、新しいシステムを共同創造するために必要な努力を再考することを要求する。そのためには、農民とそのコミュニティ、市民社会グループ、社会運動、開発専門家、科学者、その他の関係者が、これらの問題に対する認識を高め、解決策を実行し、例えば、より持続可能で公正な農業、天然資源管理、人間の栄養の代替パラダイムに向けて「新しい道」を切り開いてきた多くの方法を強調する必要がある。
本書は、この分野で最も経験豊かで先見性のある学者、活動家、開発専門家、実務家を集め、現在の食糧・農業システム、およびその基盤となっている科学と政治経済を厳密に問い直し、「再考」するものである。この再考は、食料・農業システムが「包括的責任」の概念によってどのように導かれるのか、またそのようなシステムはどのようなものなのかを展望するための土台となる(第20章)。包括的責任に基づく食糧・農業システムは、惑星境界に基づく全体的な生態系の持続可能性の不可欠な部分として、農業生態学的持続可能性に焦点を当てるよう社会に促すものである。このようなシステムは、すべての人にとっての生活の質、多元性、公平性、正義を重要視するものである。農家、消費者、その他すべての利害関係者、そして人間以外の動物や自然界の健康、幸福、主権、尊厳、権利を重視することになる。「包括的責任」の概念は、究極的には、自然の相互関連性とその中での人間社会の位置づけと責任についての理解に基づくものである。
書籍の構成
各章は、食料・農業システムの1つまたは複数の側面を批判的に考察し、新たな前進の道を明らかにする。いくつかの章は、食料・農業システムの発展方向と関連する上記の課題という観点から、人類社会と制度の過去と現在の行動とその結果を理解することに重きを置いている。また、他の章では、新しい、あるいは代替的な前進の方法、それらが何を提供し、どのように達成されうるかを強調することに重点を置いている。
第1章から第3章の焦点は、現在の食糧・農業システムを支える重要な歴史的・哲学的ルーツ、世界観、イデオロギーを明らかにすることにある。先に述べたように、これらのルーツを理解することなしに、なぜ、どのようにしてこのような状況に陥っているのかを真に理解することはできず、また、真の意味での効果的な解決策を開発することもできない。第1章では、農業システムを支える従来の科学的パラダイムの根底にある信念と世界観について考察している。自然に対する唯物論的な哲学に基づく現在のパラダイムと、自然を有機的で生きているものとして捉えるより全体的なパラダイムが、農業の研究開発に与える影響を探っている。第2章では、自然を支配する人間中心的な世界観の歴史的ルーツを探る。第2章では、私たちがいつ、どのようにして自然から切り離されたのか、そしてその切り離しが人間や他の動物、そして地球に及ぼすさまざまな影響を明らかにする。第3章では、19世紀半ばから続く国際的な食糧体制という観点から、世界の食糧・農業システムの政治経済について分析する。植民地主義や資本主義などの歴史的条件が、現在のグローバルな食糧・農業システムの危機の根底にあり、気候変動と生態系の複合的な緊急事態に貢献していることを述べている。
第4章と第5章では、現在の食糧・農業システムと、それがもたらす気候・生態系・食糧安全保障の危機の根源を探り、主流の開発機関が提供する物語と解決策の例を批判的に検討する。これらの物語は、しばしば無批判に受け入れられ、企業部門が継続的な蓄積と拡大を正当化するために利用されている。第4章では、新植民地主義のレンズを通して、「食料安全保障と栄養のための新同盟」の例と、より広範なSDGsを分析する。また、援助を通じて促進・維持される新植民地主義的な関係が女性に与える影響について、ジェンダーの視点から取り上げる。第5章では、国連食糧農業機関やその他の国際機関のモデルによって予測されてきた、潜在的な食糧危機と「100億人を養う」という課題についての物語を批判的に検証している。この予測や危機の物語は、企業のアグリビジネスや主流の開発組織によって、遺伝子組み換え作物の開発・使用を含む工業的農業の拡大・強化を正当化するためにしばしば利用されている。
第6章では、「包括的責任」が自然の相互関連性、そしてすべての人にとって持続可能で公正な食料・農業システムを重視していることから、倫理の問題に焦点を当てる。環境倫理、食品倫理、動物倫理の分野における重要な概念や議論を検討し、人間中心主義的な倫理観や世界観に挑戦している。これは、この後の多くの章に倫理的な枠組みを提供するものである。
第7章と第8章では、食と農のシステムの概要を説明している。このシステム、特に工業的農業が地球上に与えた影響、そして現在も与え続けている影響の大きさとその方法について、理解を深めるのに役立つ。また、このシステムが環境、健康、生活の質に与える影響と、それらの関連性を探る。両者とも、食糧と農業のシステムを変革するための前進の道を示唆している。第7章は、公共政策、民間企業、そして消費者が進むべき道を示唆している。第8章では、食品と農業のシステムが、気候変動を抑制し、人間、動物、地球の全般的な健康を改善するための解決策を提供できるかを概説している。
これらの食料・農業システムの概要に続き、第9章から第12章では、特に農業の生産システムに焦点を当てている。第9章では、科学の2つのパラダイム(第1章で検討)に基づき、工業的農業とアグロエコロジー的農業という2つの農業モデルを提示している。第10章では、現在主流となっている産業的なグリーン革命農業のパラダイムの歴史と影響を含め、農業のさまざまなパラダイムを検証している。また、有機農業、アグロエコロジー、再生農業、保全農業など、支配的なパラダイムと並行して、あるいはそれに対抗して登場・発展してきた主要な代替パラダイムの生態学的持続性を検討・分析する。そして、これらの代替パラダイムが、包括的な責任ある食料・農業システムの中で、生態系の持続可能性を高めるための方法を提案している。第11章では、従来の農業による土壌劣化という大きな問題に焦点を当てる。歴史上、社会を悩ませてきたこの問題を解決し、持続可能な食糧生産のために土壌の健全性を高めることに貢献する、保全農業という代替パラダイムの可能性を探っている。第12章では、従来の農業が温室効果ガスの排出源であり、気候変動に対して脆弱であるという問題を取り上げている。この章では、保全農業という代替パラダイムが、気候変動への適応と緩和を可能にする様々な方法を検証している。
第13章と第14章では、大手アグリビジネスの遺伝子組み換えや遺伝子編集による解決策に代わる、農家を中心とした種子システムと農業の生物多様性について検討し、今後の方法を提案している。第13章では、遺伝子組み換え作物の持続可能性への影響を評価する。本章では、除草剤耐性およびバチルス・チューリンゲンシス殺虫性GM作物の持続可能性への影響をまとめ、GM作物開発の概要を説明し、商業GM作物における意図しない形質の問題に焦点を当て、ゴールデンライスのケーススタディで説明している。また、本章では、植物育種家が持続可能な農業をどのように支援・促進し、ひいては持続可能な食糧システムの構築に貢献できるかを論じている。
第14章では、農業生産に重要な生物多様性の一部である「農業生物多様性」の歴史、範囲、機能、およびガバナンスについて考察している。
本章では、農業生物多様性、特に種子の管理に関する競合するパラダイムと、その多様性に対する脅威を説明する。この章では、公平で持続可能かつ責任ある食料システムに不可欠な、弾力的な農業生態学的生産システムを支える異種の種子を持続させるための社会的・環境的要請を強調している。
第 15 章から第 18 章では、農業生産システムの枠を超え、包括的で責任ある食料・農業システムの推進役および支援役となる他の重要な分野での新たな前進の道を探ることにする。健康的で持続可能な食生活、知識システム、社会運動、政府政策などである。第 15 章では、責任ある食糧システムを導く上で、健康的な食生活が果たす重要な役割に焦点を当てている。健康や病気に及ぼす食事の影響に関するデータを検討し、食事と人類や地球の健康との関係を探り、人類や地球の健康を最適化する食事パターンを提案する。第16章では、包括的な責任ある食と農のシステムを支えるために、私たちが採用すべき知識システム、態度、行動の種類を探求している。この章では、知識システムがしばしば、強力な利害関係者や埋め込まれたニュートン的な専門的パラダイムに立ち向かい、変革しなければならないことを明らかにしている。また、農家の複雑で多様な状況を学ぶための「厳密性の規範」を提案し、地域と支配的な知識システムの間の新しいバランスと最適性を主張している。第17章では、食料・農業システムの抜本的な変革における草の根社会運動の役割を検証している。企業、政府、慈善事業者が推進する解決策は現状維持に陥りがちであるため、この変革は草の根からもたらされなければならないと論じている。第18章では、現在の食糧システムが、「自由貿易」協定、補助金、大規模かつグローバルなものに偏った税制によって、いかにグローバル化によって生み出されてきたかを考察している。そして、愛とつながりの欲求を反映した豊かな地域経済を築いている人々の新しいあり方を紹介し、グローバリゼーションに対するシステム的な解決策と変革へのステップを提案している。
第19章では、「生きる道」と「死ぬ道」という2つの道の本質を提示する。生命の道とは、自然との共創、多様性、還元、地球の恵みの共有という原則に基づくものである。一方、死の道とは、化石燃料と毒物に基づく工業的な道であり、地球とその生物多様性に対する戦争である。
結論の章では、前章までの主要なテーマと進むべき道を抽出している。これらのテーマをもとに、食料と農業に関する「包括的責任」の概念をさらに詳しく説明し、包括的責任のある食料と農業システムの可能なビジョンを提示している。
私たちは、本書を包括的なものとし、食料・農業システムのあらゆる側面を深くカバーしたいのだが、それはあまりにも広大な領域であるため、不可能だった。そこで、本書では、土地に根ざした食糧と農業の生産に限定している。その範囲内で、私たちが埋めたいと思った明らかなギャップは、食品廃棄物の原因と対処法、再野生化などの生態学的修復、土地正義などの構造的解決策、食料・農業システムを変革し貧困と飢餓を緩和するための基本、脱成長戦略への移行、畜産業の社会・生態的影響に関するものだった。その他、もっと深く掘り下げたいと思った分野もある。食料・農業システムの資本主義的性質、その植民地的ルーツ、そしてそれが位置する広範な新自由主義的企業資本主義経済システムの検証。これらの世界的危機を助長し対処する上での国際開発および関連機関(国連およびその機関、世界銀行および地方銀行、国際通貨基金、CGIARシステム、二国間および多国間援助コミュニティ、企業・非企業双方の財団、「慈善資本家」を含むなど)の役割について。社会組織や生産者組織、ラ・ヴィア・カンペシーナや絶滅の反乱などの社会運動組織、そして、食料・農業システムに対する企業の買収に抵抗し、変革的な構造改革を要求するために必要な社会的・政治的パワーを構築する市民社会の役割、食料主権の支援と維持における地元企業の役割、持続性と進歩のための環境づくりにおける公共・民間部門の制度や政策の役割。また、実験室で栽培された食品の将来性と結果を評価する章を設けることも適切であっただろう。最後に、本書は、グローバル・サウスからの寄稿を増やすことで大きな利益を得ることができただろう。
農業のイノベーションを自由にする
ルパート・シェルドレイク(Rupert Sheldrake)
英国デヴォン州ダーティントンにあるシューマッハ・カレッジ。
1.1 はじめに
19世紀後半以降、地球はテクノロジーや現代医学による科学の応用によって変貌を遂げた。また、集約的な農業や工場での耕作によって、食糧生産が大幅に増加した。同じ時期に、資本主義は、貿易、投資、グローバル企業など、利益を上げることを仕事とする国際的なシステムを作り上げた。すべての人が影響を受けているのである。現在、ほとんどすべての農業システムは、このような科学技術や資本主義の環境の中で運営されている。世界のほとんどの地域で、伝統的な農法は、機械化や工場で作られた肥料、除草剤、殺菌剤、殺虫剤の使用など、現代の「科学的」手法に取って代わられた。灌漑は広く普及し、再生不可能な帯水層を犠牲にしていることも多く、地域によっては灌漑が塩害によって土壌を劣化させている。農地を増やすための大規模な森林伐採は、生物多様性を驚くほど失わせる要因となっている。1800年の10億人から1927年の20億人、2019年の77億人へと人口が増加したことで、食料に対する需要が大幅に増えた。また、肉食が増えたことで、穀物や大豆などの作物を使って工場で何十億もの動物を養う必要が生まれた。
科学と経済学は理論的に中立ではない。世界観の表現であり、一般的な世界観を意識しなければ、盲信してそれに従うことになる。
科学史家のトーマス・クーンは、その影響力のある著書『科学革命の構造』(1962)の中で、科学はいつでも、パラダイムと呼ばれる現実の特定のモデルによって形作られていると主張した。パラダイムは、研究を行うための有効な方法を定義する。しかし、時折、科学革命が起こり、より包括的な新しいパラダイムが古いパラダイムに取って代わることで、現実の基本的なモデルが変化することがある。パラダイムの変化を促す要因として、既存のパラダイムに当てはまらない厄介な現象である「アノマリー(異常)」がある。パラダイムが変化し、現実をより広く見ることができるようになり、これらの異常が含まれるようになるまで、通常、これらの異常は却下、否定、または説明されることが多い。
現代の科学と経済は、すべての現実は物質的または物理的であるとする唯物論的な自然哲学に基づいている。物質的な現実以外には現実は存在しない。物質は非意識的である。人間の意識は、脳の物理的活動の機能しない副産物である。神は人間の心の中の観念としてのみ存在し、したがって人間の頭の中にも存在する。自然界には目的がなく、進化も無目的である。この唯物論的パラダイムにとって、一つの大きな異常は意識そのものであり、その存在そのものが、心の哲学において「難問」と呼ばれている。もう一つは「目的」である。もちろん、人間にも目的はあるし、人間以外の動物にも目的はある。では、自然が基本的に無意識で無目的であるならば、意識や目的はどこから来るのだろうか。哲学者の中には、このような疑問と格闘している人もいるが、ほとんどの人は単に無視するだけだ。
このような唯物論的信念が強力なのは、ほとんどの人が批判的に考えるからではなく、考えないからなのである。科学の事実は十分に現実的であり、科学者が使う技術も、それに基づく技術もそうである。しかし、従来の科学的思考を支配する信念体系は、19世紀のイデオロギーに根ざした信仰行為である。私たちの経済システムの根底にも、同じように唯物論的な前提がある。そして、これらの思想体系の根底には、機械論的な科学が確立された17世紀以来の、新しい神職に導かれた世界を変革する活動としての科学というビジョンがある。科学と技術の巨大な成功は、確かに世界を変革し、このビジョンが真実であることを証明したように思われる。しかし、それは同時に、無批判な科学的教条主義をもたらし、自由な探求を阻害し、無数の種の生存を脅かし、人類の生存を危うくしている。
1.2 科学の神権
フランシス・ベーコン(1561-1626)は、政治家であり弁護士であり、イングランド大法官となったが、誰よりも組織科学の力を予見していた。そのためには、自然を支配する力を手に入れることが、何ら不吉なことではないことを示す必要があった。彼が執筆していた当時は、魔術や黒魔術に対する恐怖が蔓延していた。彼は、自然の知識は神が与えたものであり、悪魔に感化されたものではないと主張し、それに対抗しようとした。科学とは、堕落する前のエデンの園にいた最初の人間、アダムの無垢な姿に戻ることだった。
ベーコンは、聖書の最初の書である「創世記」が科学的知識を正当化すると主張した。彼は、人間が自然を知ることを、アダムが動物に名前をつけたことと同一視した。神は「アダムが何を呼ぶかを見るために、彼らをアダムに引き合わせた。アダムがすべての生き物を呼んだものは、その名前であった」(創世記2:19-20)。エバが創造されたのはその2節後なので、これは文字通り人間の知識であった。ベーコンは、人間が自然を技術的に支配することは、新しいことではなく、神から与えられた力の回復であると主張した。彼は、人々が新しい知識を賢く、うまく使うことを確信した。「ただ、人類が、神の遺贈によって人類に与えられた自然に対する権利を回復させるだけである。
この自然に対する新しい力の鍵は、組織的、組織的な研究であった。ベーコンは『ニュー・アトランティス』(1624)の中で、科学的聖職者が国家全体の利益のために意思決定を行う技術主義的ユートピアを描いている。この科学的な「教団または協会」のフェローたちは、長いローブを身にまとい、その権力と尊厳が必要とする敬意をもって扱われた。教団のトップは、輝く黄金の太陽像の下、豪華な馬車で移動した。彼は行列に乗りながら、「民衆を祝福するように、素手を上げて進んだ」
この財団の一般的な目的は、「物事の原因や秘密の運動を知り、人間の帝国を拡大し、あらゆることを可能にすること”だった。学会には、爆発物や軍備を試験するための機械や施設、実験炉、植物育成のための庭園、調剤薬局などが備えられていた。
1660年にロンドンで設立された王立協会をはじめ、多くの国の科学アカデミーや研究所が、この先見性のある科学機関によって研究されるようになったのである。しかし、これらのアカデミーのメンバーはしばしば高く評価されたものの、ベーコンが想像した原型のような壮大さと政治的権力を持つものはなかった。
ベーコンの時代のイギリスでは(現在でも)、イギリス国教会が既成の教会として国家と結びついていた。ベーコンは、科学者の神職もまた、国家の庇護を受けることで国家と結びつき、一種の確立された科学の教会を形成することを想定していた。そして、ここでもまた、彼は予言的であった。資本主義国家でも共産主義国家でも、公的な科学アカデミーは科学界の権力中枢であり続けている。科学と国家の分離はない。科学者たちは、戦争、産業、農業、医学、教育、研究などに関する政府の政策に影響を与える、確立された神権のような役割を担っている。
ベーコンは、政府や投資家から資金援助を得るための理想的なスローガンを作り上げた。「しかし、科学者が政府から資金を引き出すことに成功するかどうかは、国によって異なる。19世紀後半まで、ほとんどの研究は私費で行われるか、チャールズ・ダーウィンのような裕福なアマチュアが行っていたのである(Kealey, 1996)。
フランスでは、ルイ・パスツール(1822-95)が、科学は真理を探究する宗教であり、研究所は人類が最高の潜在能力を発揮するための神殿のようなものであるとして、有力な提唱者となった。
私たちが実験室という表現で呼んでいる神聖な施設に関心を持ってみよう、お願いしたい。彼らは富と未来の神殿である。そこでは、人類が成長し、より強く、より良くなるのである。
20世紀初頭までに、科学はほぼ完全に制度化・専門化され、第二次世界大戦後は政府の庇護のもと、また企業の投資によって膨大な規模に拡大した。e 最も資金が豊富なのは米国で、2015年の研究開発費総額は4950億ドル、うち1210億ドルが政府からのものだった。f しかし政府や企業は通常、科学者に研究費を支払うのは、堕落以前のアダムのような無垢な知識が欲しいからではない。熱帯雨林の絶滅危惧種である甲虫を分類するように、動物に名前をつけることは優先順位が低いのである。ほとんどの資金援助は、ベーコンの説得力のあるスローガン「知識は力なり」に応えるものである。
1950年代、組織的な科学がかつてないほどの権力と名声に達したとき、科学史家のジョージ・サートンは、まるで宗教改革前のローマ・カトリック教会のような言い方で、この状況を肯定的に表現している。
真理は専門家の判断によってのみ決定される…。
すべてのことは、非常に小さなグループ、実際には、一人の専門家によって決定され、その結果は慎重にチェックされるが、他の数人の専門家によってチェックされる。国民は何も言うことができず、ただ渡された決定事項を受け入れるだけである。科学的活動は、大学、アカデミー、科学協会によってコントロールされているが、このようなコントロールは、民衆のコントロールから可能な限りかけ離れている。
トーマス・クーンは、科学の社会的側面に注目し、科学が集団的な活動であることを思い出させるのに貢献した。科学者は、仲間集団の圧力や集団の規範に従う必要性など、人間の社会生活における通常の制約をすべて受けている。クーンの議論は主に科学史に基づくものであったが、科学社会学者は、科学者が支援のネットワークを構築し、資源や結果を利用して権力や影響力を高め、資金や名声、認知度を競う方法を研究し、実際に実践されている科学を研究することによって、彼の洞察をさらに深めた。ブルーノ・ラトゥールの『行動する科学』。1987)は、この伝統的な研究において最も影響力のある研究の一つである。
ラトゥールは、科学者が日常的に知識と信念を区別していることを観察した。専門家グループ内の科学者は、自分たちの科学分野がカバーする現象について知っているが、ネットワークの外の人たちは歪んだ信念しか持っていない。科学者は、自分たちの集団の外にいる人々について考えるとき、どうして彼らはまだそんなに不合理なことができるのだろうかとよく考える。
科学者が描く非科学者の姿は暗澹たるものになる。少数の頭脳が現実とは何かを発見する一方で、大多数の人々は不合理な考えを持ち、少なくとも、時代遅れの偏見に頑なに固執する多くの社会的、文化的、心理的要因の囚人である。この図式の唯一の救いは、人々を偏見の虜にしているこれらの要因をすべて取り除くことが可能であるならば、彼らは皆、直ちに、何のコストもかけずに、科学者のように健全な精神状態になり、現象を理解できるようになるということである。私たち一人ひとりの中に、眠っている科学者がいて、社会的・文化的な条件が整わない限り、目覚めることはないのである。
「科学的世界観」の信奉者にとっては、教育やメディアを通じて科学に対する一般の理解を深めればよいのである。
19世紀以来、唯物論は実に見事に広まり、何百万人もの人々が、科学そのものについてほとんど知らないにもかかわらず、この「科学的」見解に改宗した。彼らはいわば、科学者たちが司祭を務める科学教会、あるいは科学主義の信奉者なのである。無神論者の著名な一般人であるリッキー・ジャーヴェイスは、『タイム』誌の「世界で最も影響力のある100人」に選ばれた2010年に、『ウォールストリート・ジャーナル』でこのような姿勢を示している。Gervaisはエンターテイナーであり、科学者でも独創的な思想家でもないが、自分の世界観を支持するために科学の権威を借りている。
科学は真実を追求する。科学は真実を追求するものであり、差別はしない。科学は真理を探究し、差別はしない。良くも悪くも、物事を発見する。科学は謙虚である。科学は謙虚であり、自分が知っていることも、知らないことも知っている。結論と信念は、確固たる証拠、つまり常に更新され、アップグレードされる証拠に基づいている。新しい事実が出てきても、怒ったりはしない。知識の体系を受け入れるのである。伝統だからといって、中世の慣習にしがみつくこともない。
ジャーヴェイスの理想化された科学観は、科学の歴史や社会学の文脈からすると、絶望的にナイーブなものである。科学者は真理を求めるオープンマインドな人であって、資金や名声を競い、仲間内の圧力に縛られ、偏見やタブーに閉ざされた普通の人ではない。
科学者の聖職者というベーコンの構想は、今や世界的な規模で実現されている。しかし、自然に対する人間の力が「健全な理性と真の宗教」によって導かれるという彼の自信は見当違いであった。
1.3 全知全能というファンタジー
全知全能という幻想は、科学者たちが神のような知の総体を目指す、科学史の中で繰り返し登場するテーマである。19世紀初頭、フランスの物理学者ピエール・シモン・ラプラスは、すべてを知り、予測することのできる科学者の心を想像していたi。ダーウィンの進化論を広めたトーマス・ヘンリー・ハクスリーは、機械的決定論を進化の過程全体に拡大解釈した。
もし進化の基本的命題が真実で、生きているものも生きていないものも含めた全世界が、宇宙の原始的な星雲が構成された分子が持つ力が、明確な法則に従って相互に作用した結果であるとするならば、現存の世界が潜在的に宇宙の蒸気の中にあったことは間違いなく、その蒸気の分子の特性に関する知識から、十分な知性が、例えば1869年のイギリスの動物相を予測できたかもしれない。
決定論の信念が人間の脳の活動に適用されたとき、脳の分子的、物理的活動に関するすべてが原理的に予測可能であるという理由で、自由意志が否定される結果となった。しかし、この確信は科学的な証拠に基づくものではなく、すべてが数学的な法則によって完全に決定されているという仮定に基づくものであった。現在でも、多くの科学者が「自由意志は幻想である」と考えている。脳の活動が機械のようなプロセスで決定されるだけでなく、選択を行うことのできる機械的でない自己は存在しないのである(Chivers, 2010)。
1927年、量子物理学における不確定性原理の認識により、不確定性は物理世界の本質的な特徴であり、物理的な予測は確率の観点からしか行えないことが明らかとなった。その理由は、量子現象は波動であり、波動はその性質上、空間と時間に広がっているため、ある瞬間にある一点に局在することができず、より厳密に言えば、その位置と運動量はともに正確に知ることができないからだ。量子論は統計的確率を扱うものであり、確実なものではない。量子事象において、ある可能性が他の可能性よりも実現されるという事実は、偶然の問題なのである。ネオ・ダーウィンの進化論では、量子事象である遺伝子の偶然の突然変異を通じて、ランダム性が中心的な役割を担っている。偶然の出来事が違えば、進化の仕方も違ってくる。T・H・ハクスリーは、進化の過程が予測可能であると信じたのは間違いだった「進化生物学者のスティーブン・ジェイ・グールドは、「生命のテープを再生すれば、異なる生存者たちが今日の地球を飾るだろう」と言った(グールド、1989)。
20世紀に入ると、量子過程だけでなく、液体の乱流や海辺の波打ち際、天気など、ほとんどすべての自然現象が確率的であることが明らかになり、正確な予測を超えた自発性と不確定性を示すようになった。気象予報士は、強力なコンピューターと人工衛星からの絶え間ないデータにもかかわらず、いまだに失敗を繰り返している。これは、彼らが悪い科学者だからではなく、気象は本質的に詳細が予測できないからだ。秩序がないという日常的な意味ではなく、正確に予測できないという意味で、カオスなのだ。カオス理論」と呼ばれるカオス力学の観点から、気象を数学的にモデル化することはある程度可能だが、これらのモデルは正確な予測をすることはできない(Gleik, 1988)。量子物理学の世界と同様に、日常世界においても確実性は達成できない。長い間、機械論的な科学の中心であると考えられてきた太陽を中心とする惑星の軌道でさえ、長い時間スケールではカオスであることが判明している。
19世紀から20世紀初頭にかけて、多くの科学者が強く信じていた決定論は、妄想であったことが判明した。科学者がこのドグマから解放されたことで、自然界の不確定性、特に進化の不確定性が新たに認識されるようになったのである。科学は、決定論という信念を捨てることで終わりを迎えたわけではない。同様に、科学は、いまだ科学に縛られているドグマを失っても生き残り、新たな可能性によって再生されるであろう。
19世紀末には、科学的全知全能の幻想は、決定論への信仰をはるかに超えていた。1888年、カナダ系アメリカ人の天文学者サイモン・ニューコムは、「私たちは天文学について知り得ることの限界に近づいているのだろう」と書いた。1894年、後にノーベル物理学賞を受賞するアルバート・マイケルソンは、「物理学のより重要な基本法則と事実はすべて発見され、それらは現在、非常に強固に確立されており、新しい発見によって取って代わられる可能性は極めて低い」と宣言した。1900年、物理学者で大陸間電信の発明者であるケルビン卿ことウィリアム・トムソンは、よく引用される(アポクリファかもしれないが)主張の中で、この最高の自信を表現した。物理学で新たに発見されるものは何もない。”あとは、より正確な計測を行うだけだ。
20世紀には、量子物理学、相対性理論、原子爆弾や水爆に代表される核分裂・核融合、銀河系の発見、ビッグバン理論(約140億年前に宇宙は非常に小さく非常に高温で始まり、それ以来成長、冷却、進化を続けているという考え)などがあり、これらの確信は打ち砕かれた。しかし、20世紀末になると、20世紀の物理学の勝利と神経生物学と分子生物学の発見によって、全知全能の幻想が再び甦った。1997年、『サイエンティフィック・アメリカン』のシニア・サイエンス・ライターであるジョン・ホーガンは、『科学の終わり』という本を出版した。1997年、『サイエンティフィック・アメリカン』誌のシニア・サイエンス・ライターであるジョン・ホーガンは、『科学の終焉:科学時代の黄昏における知の限界に直面する』という本を出版した。多くの一流の科学者にインタビューした後、彼は挑発的な論文を発表した。
科学を信じるなら、科学的発見の偉大な時代が終わったという可能性、あるいはその可能性さえも受け入れなければならない。科学とは、応用科学ではなく、宇宙とその中での私たちの位置を理解しようとする人間の根源的な探求であり、最も純粋で偉大な科学という意味である。これ以上研究を続けても、偉大な発見や革命は生まれないかもしれない。しかし、その成果は漸減していくばかりだ。
ホーガンは、DNAの構造のように、一度発見されたものは、それ以上発見し続けることはできない、ということを確かに言っている。しかし、彼は従来の科学の信条が真実であることを当然と考えた。最も基本的な答えはすでに知られていると思い込んでいたのである。そして、そのどれもが、より興味深く、実りある問いに置き換えられることを、私は拙著『科学の妄想』(米国では『Science Set Free』)の中で明らかにしている。
1.4 唯物論の信頼性の危機
生物学の分野では、1970年代から1980年代にかけて、極端な唯物論が分子生物学という形で定着し、やがてそれが主流となった。よりホリスティックな生物学は疎外されたのである。この分子生物学のパラダイムは、医学や農業に大きな影響を与え、研究の中心を分子レベルに移した。ゲノムプロジェクト、動植物の遺伝子組み換え、遺伝子編集技術、その他の独創的なバイオテクノロジーに、何千億ドルもの公的・民間資金が注ぎ込まれた。ヒトゲノムや他の多くの生物種のゲノム配列決定という技術的勝利のような印象的な成功もあれば、ヒトの稀な遺伝性疾患の遺伝的基盤の同定のような特殊な応用もあった。作物の遺伝子組み換えが商業的に成功した例もいくつかある。これらの遺伝子組み換え品種の特許を所有する企業には、何十億ドルもの利益がもたらされている。しかし、このような一方的な分子生物学的アプローチは、医学や農業における他の多くの可能性から注意をそらしている。
200年以上前から、唯物論者は「科学はいずれ物理と化学ですべてを説明できるようになる」と約束してきた。科学は、生物は複雑な機械であり、心は脳の活動に過ぎず、自然は無目的であることを証明するだろう。信者は、科学的発見が自分たちの信念を正当化してくれるという信仰に支えられているのだ。科学哲学者のカール・ポパーは、このような姿勢を「約束手形的唯物論」と呼んだが、これはまだなされていない発見に対して約束手形を発行することに依存しているためである。
1963年、ケンブリッジ大学で生化学を学んでいた私は、キングス・カレッジのブレナーの部屋で、フランシス・クリックとシドニー・ブレナーとの一連のプライベートミーティングに、数人の同級生とともに招待された。クリックとブレナーは、最近、遺伝暗号の「解読」に貢献したのである。二人とも熱心な唯物論者で、クリックは過激な無神論者でもあった。彼らは、生物学には「発生」と「意識」という2つの大きな未解決問題があると説明した。この2つの問題は、分子生物学者でもなく、優秀な研究者でもなかったため、解決されなかったのである。クリックとブレナーは、10年か20年以内にその答えを見つけるつもりだった。ブレナーは発生生物学を、そしてクリックは意識を研究するのである。そして、私たちを誘ってくれたのである。
2人ともベストを尽くした。ブレナーは、小さな虫、線虫の発生に関する研究で 2002年にノーベル賞を受賞した。クリックは 2004年に亡くなる前日に、脳に関する最後の論文の原稿を添削した。葬儀の席上、息子のマイケルは、彼の心を動かしたのは、有名になりたい、裕福になりたい、人気者になりたいという欲求ではなく、「バイタリズムの棺桶に最後の釘を打ち込むこと」(Ridley, 2011)だったと語った。(バイタリズムとは、生物は物理学や化学だけでは完全に説明できないとする理論である)。
CrickとBrennerは失敗した。発生と意識の問題は未解決のままである。多くの詳細が発見され、何十ものゲノムの配列が決定され、脳スキャンはますます精度を増している。しかし、生命や心が物理や化学だけで説明できるという証明はまだない。
物質主義の基本命題は、物質が唯一の現実であるということである。したがって、意識は脳の活動にほかならない。何もしない影のようなもの、「エピフェノメノン」だろうか、脳の活動を別の言い方で表現しているに過ぎない。しかし、現代の神経科学や意識研究の研究者の間では、心の性質についてコンセンサスが得られていない。『Behavioural and Brain Sciences』や『Journal of Consciousness Studies』といった一流の雑誌には、唯物論の教義に深い問題があることを明らかにする論文が数多く掲載されている。哲学者のデビッド・チャルマーズは、主観的な体験の存在そのものを「難問」と呼んでいる。難しいのは、それがメカニズムという観点からの説明を拒むからだ。たとえ目や脳が赤い光に反応する仕組みがわかっても、赤さの体験は説明できないのである。
生物学や心理学では、唯物論の信頼性は低下している。物理学は救いの手を差し伸べることができるのか?唯物論者の中には、自分たちの希望が19世紀の物質理論ではなく、現代の物理学に依存していることを強調するために、自分たちを物理主義者と呼ぶことを好む人もいる。しかし、物理主義の信頼性は、物理学自身によって低下している。
第一に、一部の物理学者は、観測者の心を考慮しなければ量子力学は成立しないと主張している。彼らは、物理学は物理学者の心を前提としているため、心を物理学に還元することはできないと主張している(D’Espagnat, 1976)。
第二に、最も野心的な物理的現実の統一理論である超ひも理論とM理論は、それぞれ10次元と11次元を持ち、科学を全く新しい領域へと導くものである。弦理論やM理論は、現在のところ検証不可能であるため、実験ではなく、他のモデルを参照することでしか判断することができない。また、これらの理論は数え切れないほどの他の宇宙にも適用されるが、そのような宇宙は一度も観測されていない。理論物理学者のリー・スモリンは、著書『The Trouble With Physics』の中で、このアプローチ全体に深い懐疑的な見方を示している。The Rise of String Theory, the Fall of a Science and What Comes Next (2006)という本の中で、理論物理学者のリー・スモリンが示している。弦理論、M理論、そして経験的証拠によってではなく、他のモデルに対してモデルをテストする「モデル依存の実在論」は、唯物論や物理主義、その他の信念体系の土台を揺るがすものである。
第三に、21世紀に入ってから、既知の種類の物質とエネルギーが宇宙の約5%を占めるに過ぎないことが明らかになっている。残りは「ダークマター」と「ダークエネルギー」で構成されている。物理的現実の95%の性質は、文字通り不明瞭である。
第四に、「宇宙人間原理」は、ビッグバンの瞬間に自然の法則や定数が少し違っていたら、生物は誕生しなかったし、それゆえに私たちはここで考えることもなかったと主張する。では、最初に神の心が法則や定数を微調整したのだろうか?創造主である神が新たな姿で現れるのを避けるため、ほとんどの主要な宇宙論者は、M理論も示唆するように、私たちの宇宙は、異なる法則や定数を持つ、膨大でおそらく無限の並行宇宙の一つであると考えることを好んでいる。私たちは、たまたまその中の1つに存在しているに過ぎないのである。
この多元宇宙論は、オッカムの剃刀、つまり「必要以上に実体を増やしてはならない」という哲学的原則、言い換えれば「仮定をできるだけ少なくする」という原則に究極的に反するものである。また、検証不可能という大きな欠点もありますq さらに、神を排除することにも成功していない。無限の神は、無限の宇宙の神となりうるのである。
唯物論は、19世紀後半には一見シンプルでわかりやすい世界観を提供したが、21世紀の科学はそれを大きく置き去りにしてしまった。その約束は果たされず、その約束手形はハイパーインフレで切り下げられた。
私は、科学全般、特に農学が、強力なタブーに守られながら、ドグマとして固まった思い込みに阻まれていると確信している。これらの信念は、既成の科学という城壁を守る一方で、オープンマインドな思考を阻む障壁となっている。
1.5 分子生物学の果たされなかった約束
1980年代、新しい技術によって遺伝子のクローンが作られ、その遺伝暗号の「文字」の並びが発見されたときの高揚した雰囲気は、今でも思い出すことができない。生命そのものの遺伝子がついに明らかになり、生物学者が植物や動物を遺伝的に改変し、想像を超える豊かな成長を遂げる可能性が開かれたのである。「科学者ががんと闘う遺伝子を発見」「遺伝子治療が関節炎の患者に希望を与える」「科学者が老化の秘密を発見」などなど、毎週のように新聞の見出しに新しい発見が掲載された。
この新しい遺伝学は非常に有望で、すぐに生物学研究者全体がその技術を自分の専門分野に応用することに躍起になった。その結果、「ヒトゲノムの遺伝子をすべて解明する」という、壮大で野心的なビジョンが生まれた。ハーバード大学のウォルター・ギルバートは、「私たちが何者だろうかという。「聖杯」の探求は、今や頂点に達した。最終的な目標は、ゲノムの全容を把握することである”ヒトゲノム計画は、1990年に30億ドルの予算を見込んで正式に開始された。
ヒトゲノム計画は、生物学に「ビッグサイエンス」を持ち込もうとする意図的な試みだった。ミサイルや水爆、スターウォーズ、数十億ドル規模の粒子加速器、宇宙開発、ハッブル宇宙望遠鏡などに莫大な予算が投じられていた。野心的な生物学者たちは、物理学への羨望に苦しんでいた。彼らは、生物学が注目され、高い名声を得て、数十億ドル規模の国際的なプロジェクトが行われる日を夢見ていた。ヒトゲノム計画はその答えだった。
同時に、1990年代の市場投機の潮流は、バイオテクノロジーのブームをもたらし 2000年にピークに達した。公式のヒトゲノム計画に加えて、セレラ・ジェノミクス社は、クレイグ・ベンター氏を代表とする民間のゲノムプロジェクトを実施した。同社は、数百のヒト遺伝子を特許化し、その商業的権利を所有することを計画していた。セレラ・ジェノミクスの市場価値は、他の多くのバイオテクノロジー企業と同様に 2000年初頭には目もくらむような高さにまで跳ね上がった。
皮肉なことに、公的なゲノムプロジェクトと民間のゲノムプロジェクトが対立し、ゲノムの配列決定が完了する前にバブルが崩壊することになった。2000年3月、公的ゲノムプロジェクトのリーダーたちは、自分たちの情報はすべて誰でも自由に利用できるようにすることを公表した。その結果 2000年3月14日、クリントン米大統領が「私たちのゲノム、それはすべての人間の人生が書かれている本であり、人類のすべてのメンバーのものである…私たちは、ヒトゲノム研究の利益が、ドルではなく、人間の生活の向上で測れるようにしなければならない」と発言したs。報道では、大統領はゲノム特許を制限するつもりだと伝えられた。株式市場は大きく反応した。ベンターの言葉を借りれば、「病的な」s Venter (2007), p. 299.
スランプ”となった。2日のうちにセレラの評価額は60億ドル、バイオテクノロジー株の市場規模は5,000億ドルも暴落したのである。
2000年6月26日、クリントン大統領とトニー・ブレア英国首相は、クレイグ・ベンターと公式プロジェクトの責任者であるフランシス・コリンズとともに、ヒトゲノムの最初の草案を公表したことを発表した。ホワイトハウスでの記者会見でクリントン大統領は、「私たちは今日、全人類ゲノムの最初の調査が完了したことを祝うためにここにいる。間違いなく、これは人類がこれまでに作り出した中で最も重要で、最も不思議な地図である。これは、すべてではないにしても、ほとんどの人間の病気の診断、予防、治療に革命をもたらすだろう。「人類は、治癒のための計り知れない、新しい力を手に入れようとしている」英国の科学大臣セインズベリー卿は、「私たちは今、これまで医学に期待してきたことをすべて実現できる可能性がある」と述べている。[そして、人間の形を考えうる限りあらゆる形に作り上げることができるようになる。ヒトゲノムの解読という驚異的な成果は、私たちの人間観を一変させたが、それは予想されたことではなかった。まず驚いたのは、遺伝子の数があまりに少ないことである。予想されていた10万個以上の遺伝子ではなく、最終的に約23,000個という数は非常に不可解であり、私たちよりもはるかに単純な他の動物のゲノムと比較すればなおさらである。ミバエには約17,000個の遺伝子があり、ウニには約26,000個の遺伝子がある。また、植物の中には、私たちよりもはるかに多くの遺伝子を持つ種があり、例えば、イネは約38,000個も持っている。
ヒトゲノムプロジェクトをきっかけに、ムードは一変した。分子生物学者が生物の「プログラム」を知れば生命は理解できるという楽観的な考えから、遺伝子配列と実際の人間との間には大きな隔たりがあることがわかったのだ。実際、ヒトゲノムの予測値は小さいことが判明した。例えば、身長の場合、ゲノムの予測値は、メジャーで測った値よりも低いのである。背の高い親には背の高い子供が、背の低い親には背の低い子供が生まれる傾向がある。両親の身長を測れば、その子供の身長は約80%の精度で予測できる。つまり、身長は約80%の確率で遺伝するのである。2008年になると、3万人のゲノムを比較する「ゲノムワイド関連研究」が行われ、身長や低身長に関連する約50の遺伝子が特定された。驚いたことに、これらの遺伝子を合わせても、身長の遺伝に占める割合は5%程度に過ぎなかった。つまり、「身長」遺伝子は、身長の遺伝率の75%を占めていなかったのである。遺伝力のほとんどが欠落していたのである。このほかにも、多くの病気の遺伝率など、遺伝率が欠落している例が多く知られており、「パーソナルゲノム」の価値は非常に疑問視されている。2008年以降、科学文献ではこの現象は “missing heritability problem”と呼ばれている。
2011年に発表された研究では、血縁関係のない人のゲノムから予測できる身長の遺伝率の割合は15%で、以前の方法よりは改善されたが、遺伝子の配列を全く調べずに親戚の身長を測定して予測できる遺伝率には、まだ遠くおよびませんだった。また、巻尺で身長を測るのは、ゲノム解析よりも何十億円も安い。
分子技術から大きな利益が得られると期待した株式市場の投資家たちの楽観論は、何度も打撃を受けた。2000年のバイオテクノロジー・バブル崩壊後、多くのバイオテクノロジー企業が倒産するか、製薬会社や化学会社に買収された。2004年のウォール・ストリート・ジャーナル紙には、「バイオテックの悲惨な底辺」と題する記事が掲載されている。バイオテクノロジーが経済成長の原動力となり、致命的な病気を治すことができるかもしれない。しかし、それが良い投資であることを主張するのは難しい。2006年、ハーバード・ビジネス・スクールは、バイオテクノロジー産業の詳細な分析を発表した。2006年、ハーバード・ビジネス・スクールは業界の詳細な分析を発表し、利益を上げたバイオテクノロジー企業は「ごく一部」であり、ブレイクスルー発明を約束しても何度も失敗していることを明らかにした。バイオテクノロジー業界を擁護する人々は、もっと時間が必要だと主張したが、ハーバード・ビジネス・スクールの分析は逆の結論を示している。「バイオテクノロジー業界全体、そして特定の企業の長期的な業績が極めて芳しくないことを考えると、資本はどちらかといえば忍耐強すぎたのである」
それにもかかわらず、新しいバイオテクノロジー企業は絶えず立ち上げられている。その多くは、何億ドルもの資金を投じて、何の成果も得られないまま終わっている。2015年頃から、CRISPR-Cas9と呼ばれる新しい遺伝子編集技術が、バイオテクノロジーの新興企業の新しい波の基盤として人気を博している。これらの技術は、特に希少な遺伝子疾患に対する特殊な医療用途や、植物育種におけるニッチな用途につながる可能性が十分にある(Jaganathan, Ramasamy, Sellamuthu, Jayabalan, & Venkataraman, 2018)。プライム編集」と呼ばれる新しい遺伝子編集技術では、DNA配列のさらに精密な操作が可能である(Cohen, 2019)。しかし、非常に多くの形質が多遺伝子であるため、精密な遺伝子編集は非常に限られた用途しかない。それらは、精密に編集できる1つか2つのマスター遺伝子ではなく、非常に小さな影響を持つ多数の遺伝子によって影響を受ける。
分子生物学とバイオテクノロジーへの膨大な投資は、その事業実績が不十分であったにもかかわらず、多くの雇用を創出することによって、生物学の実践に広範な影響を及ぼしている。分子生物学を専攻する卒業生への需要は、生物学の教育を一変させた。現在、ほとんどの大学で分子生物学的アプローチが主流となっており、中等教育での科学教育にも強い影響を及ぼしている。しかし、分子生物学がこれほどまでに重視されるようになったからこそ、その限界がますます明らかになりつつある。動物や植物の種が増え続け、ゲノムが解読され、何千ものタンパク質の構造が明らかになったことで、分子生物学者は自分たちのデータに溺れそうになっている。ゲノムの配列やタンパク質の解析には限界があるのだ。分子生物学者は現在、急成長しているバイオインフォマティクス分野のコンピュータ専門家に、「データ雪崩」とも呼ばれるこの前例のない量の情報を保存し、意味を理解しようと頼っている(Howe & Rhee, 2008)。
1.6 より総合的なアプローチに向けて
こうした歴史的展開や哲学的動きは、資本主義の成長、特にグローバル化した新自由主義的形態とともに、農業に多大な影響を及ぼしてきた。これらの前提を認識することで、農業の研究がより全体的な方向へ向かうことが容易に理解できるようになる。
機械論的な唯物論のパラダイムは、今、自然を有機的で生きているものとしてとらえる、よりホリスティックなパラダイムに挑戦している。一部の心の哲学者や神経科学者は現在、代わりに、原子を含むすべての自己組織化システムが心や主観的経験の要素を持つとする汎心論的世界観を採用している(Goff, 2019)。地球を生命体として捉える「ガイア仮説」は、この変化の一例である。しかし、こうした理論的な議論に興味のない人にとっても、現実的な理由だけでも、私たちは考え方や行動の仕方を変える必要があるのである。
- 1. 科学聖職者崇拝は、大学の科学者、特に先進国の大学の科学者の知識を、地元の農家や農法よりも優遇する。伝統的な知識体系が迷信や無知として否定され、技術専門家の現在流行している見解が優先される。農業に対するより総合的なアプローチは、伝統的な知識、地元に適応した作物品種、実践、経験を無視するのではなく、それらについてより深く知り、適切な場合には農業開発政策に統合しようとするものである。
- 2. 全知全能の幻想は、自然界に存在する複雑な生態系や相互関係のパターンを無視し、定量的な正確さに執着することにつながる。このことは、モノカルチャーの独占に反映され、また、改良された品種は、農家が事前に栽培していた何千もの地域に適応した品種とは異なり、科学的に改良された新しい作物であるため、広い地域に展開することができるという考えにつながる。植物育種では、遺伝子組み換えが可能な少数の遺伝子に焦点を当て、CRISPR-Cas9やプライムエディットでは、単一遺伝子や単一遺伝子の小さな部分に焦点を当てることになるのである。また、マメ科植物の根粒で窒素を固定する根粒菌のように共生が認められる場合、土壌中には根粒菌と共生菌根粒菌が複雑に混在しているにもかかわらず、単一の決まった菌株の植菌が行われている。定量的な精度にこだわるのではなく、混作や土壌微生物群の重要性、さまざまな遺伝子の重要性、遺伝におけるエピジェネティックな影響などを認識することが必要である。
- 3. 経済学や政府計画では、利益や国民総生産の成長といった定量的な結果にこだわるあまり、農民の社会的・文化的生活、土地所有や労働に関する倫理的問題、動物福祉、生態系の健康、汚染の影響、土壌侵食や劣化による長期的な損害などを無視したトンネルビジョンに陥ってしまう。より全体的なアプローチでは、これらの要素を無視するのではなく、考慮することになる。
- 4. 経済的な測定は、国民総生産の年間成長率、企業の年間利益、作物の年間収穫量の数字のように、通常、短期的に行われるものである。企業や株主も毎年の利益で考え、民主主義国家では政治家も数年の選挙サイクルで考える。しかし、地球の生態系、土壌の健康、農法の持続可能性などは、もっと長期的な思考が必要である。
- 5. ビジネスのグローバル化が進み、巨大な力を持つ企業は、広告を通じて消費者の需要を、ロビー活動を通じて政府の政策を左右することができるようになった。彼らの動機は、人間の福祉や健康、生態系の長期的な持続可能性、健全な人口ではなく、短期的な利益である。その結果、土壌の劣化、多くの種の絶滅、有毒廃棄物、農場からの流出物、肥満の蔓延などが起こっている。よりホリスティックなアプローチでは、必然的に長期的かつ持続可能な視点が必要となる。
これらの原則に照らして、農業研究開発の新たな可能性についていくつか考察する。土壌劣化、環境汚染、森林破壊、生物多様性の喪失、人口増加、気候変動、限られた水資源、そして肥満の蔓延という状況の中で、従来通りのビジネスは持続不可能である。大小さまざまな規模での新しい取り組みが必要である。ここでは、いくつかの可能性を紹介する。土壌のマイクロバイオームの強化、実用的な植物育種、エピジェネティックな遺伝など、世界観の変化に依存するものもあれば、唯物論と非物質論の両方の思考の枠組みの中で意味を持つものもある。
1.6.1 伝統的な慣習の再発見と検証
世界中の農家には、機械論的な科学が登場するずっと以前から発展してきた伝統的な慣習がある。その中には混作(間作を含む)のように、思想的な観点から比較的議論の余地のないものもある。しかし、機械化の結果、混作などの伝統的な農法がモノカルチャーに取って代わられることが多くなっている。しかし、先進国でも牧草地では、牧草とクローバーの混合栽培が行われており、クローバーの根粒による窒素固定によって窒素肥料の必要性を減らしている。メキシコの「3姉妹」のように、トウモロコシ、インゲンマメ、カボチャを群生させるなど、マメ科の作物と穀物を組み合わせる伝統的な混作・間作システムも同じ原理である。インタークロッピングシステムは、例えばインドのソルガムとキマメのように、穀類とマメ科作物を交互に畝に植えていく、よりシステム化されたシステムである。このシステムでは、キマメは土壌を窒素で豊かにし、根の残渣や落ち葉を通じて後続作物に恩恵を与えるだけでなく、早生ソルガムの収穫後も空いたスペースを利用して成長を続け、モンスーンの終了後もその深い根を通じて残留土壌水分を抽出する(Sheldrake、1984)。このような間作システムは、土地や資源の利用効率が高く、さまざまな環境条件下で高い収量を示し、また、不作のリスクも低減する(Rao & Willey, 1980)。
ソルガム・ピジョンピース・インタークロッピング・システムの調査のように、伝統的な作物栽培システムやその他の実践を試すことで、有益な教訓を得ることができる。土地と水をより効率的に利用し、病害虫の被害を軽減する新しい混合作物システムの可能性は大いにあると思われる。適切なシステムを探すには、まず、世界各地の小規模農家が行っている伝統的な栽培方法に目を向ける必要があるのではないだろうか。研究者は、これらのシステムを現代の文脈でどのように使うことができるかを探ることができる。精密農業機器によって、新たな可能性が開かれる。
残念ながら、農業研究者、援助機関、政府は、近代科学が最もよく知っており、伝統的なやり方を、肥料、除草剤、殺虫剤などの化学物質を投入して機械化された単一栽培で栽培された新しい品種の使用を含む近代農法に置き換える必要があると、しばしば考えていた。これとは逆に、農家が伝統的にどのような農法を用いてきたのか、そしてその理由を探るというアプローチもある。これは農業人類学の研究分野の一つであるが(Rhoades & Rhoades, 2008)、持続可能な農業に重要な貢献をする可能性のある研究分野として、まだ軽視されている(Sarkar, 2017)。
1.6.2 土壌のマイクロバイオームを強化する
土壌微生物が土壌の肥沃度を高めることは、自由生活する微生物による窒素固定とマメ科植物の根粒の両方において、古くから知られている。同時に、菌根菌は植物が土壌からリンや窒素を動員するのを助け、また水を取り込む能力を高める。化学肥料を土壌に添加すると、菌根菌や窒素固定微生物の活動は抑制される。マイクロバイオームの豊かさと菌根菌の多様性は、一般に有機農業システム下で最も高い(Manoharan, Rosenstock, Williams, & Hedlund, 2017)。ここ数十年、化学肥料の使用が重視され、土壌微生物の生態には比較的注意が払われてこなかった(Hart & Trevors, 2005)。
持続可能な農業を行い、土壌の肥沃度と構造を維持することがより必須となる中、土壌微生物の生態に関する新たな研究の波は非常に有益であると考えられる。さまざまな土壌管理方法、作付けシステム、ローテーションの影響は、微生物の生態系に大きな影響を与える(Bender, Wagg, & van der Heijden, 2016)。土壌の生態系は耕作によって大きく乱され、保全農業のセールスポイントの一つは、このような攪乱を回避することである。しかし、耕作・不耕作を問わず、従来のシステムがグリホサートなどの化学除草剤に依存している限り、土壌生態系に悪影響を及ぼす可能性もある。グリホサートは菌根菌に毒性があり、菌根菌の胞子の生存率や菌根菌の根への定着率を低下させる(Druille, Cabello, Omacini, & Golluscio, 2013)。有機農法やバイオダイナミック農法はこの問題を回避しているが、それでも耕起によって土壌微生物の生態系を乱している。
保全型農業は一般に、従来の耕作農業と比較してヘクタールあたりのグリホサート使用量が少なく、多くの小農は保全型農業システムでグリホサートを使用していない。Goss, Carvalho, and Brito(2017)によるグローバルレビューでは、保全型農業システムにおいて、グリホサート散布が菌根の多様性、コロニー形成、機能に悪影響を及ぼす証拠はないとしており、その理由は、健全な土壌、土壌攪乱を行わないか最小限に抑える、保護土壌マルチカバーの維持、作物と根系の多様化などがあり、これらはすべて菌根を促進することが分かっている。一方、反転耕起は、菌根の生息環境と土壌の健全性が低下するため、土壌中の菌根と後続作物に接種する能力に悪影響を及ぼすことが明らかになった(Goss et al., 2017)。
作物に菌根菌を接種すればこれらの問題が解決すると考えたくなり、現在多くの企業がこれを目的とした「バイオイノキュラント」製品を販売している。しかし、外因性の菌根菌を土壌に添加しても、作物の成長や収量にほとんど影響がないことが多く、有害である可能性さえある。添加した生物は、すでに土壌に存在し、その特定の生態系条件で進化してきた菌根菌と競合するためだ(Hart, Antunes, Chaudhary, & Abbott, 2017)。窒素固定細菌であるリゾビウムの菌株をマメ科作物に接種する場合も同様である。無菌土壌で栽培された植物を用いた実験室での実験では、このような接種によって生育が劇的に改善されることがあるが、畑では土壌は無菌ではなく、すでに複雑な微生物生態系を含んでいる。私たちは、土壌の生態系に害を与えるようなやり方ではなく、土壌の生態系の健全性と有効性を高めるような輪作を含む持続可能な農法を必要としている。作物回転、有機および無機栄養剤の添加、耕起または不耕起の実践が土壌マイクロバイオームと菌根の動態に及ぼす影響については、多くの研究が必要である(Goss et al., 2017)。
1.6.3 人工知能を搭載した自動化機器は小規模農家を支援する可能性がある
ヨーロッパ、北米、オーストラリアなどでは、人件費削減の必要性から、畑や農場がどんどん広くなり、機械も大きくなっている。非常に大きな機械を一人で操作する方が、小さな機械を数人で操作するよりも人件費は安くなる。しかし、人件費は削減できても、資本コストは多くの農家にとって手の届かないレベルまで上昇する。例えば、2018年のトラクターなどの大型機械と合わせて、農業には莫大な設備投資や機械リース費用がかかるため、農業の経済性はますます大型化する。1000エーカー以上の農場はますます一般的になってきている。耕作地の価格が1エーカーあたり8000ポンド(1万ドル)程度であることから、このような農場の土地だけでも約8ポンドかかる。
ミリオン(1050万円)。このような状況で、莫大な相続財産や多額の資金援助がない限り、若者が農家になることはありえない。
しかし、私たちは大きな変化の入り口に立っているのかもしれない。GPSナビゲーションシステムを利用した自動運転車や自動運転農業機械の技術が急速に向上し、運転手のいないトラクターやハーベスターが普通の機械になる。また、人工知能(AI)により、精密な播種、掘削、収穫の方法が可能になる。土の状態の違いに応じて、肥料を必要な部分に適切に散布したり、雑草を認識して物理的に除去したり、害虫が発生している場所に殺虫剤を散布したり、果物や野菜を熟した時点で選択的に収穫したりすることができるのである。やがて、人件費削減のために巨大な機械で広大な農場を持つ必要はなくなるかもしれない。より小型で安価なインテリジェントマシンを導入した小規模な農場が、経済的に成り立つようになるかもしれない。圃場面積や農地面積が縮小され、作付体系が多様化する可能性がある。
また、機械化しないと人件費が高すぎて実現できない混作も、コンピュータの高度な技術で容易にできるようになる。AIを活用した新しい高度な技術により、単一栽培に代わる効率的で生産性の高い多様な混作システムが実現する可能性がある。
1.6.4 アーバンガーデンとパートタイム農業
世界のいくつかの地域では、イギリスのアロットメント(Acton, 2011)や、アメリカやカナダのコミュニティガーデンのように、プロの農家ではない人々によって、大量の食料が栽培されている。キューバやアフリカの一部など、世界の他の多くの地域では、アーバンガーデンが地域の食料生産の重要な場となっている。合わせて、現在、世界中で推定8億人が何らかの形で都市農業を実践している(Edmondson, Davies, Gaston, & Leake, 2014)。
単位面積あたりの生産性という点では、これらの庭園は農業の単一栽培よりも優れていることが多い。イギリスでは、第二次世界大戦中、「勝利のための発掘」キャンペーンにおいて、耕作面積の1%未満にもかかわらず、割り当てや庭園が消費される食料の約10%を提供した。最近の研究では、庭やアロットメントでは、従来の農作物の下で同じ面積を栽培した場合と比べて、4~11倍の果物や野菜の収量が得られることが示されている(Thompson, 2014)。さらに、イギリスのアロットメントと従来の耕作地の土壌を比較した研究では、アロットメントでは有機炭素が平均32%、全窒素が25%多く含まれていることがわかった。また、圧縮も少なかったそうだ。また、多くの耕作農家とは異なり、95%のアロットメントガーデナーがバイオマスをその場で堆肥化し、有機肥料や市販の堆肥を加えていた。彼らは多くの農家よりも持続可能な農業を行い、より高い土質を維持していたのである(Edmondson et al., 2014)。
世界のある地域では、子供が家の土地を共有する相続制度によって、家族の農場が分割され続けており、その結果、ドイツのフライブルク・イン・ブライスガウの近隣のように、都市に住む人々が近くに小さな農場を持っていて、週末の農場仕事と都市の仕事を組み合わせて、パートタイムで世話をしている人もいる。
小さな農園や庭、果樹園を街に住む人たちが利用できるようにすることで、このような食料生産が増える可能性は大いにある。世界には、都市部やその周辺で、趣味やアルバイトとして食料を栽培する機会を求めているところがある。例えば、イギリスでは、アロットメントを求める人が長く待ち、慢性的に供給不足に陥っている。
このように、都市部やその周辺にガーデニングや小規模農業ができる場所を増やすことで、より多くの食料を健康的かつ多様な方法で生産することができる。しかし、その土地はどこから来るのだろうか。
都市近郊の土地所有者は、土地を売りたがらず、建築用地として開発された場合、莫大なキャピタルゲインを失う可能性があるが、5年または10年のリースで土地を貸してくれる可能性はある。例えば、イギリスの地主が家庭菜園や果樹園のために土地を貸し出した場合、1エーカーの5分の1、つまり800平方メートルという小さな単位で貸し出せば、農家に貸して耕作用のモノカルチャーを作るよりも、1エーカーあたり少なくとも20~30倍の年間賃料を得られると計算した(Sheldrake, 2013)。また、これらの果樹園や庭の世話をする家族の肉体的、精神的な健康もおそらく改善されるであろう。
1.6.5 人間の排泄物の利用
インドのICRISATで研究をしていた頃、ヒヨコマメのフィールド調査の一部は、チベットとの国境に近いラホール渓谷のヒマラヤの村で行われた。私は、伝統的な農家にホームステイしていた。トイレは、床が高くなった部屋に穴が開いていて、そこから排泄物を藁の中に落とし、肥やしにする。これを毎年、自分の家の土地に戻すことで、養分を直接循環させていたのである。私はその時、東アジアの広い地域で何千年も続いてきた伝統的な慣習を目の当たりにし、それに参加しているとは思ってもみなかった。F.H.キングがその名著『40世紀の農民たち』(1911)で示したように、無駄なものは何もない。人間の排泄物である「夜の土」は貴重な商品であった。公衆トイレは有用な資源を集める手段であった。キングが横浜から東京に向かう旅で見たように、「鉄道の駅などでは、無駄遣いではなく保存のための設備が整っており、田舎道でも、旅人に立ち寄るよう促すスクリーンがある」(キング、1911年、9ページ)。
一方、私たちの多くは、自分たちの排泄物が、大量の水を必要とする配管システムで流され、文字通り無駄になるシステムに慣れ親しんできた。一方、尿素をはじめとする肥料は、工場で作られ、あるいは大地から採掘され、土壌に大量に施され、その流出によって河川や湖沼、海が大規模に汚染される。
人間の排泄物のリサイクルの出発点として、尿を分別収集することが考えられる。尿は処理しやすく、栄養分も豊富で、特に尿素が多い。一般的に人間は1日に12〜20g、1年で7kgの尿素を排泄すると言われている。1人あたり年間5kgという穏当な図にすると、世界の人口が約75億人なので、約380億kg、3800万トンの尿素が排泄されることになる。2016年の世界の工場での尿素生産量は年間約1億7000万トンで、af人間が生産する量の約5倍もある。つまり、尿だけでは使用する尿素のすべてをまかなうことはできないのである。とはいえ、尿からの尿素は大きく貢献できる可能性がある。一番簡単なのは、男性用小便器から尿を回収してタンクに貯蔵し、タンカーで輸送して、わらなどの窒素分の少ない生物性廃棄物の堆肥化を促進したり、嫌気性バイオマス消化槽に添加して、消化を行う微生物の増殖を促進し、メタンを生成して発電することである。
つい最近まで、ヨーロッパでは、トイレは土蔵のある家か、屋外にあった。人間の排泄物は土に還るのである。しかし、現代人の多くは田舎ではなく都会に住んでおり、集合住宅に住んでいることも少なくない。このような状況下で、人間の排泄物を文字通り無駄にするのではなく、リサイクルするためにはどうしたらよいのだろうか。一つは、トイレの排水を台所の流しや風呂、シャワーなど他の水性廃棄物と一緒にせず、トイレの配管を別にすることである。トイレ以外の排水は別々に処理することができ、より簡単に使用可能な水に再生することができる。都市部の近隣や大規模なアパートから出るトイレの排水は、地域のバイオガス消化施設に直接パイプで送ることができる。生ゴミも回収して、そこに加えることができる。地域のバイオガスプラントは、発電と熱生産の両方を行い、地域の暖房設備に利用することができる。
動物性廃棄物は、すでにこのような方法で利用されている。インド亜大陸には、家庭用のゴーバルバイオガス消化装置が約200万台あり、牛糞の消化によって一家分の調理ガスを供給し、残渣から貴重な肥料を得ることができる。中国にはさらに多くの施設がある。ヨーロッパでは、生ゴミを利用した大規模な都市型バイオガス消化設備が多数設置されている。また、牛舎から出るスラリーを消化槽に投入し、牧草などのバイオマスと混合している農場もある。これらの技術は、比較的容易に人間の排泄物を利用することができ、実際、いくつかの下水処理場にはすでに嫌気性消化槽が設置されている。
嫌気性消化槽から出る液体残渣は、窒素やその他の植物栄養素を豊富に含む優れた有機肥料であり、適切に希釈した後、液体肥料として土地に散布することができる。
1.6.6 雑草の利用
雑草とは、定義上、間違った場所に生えている植物である。作物と競合し、収穫を減少させるため、不要な植物である。しかし、雑草の中には、丈夫で元気に育つからこそ、雑草となるものも少なくない。しかし、雑草の中には、丈夫で元気に育つからこそ雑草であるものも多く、バイオマスという収穫物を得るには有利な場合もある。ほとんどの雑草は人間の食用や家畜の飼料にはならないが、中には消化器用バイオマスの生産に役立つ作物があるかもしれない。雑草をバイオガスディジェスターに利用する可能性は、ホテイアオイですでに確立されている(Almoustapha, Kenfack, & Millogo-Rasolodimby, 2008)が、これは多くの熱帯・亜熱帯地域で非常に侵略的な水草で、地球上で最も早く成長する植物種の一つである。
1.6.7 食用作物をバイオ燃料に利用することを段階的に中止する
気候変動への懸念もあり、世界の一部では、食用作物が食糧としてではなく、エタノールを中心とした液体燃料の生産に利用されている。例えばアメリカでは、トウモロコシの総生産量の約40%がエタノールに、36%が飼料に使用されている。残りのほとんどは輸出されている。アメリカ人の食用になるトウモロコシは全体のごく一部で、その多くは高フルクトースコーンシロップの形で使われている。トウモロコシから生産されるエタノールのエネルギーバランスは非常に緩やかで、エタノールに含まれるエネルギーは、トウモロコシの栽培とエタノールの生産に必要な化石燃料の約2倍である。そして、この政府の資金の多くは、米国市場を支配する巨大な食品会社に利益をもたらしている。アーチャー・ダニエルズ・ミッドランド社とカーギル社という世界最大の民間企業である。これらの企業は、米国政府へのロビー活動や、彼らに多くの利益をもたらす補助金制度の形成に多額の投資を行っている(Pollan, 2006)。例えば、流出や浸食を減らし、土壌の質を向上させ、より多様で持続可能な作付けシステムを促進するために、これらの補助金をもっと違った形で使うことができるだろう。これは主に政治的な問題だが、現在のシステムに代わる農業の開発には、大きな研究余地がある。エネルギー生産の面では、太陽光発電や風力発電のように、液体燃料に代わるものがすでにあり、電気自動車へのシフトが加速するにつれて、その重要性はますます高まるだろう。
1.6.8 人間の食生活の変化による需要削減
2011年の分析によると、牧草地を含む全農地の約75%が畜産に充てられている。穀物や大豆など、集約的に畜産された動物に与えるための作物の割合は、質量で全農作物生産の約24%、カロリー含有量で約36%にのぼるとされている。また、畜産は温室効果ガス排出量の約18%を占めている(Cassidy, West, Gerber, & Foley, 2013)。
これらすべての食料の肉への変換効率は、非常に低いである。カロリーに換算すると、動物に与えたカロリーのうち、最終的に肉などの動物性食品を通じて人間の食生活に貢献するのは約10~12%に過ぎない(Foley, 2013)。植物性食品を人間が直接食べるようになれば、増え続ける世界の人口に対して、はるかに多くの食料を供給できるようになる。最近のある分析によると、「人間が直接食べるためだけに食物を栽培すれば、原理的には、利用可能な食料カロリーを70%も増やすことができ、さらに40億人を養うことができる」家畜の飼料となる作物の配分を少し変えるだけでも、世界的に食料の利用可能量を大幅に増やすことができる(Cassidy et al., 2013)。食肉用動物の飼育には大量の水も使用される。また、肉の過剰摂取は、いくつかの種類のがん、特に結腸がんや直腸がんのリスクを高める(Larsson & Wolk, 2006)。
欧米の一部の国では、菜食主義者、ベジタリアン、「フレキシタリアン」の食生活を送る人の割合がすでに増え、肉の消費量が減っており、EU全体では、20-30年までに肉、特に牛肉の消費が減少すると予想されている。
また、ソルガムやキビのような干ばつに強い作物の使用を増やすことも、食糧供給全体に大きな影響を与える可能性がある。例えば、水が不足しているインドの多くの地域では、利用可能な水の多くが、最も水を必要とする作物である米の比較的小さな面積の栽培に使用されている。しかし、同じ水をもっと薄く、特にソルガムやキビを使って広げれば、作物の収量は増え、全体の食糧生産量は上がり、食糧供給もより栄養価が高くなる(Davis et al., 2018)。政府の政策、補助金制度の変更、マーケティング、食品産業との連携などを組み合わせることで、嗜好が変わり、米の生産を減らしてより水効率の高い作物に切り替えることで、農業システム全体がより効率的になる可能性がある。
このように、今後数十年の農業を形作る最も重要な要素のひとつは、食生活の変化である。これらは農業の領域外ではあるが、農業の未来を語る上で無視することはできない。また、農業に影響を与える経済力は、単に人間の食欲や可処分所得の結果ではなく、農業や食品企業の経済的利益やロビー活動、食品広告、そして税制や補助金などの政府の政策によって影響を受ける。肉や米の消費に課税することは、おそらくほとんどの国で政治的に困難であろうが、政府は補助金制度を変えることによって大きな変化をもたらすことができる。例えば、アメリカでは、多額の補助金が飼料用穀物、特にトウモロコシや大豆など、工場農業に使われる作物の生産を奨励しているため、間接的に食肉の集約生産に補助金を出し、他の国よりもはるかに安い値段にしている。インドでは、政府の補助金によって、他の穀物よりも灌漑用の小麦や米の生産が優遇され、水の無駄使いを助長するような市場の歪みが生じている。
1.6.9 実践的な植物育種
ダーウィンやメンデルの時代よりずっと以前から、人々は植物や動物を自然発生した変異株から交配したり、有望な親同士を交配させ、その子孫から選抜したりしていた。ペキニーズ、アフガンハウンド、牧羊犬など、多くの犬種は、遺伝学の時代よりずっと前に生まれたものである。また、ケール、ブロッコリー、芽キャベツなど、キャベツの種類も多く、これらはすべて同じアブラナ科の植物である。ダーウィンは、このような実用的な動植物の品種改良の事例から、淘汰の力について考える材料を得たのである。ダーウィンの著書『種の起源』(1859)には多くの事例が紹介されており、『家畜化された動植物の変化』(1868)にはさらに詳しく書かれている。同様に、メンデルが遺伝の原理を解明できたのは、植物育種家の活動によって、さまざまな品種のエンドウ豆がすでに存在したからだ。植物育種の成功が先だったのである。遺伝学はその後である。
20世紀の大半の間、政府は農作物の育種に資金を提供し、農業研究所や大学で研究を行った。その目的は、農家に適した新品種を作ることであり、農家は一度だけ種子を購入すれば、その後は自分たちの種子を使うことができる。例えばイギリスでは、ケンブリッジ近郊のトランピントンにある植物育種研究所が、イギリス政府から農業研究評議会を通じて資金援助を受けて多くの新品種を生み出し、その中にはマリスパイパーというジャガイモや醸造用大麦マリスオッターなど、今でも商業的に栽培されているものもあった。しかし、1980年代以降、農業研究はほとんど民営化された。植物育種研究所は1987年に民営化され、ユニリーバに売却され、1998年にモンサントに売却された。研究の量は減少した。2009年に建物は取り壊され、モンサントは土地を住宅用のデベロッパーに売却した。
植物育種や農業研究という公益的な概念は、企業の利益を最大化するという目標に取って代わられたのである。農家が前年度の種子を保存するのではなく、毎年新たに種子を購入しなければならない場合、利益は最大となる。品種改良権という古い制度では、農家は自分たちの種を翌年も使うことができる。しかし、企業は種子の特許を取得し、農家に何度も種子を買わせるというシステムを好んでいるのである。モンサントは、自分の種を使った農家がモンサントの特許を侵害しているとして、米国内の数百の農家を訴え、農家から2300万ドル以上を勝ち取ることに成功した(Harris, 2013)。
2018年、ドイツに本社を置くバイエル社がモンサントを買収した。バイエル/モンサント、シンジェンタ、デュポンの3つの多国籍企業が、世界の種子市場の大半を支配するようになった。
1980年代の民営化の波と同時に、遺伝子工学による遺伝子活用の展望に対する楽観論が高まった。従来の植物育種では、まず望ましい性質を選択し、その後に遺伝子解析を行う。遺伝子工学では、まず遺伝子を特定し、バイオテクノロジーによって植物に組み込む必要がある。例えば、殺虫タンパク質をコードするバチルス・チューリンゲンシスの遺伝子を綿花やトウモロコシに導入し、昆虫に毒を与えることができるようにした。同様に、除草剤グリホサート(モンサント社、現在はバイエル社からラウンドアップとして販売されている)を代謝するバクテリアの遺伝子が、大豆を筆頭にさまざまな作物に移植された。これらの作物を「ラウンドアップ・レディ」にすることで、畑全体にラウンドアップを散布し、この毒を破壊する酵素を持つラウンドアップ・レディの作物を除くすべての植物を枯らすことができる。現在、米国で栽培されている大豆の大半は、ラウンドアップを分解できるように遺伝子組み換え(GM)されている。このシステムは、ラウンドアップの販売を促進し、極端なモノカルチャーを作り出すが、必ずしも収量が増えるわけではない。1999年、米国で行われた8000以上の野外試験の分析では、ラウンドアップ・レディの大豆は、従来品種に比べて平均6.7%収量が少なかった(Benbrook、1999)。この「収量ドラッグ」は、他の遺伝子を巻き添えにする遺伝子組換えという技術的プロセスの副産物であると考えられる。モンサント社は、ラウンドアップ・レディの新バージョンがこの問題を克服していると主張している。しかし、同社自身のデータによれば、その遺伝子組み換え品種の収穫量は、その形質が挿入された従来品種よりも少ない。
遺伝子操作による農業の変革について多くの約束がなされているにもかかわらず、作物のほとんどの特性は、作物に組み込むことができる単一の遺伝子によって制御されていないという事実によって、実際の応用は制限されている。実際、植物と動物の両方におけるゲノムワイド関連研究により、ほとんどの複雑な遺伝性形質には、数十から数百の遺伝子が関与しており、そのほとんどが小さな影響を及ぼしていることが分かっている。これらの特性は「多遺伝子性」であり、遺伝子工学によって単一の遺伝子、あるいは2,3個の遺伝子を挿入したり削除したりしても制御できない(Sheldrake, 2020)。
同様の理由で、最近のCRISPR- Cas9システムによる遺伝子編集の技術は、専門的な場面で限定的に使用できるかもしれないが、遺伝子工学の場合と同様に、単一遺伝子、あるいは単一遺伝子内の単一塩基対に非常に焦点を当てたアプローチでは、多数の遺伝子に依存する作物植物における重要な特性に対処できない。
新しいプラグマティズムの一側面として、作物を混合物として育種することが考えられる。小麦のような特定の作物の混合品種を植えれば、純系の単一栽培に比べ、病気への感受性を減らすことができる。害虫や病気、気候の悪条件に対して、すべての植物が同じように反応するのではなく、一部の植物が他とは異なる効果的な反応を示すため、遺伝子組み換え作物は生存率や収量の面で有利になる可能性がある。例えば、中国で行われた大規模な実験では、数千の農場に稲の品種を混ぜて播種した。その結果、イネの最も重要な菌類病であるいもち病の被害が、殺菌剤を使用することなく、許容範囲内にまで減少したのである。『Nature』誌によれば、「この方法は、植物遺伝学の新しい発展に後押しされて、農業全体に広がっている極端なモノカルチャーを逆手に取ったものである」(Wolfe, 2000)のだという。
要約すると、分子パラダイム、バイオテクノロジー特許、企業帝国建設の組み合わせが、植物育種の分野を歪めてしまったということである。実用的な植物育種は、適切な場合には分子遺伝学的技術に助けられ、単一遺伝子のアプローチよりも優れた作物の育種につながる可能性が高くなる。混合栽培が可能な作物を育種することで、昆虫、病気、干ばつに対する耐性がさらに向上し、収量がより安定する可能性がある。
1.6.10 エピジェネティック遺伝とその応用の可能性
20世紀、西洋の生物学では、後天的な性質の遺伝、すなわち「ラマルク遺伝」が異端視されたことがある。それは、そのような遺伝の可能性を明確に否定し、交配集団内の遺伝子頻度の自然選択と遺伝子のランダムな突然変異に焦点を当てたネオダーウィン進化論に反するものであった。この科学的な問題は、冷戦の時代にも大きく政治的に扱われた。ソ連では、トロフィム・リセンコが指導する生物学の正統派が、後天的な特性の遺伝を強く支持していた。メンデル派の遺伝学者たちは迫害された。
しかし 2000年に入ってからは、後天性遺伝は「エピジェネティック遺伝」と呼ばれるようになり、主流になりつつある。この分野は、生物学で非常に活発に研究されている分野である。植物が、両親やもっと遠い祖先から、生育環境に応じて獲得した特性を受け継ぐことがあることは、現在では疑う余地がないほどだ。1960年代から1970年代にかけて、現在エピジェネティック研究と呼ばれるものの先駆者であるアラン・デュラントが、亜麻を用いた研究で、土壌肥沃度によって顕著かつ永続的な変化が生じることを明確に示した。例えば、窒素肥料を大量に与えて育てた亜麻は、背が高くなり、枝分かれが少なくなった。この特徴は、土壌中の窒素濃度が高くなくても、その後の世代に現れる(Durrant、1962)。デュラントは、ネオ・ダーウィンの潮流に逆らっていたが、最近の研究で、エピジェネティック遺伝の他の多くの例が明らかになっている。動物でも同様のエピジェネティックな作用が起こる(Miska & Ferguson-Smith, 2016)。
しかし、農業においては、ある品種の種子はその品種のDNAを運ぶだけのものであり、栽培された条件には影響されないとする考え方がまだ一般的である。エピジェネティクスの研究に照らし合わせると、これはありえないように思える。作物にはエピジェネティックな効果があるのかもしれない。種子や植物繁殖体が育てられた条件が、思いもよらない影響を及ぼしているのかもしれない。
実際、植物におけるエピジェネティックな遺伝に関する最近の研究は、有用な応用の可能性を示唆している(Hauser, Aufsatz, Jonak, & Luschnig, 2011)。多くの植物種には誘導性防御システムがあり、最初の攻撃を受けた後、植物はさらなる攻撃に耐える能力を高める物質を生産する。防御シグナルシステムのひとつは、ジャスモン酸とその関連代謝物の植物体内での生成と移動に依存している。シロイヌナズナとトマトを用いた実験では、植物にイモムシをつけて葉を食べさせ、種から育てた子孫のイモムシ攻撃に対する抵抗性を評価した。その結果、イモムシの成長は、コントロールに比べて約40%減少した。この抵抗性は、加害植物から生まれた第一世代をイモムシにさらさない場合でも、第二世代に引き継がれた。この抵抗性は、第3世代でイモムシの被害がなくなると消失した(Rassman et al., 2012)。このエピジェネティックに遺伝する抵抗性は、植物がジャスモン酸シグナル系を動員し、遺伝子発現のエピジェネティック変調に関与することが知られている小干渉RNAを生産する能力に依存していた(Henderson & Jacobsen, 2019)。
シロイヌナズナを用いた他の研究では、植物を干ばつにさらすこともエピジェネティックに遺伝する効果があった(Zhang, Fischer, Colot, & Bossdorf, 2013)。高温への曝露も同様であった。シロイヌナズナを用いたある研究では、親世代とF1世代で植物を穏やかな熱(30℃)にさらし、F2世代では通常の温度で栽培した。そして、F3世代で再び熱にさらした。この第3世代で、30℃で育てた植物は、祖先が熱にさらされた場合、祖先が常温で育てた対照植物に比べて6倍以上の種子を生産した(Whittle, Otto, Johnston, & Krochko, 2009)。
1930年代から1950年代にかけて、現在ではエピジェネティクスと呼ばれる現象が農業の分野で広く研究されていたのである。西側諸国では、後天的な特徴の遺伝はあり得ないと考えられていたため、これらの結果はすべて詐欺か疑似科学に違いないと一般に考えられていた。しかし、エピジェネティックな遺伝が実在することが明らかになった今、ソ連時代の生物学や農業の専門誌には多くの有益な情報が含まれている可能性があると考えられる。遺伝学、エピジェネティクス、ロシア語に詳しい人を雇って、これらの文献を徹底的に調査し、新しい研究の出発点となるような一連の総説をまとめることができるだろう。
遺伝的なエピジェネティック効果は、時間の経過とともに薄れていくことが多いので、最大の効果は、適応反応を引き起こす環境条件に親がさらされた後の世代にあると考えられる。もし、このような効果が実用化されるとしたら、このシステムは、むしろハイブリッド・トウモロコシなどの雑種作物の生産と同じようなものになるだろう。エピジェネティックに改良された種子を使うには、F1ハイブリッドのように、毎年、次の世代の種子を新たに生産することになる。このようなシステムがあれば、虫害に苦しむ有機農家が、その損失を取り戻すことができるかもしれない。虫害を受けた作物の種子収量が少ない場合、次世代に虫害に対するより高い抵抗力を付与すれば、農家はその種子にプレミアム価格をつけることができるかもしれない。
1.7 イノベーションを自由にする
現在の農業研究のほとんどは、農家や園芸家の質問によってではなく、大学や企業の科学の発展によって、そして何よりも遺伝子組み換えや編集遺伝子の特許取得や農薬の普及の見込みによって決定されているのである。補完的なアプローチとしては、農家や園芸家がどのような問題に直面し、どのような解決策を望んでいるかを尋ねることであろう。数年前、BBCのラジオ番組で、科学資金について議論したことがある。インタビュアーが、畑を持っている女性に、「もし機会があれば、研究者にどんな質問をしたいねか」と尋ねた。ニンジンを有機栽培しているのだが、ニンジン根こぶ病で苦しんでいる。ある人が、「草の刈りくずを土にまいておくと、根こぶ病の発生が抑えられる」と教えてくれた。それが本当なのか、どうすれば一番効果的なのかを知りたいということだった。そこでインタビュアーは、英国バイオテクノロジー・バイオサイエンス研究評議会という、納税者の資金を生物学的研究に回す政府機関の責任者に、このような問題を扱っているのか、あるいは扱えるのか、と尋ねてみた。と尋ねると、「作物の遺伝子組み換えの可能性やゲノムの解読など、もっと根本的で重要な問題を扱っている」と答えた。
研究によって答えが得られるこのような疑問は、おそらく何百とあるはずだ。中には、すでに答えが出ているものもあるかもしれない。もし、農家や園芸家が質問を投稿し、すでに答えがある場合はそれを受け取り、答えがない場合は新しい研究の道を開き、その質問が価値あるものであれば、これらの実用的な問題は新しい研究のための実りある刺激となり得るだろう。
もう一つのアプローチは、創造性に報酬を与えることである。世界中で何百万人もの人々が、庭や畑、農園で食用植物を育てている。その多くは長年の経験を持っている。中には、創造的で実験的な人もいる。しかし、現状では、農家や庭師がイノベーションを思いついても、それが大学の学者やグローバル企業の興味を引くことはまずない。また、一般の人が学者や研究者とコミュニケーションする可能性もほとんどない。
もし、新しい作物、新しい栽培方法、新しい園芸や農業の方法などを開発した場合に、それを賞として応募できる仕組みがあれば、プロの研究者でなくても、自主的に創造するインセンティブが生まれるはずだ。新しい作物、遺伝子組み換え植物、農薬、新しい機械などのようにトップダウンでイノベーションが起こるだけでなく、文字通り現場で調査や実験を行った人たちからボトムアップでイノベーションが起こるかもしれない。
クーンによる科学のパラダイムチェンジの分析が問題なのは、独裁を肯定しているように見えることである。新しいパラダイムが施行されると、古いモデルは否定され、信用されなくなる。新しい正統性が古い正統性に取って代わる。これは、権威主義的な政権が別の政権に取って代わられる政治革命に似た革命のモデルである。次の科学革命は違うかもしれない。新しい正統性をもたらすのではなく、寛容な多元主義への道を開くことができるかもしれない。
これこそ、私が望むところである。特定の正統派が権力や資金を独占することなく、科学研究全般、特に農業研究が、唯物論、分子勝利主義、新自由主義資本主義のドグマから解放され、土壌の質の向上と持続可能性に重点を置いて実用的に行われるとき、イノベーションは最も自由になるだろう。
5.1 はじめに
「2030/2050 年に向けた世界農業」は、世界の農業の動向を予測した主要な報告書である(Alexandratos & Bruinsma, 2012)。国連食糧農業機関(FAO)の経済部門が作成したものである。国連食糧農業機関 の著者は、その要旨において、目立つように免責事項を述べている。つまり、この報告書は、世界の食糧システムをどのように発展させるべきかの処方箋ではないのである。つまり、この報告書は、世界の食糧システムがどのように発展すべきかの処方箋ではない。これは単なる探索的モデルであり、通常通りのビジネスに関する最も信頼できる予測である (Alexandratos & Bruinsma, 2012)。
この免責条項は、おそらく、前身の報告書(国連食糧農業機関, 2006)が受けた世界的な注目度の高さから生じたものである。この「中間報告」は、2050 年までに世界が 70%以上の食糧を生産しなければならないと主張するものとして、世界中で引用された。この70%という数字は(時には「倍増」にさえ調整される)、農業の技術的近代化のための数々のアジェンダ、とりわけ遺伝子組み換え作物の推進に必ずといっていいほど採用された。こうして、2009 年に英国の主任科学者は、気候変動と食糧不足という差し迫った「パーフェクト・ストーム」 を予測したのである(Beddington 2009)。同様の分析は、科学論文、エコノミスト誌のような一般誌、そしてアグリビジネスでも繰り返された(Peekhaus, 2010; Tomlinson, 2011; Stone & Glover, 2011)。
このように、国連食糧農業機関の数字は文脈から切り離され、特別な努力や難しい妥協を必要とする壮大な挑戦として繰り返し提示された。つまり、否定的に使われていたのである。国連食糧農業機関を引用している人たちは、そんなことは言っていないかもしれない。
全人類を養うための戦いは終わった。1970年代、世界は飢饉に見舞われ、何億もの人々が、今、どのような破壊的プログラムに着手しても、餓死してしまうだろう。(『人口爆弾』ポール・エーリック、1968年)。
しかし、その意味するところは同等であった。
しかし、これは単に統計を取り違えたという単純な話ではない。2016年、同じ国連食糧農業機関の部署が、元の予測を導き出すために使用したモデリングシステム(2012年にGlobal Agriculture Perspectives System(グローバルな農業の展望システム)、GAPSと改名)をより詳細に説明した(Kavalleri et al.) 著者らは、明らかに規範的な方法で、その最新の定量的予測に注意を促し、「重要な発見は…世界の食糧生産が2005/07年から2050年までに約60%増加するはずだということである」(Kavalleri et al.、2016、1頁、強調)と書いている。Kavalleriらは、同僚たちの以前の免責事項と矛盾することによって、国連食糧農業機関が食糧危機の物語の利害関係者であることから、元の免責事項が誠実であったかどうか、国連食糧農業機関の数字の規範的使用を避けるためにもっと何かできなかったのかという問題を提起している。

本章で示された答えは、国連食糧農業機関が書いたテキストの多くに「スライド」が働いており、このスライドは国連食糧農業機関のリーダーが書いたテキストで特に問題になっているというTomlinsonの分析を裏付けるものである(Tomlinson, 2011に引用されている)。国連食糧農業機関が規範的な記述と非規範的な記述の間で揺れ動いていることを考えると、このスライドは「シャッフル」と呼んだほうがいいかもしれない。しかし、これは国連食糧農業機関が作成したかどうかにかかわらず、グローバル食糧システムのすべての定量的モデルの中心的パラドックスを完全に体現している。このパラドックスとは、国連食糧農業機関 のモデリングは本来、公平な方法で「世界の食糧および農業部門の課題を特定し、戦略的な政策展望を提供する」ために存在するはずなのに(Kavalleri ら、2016、p. 1)、GAPS が実際に行うのは食糧の数量化であるということだ。これは、農業を主に生産の問題として枠付けするものである。生産に焦点を当てることは、どんな免責事項があろうとも、貧困や食料へのアクセス、生態系コスト、社会的コストの問題を疎外するため、規範的なものである。これらは検討されないか、補助的なものである。つまり、国連食糧農業機関の報告書やモデルのタイトルは「世界農業」(World Agriculture)のように大まかなものであるが、「2015/30年に向けて」(World Agriculture: しかし、開発のための農業知識・科学・技術の国際評価(IAASTD)によれば、農業に渦巻く真の「重要」問題は生産性に関するものではない(IAASTD, 2009)。IAASTD の報告書では、生産性は目くらましであると結論づけている。IAASTD の議長 Robert Watson がその発表の場で報道陣に語ったように、農業では「Business as usual は選択肢に入らない」のである。農業の真の問題は、社会的・生態的災害を引き起こすことなく、人々を養うことができるのか、ということだ。
しかし、農民の自殺や昆虫の減少、塩害、デッドゾーン、水域の汚染など、機能不全の農業がもたらす結果は、『2030/2050年に向けての世界農業』には出てこない。さらに、『2030/2050年に向けての世界農業』は、結局のところ、生産性それ自体がモデル化の関心を集めるに値することを示すことさえできていない。第一に、このモデルが記述しているのは、必要な生産性の向上は、ビジネス・アズ・ユー・グルーで解決されるという予測だからである。第二に、このモデルは、2050 年までに「栄養不良人口の小幅な減少」も予測しているが、この減少は、経済成長の継続(すなわち、富の増加)に依存しており、農業生産に依存していない (Alexandratos & Bruinsma, 2012)。
国連食糧農業機関の以前の版、World Agriculture Towards 2015/2030は、ほぼ同じ結論に達していた。当時8億5000万人いた飢餓人口のうち、2030年までに食糧生産が目標の70%増に達した場合、飢餓から解放されるのはわずか1億2000万人だと予測していた(Bruinsma, 2003)。
つまり、国連食糧農業機関自身のモデルによれば、生産量を増やしても飢餓は解消されないということだ。この結果は、アマルティア・センがその有名な歴史書『貧困と飢饉』の中で述べた結論と同じである。飢餓や飢饉が発生したとき、生産不足が原因であったことは事実上一度もないことを彼は発見している(Sen, 1981)。多くの食料システムのコメンテーターにとって、このことは疑問の余地なく決着している(例えば、Lappé & Collins, 2015)。しかし、この発見は、国連食糧農業機関のリーダー(Tomlinson, 2011)を含む多くの人々によって無視され、代わりに国連食糧農業機関を引用して、生産性重視の必要性に伴う欠乏の物語を支持してきた(例えば、Conway, 2012)。このように、GAPS のようなモデルの知見にほぼ純粋に依拠している欠乏の見方は、せいぜい曖昧な支持しか得られていないのである。
5.1.1 世界の食糧モデルと予測
しかし、本章の目的は、GAPS のようなモデルについて、その基礎となる仮定のレベルで定量的な批判を行うことだ。
特に断りのない限り、国連食糧農業機関のGAPSに焦点を当てることにする。実際、食料をカロリーや重量として定量化することは、食料システムを完全に理解する上で有害である。むしろ、国連食糧農業機関の仕事が最も多く引用され、国連食糧農業機関のシャッフルに後押しされて、世界が圧倒的にこの70%という数字を規範として解釈していることを認識するものである。2012年、国連食糧農業機関の予測は60%に更新されたが、これは主にベースラインの変化を反映したもので、2003年当時と比べると、2050年にかなり近づいている(Alexandratos & Bruinsma, 2012)。であるから、一般的なアプローチを正当化するように見えるかもしれないが、これらのモデルが最も明らかに挑戦的であるのは、純粋に量的なレベルなのである。
1798年のマルサスは、食糧システムの最初の数学的モデルを作ったと考えられている。彼の単純な予測では、指数関数的な人口増加は、最終的に直線的な供給増加を上回ると結論づけた。それ以来、彼のモデルの基本形は、食糧供給と食糧需要を分離することであった(McCalla & Revoredo, 2001)。
国連食糧農業機関 以外にも、国際食料政策研究所(IFPRI)などの機関が独自のモデルを開発している(Robinson et al.、2015)。さらに,ミレニアム生態系評価,水管理の包括的評価,アグリモンド(2009)(フランス国立農学研究所と国際農学研究センターの共同プロジェクト)などの特別プロジェクトは,一般的な手法を拡張しつつ,水制約,気候の影響,特定の政策決定の効果などの特定の問題の調査に重点を置いている(de Fraiture et al, 2007; Fischer et al., 1988; Rosegrant et al., 1995, 2001; Parry et al., 2004; Chaumet et al., 2009)。これらはすべて、意思決定に資することを目的としている。しかし、多様な潜在的結果を探索するのに適しているものは、しばしばシナリオと呼ばれる。これらの区別は、モデルの長所と短所とともに、Reilly and Willenbockel(2010)およびWise(2013)によって以前にレビューされている。
これらのレビュアーが注目するのは、何よりも、モデルやシナリオの間で、生産を強化するための特別な措置は必要ないという強い一致が見られるということだ。国連食糧農業機関 の言葉を借りれば、「世界の生産ポテンシャルの観点からは、乗り越えられない制約はないはずである」(Alexandratos & Bruinsma, 2012)である。
そのシナリオに何らかの不始末を明確に盛り込まない限り、古典的なエールリッヒ式の危機を予見しているものはない。例えば、Millennium Assessmentは、4つのシナリオの一つとして、低協力と高貿易障壁を想定した「強さによる秩序」(OS)を掲げている。OSでは、世界的な食糧不足は生じないが、栄養失調が増加し、アフリカの一部では内戦も発生する。
これはおおむね安心できる結論であるが、それでも、気候変動の影が迫っていることには注意が必要である(Battisti and Naylor, 2009; Nelson et al.、2010)。しかし、食糧生産が(気候を除けば)危機的状況に発展する可能性は低いという合意は、メディアにおける警鐘的な物語を追放していない(例えば、Hincks, 2018を参照)。
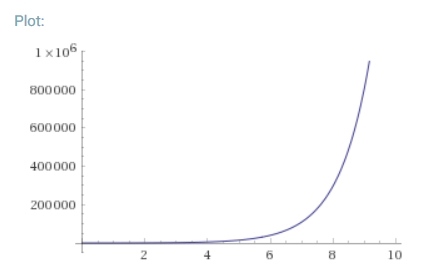
しかし、増産の必要性を予測するこの収束でさえ、過度に悲観的であると推測する根拠がある。2011年、世界銀行研究所の研究者たちは、世界はすでに140億人分の食料を生産していると提案した(Herren et al.、2011)。この数字は、国連の人口事前予測(2050年に100~110億人に達し、おそらくその後は減少する)を大きく上回っている(UN, 2017)。

さらに、モデルは世界の食糧価格の動向とも矛盾している。米国とEUのバイオ燃料政策の変更によって引き起こされた2007/2008年の価格高騰(de Gorter et al., 2015)以前は、食糧価格は年率約4%で下落していた。そのスパイク以来、価格はほぼその軌道に戻ったように見える。この長期的な下落は、農業のあらゆる部門にわたって、食糧供給が現在の食糧需要を大幅に上回っており、むしろそのギャップが拡大していることを強く示唆している。この過剰の正確な程度は明らかではないが、2017年の国連食糧農業機関による世界の穀物在庫の推定値は7億6200万トンである。この量は、世界の年間生産量の約3分の1に相当する(国連食糧農業機関STAT)。このように、どのようなモデル化にも関係なく、最も楽観的なモデルでさえも悲観的であると推測する強い根拠がある。需要を過大評価しているのか、供給を過小評価しているのか、あるいはその両方なのである。そこで問題となるのは、次のような点である。GAPSや他のモデルが主張する、より多くの食料を生産する必要があるという予測と、低い(そして低下している)食料価格や持続的な世界的商品余剰をどのように調和させるのか?モデルに欠陥があるのだろうか?もしそうなら、それはどのような欠陥なのか?
5.1.2 食料システム・モデルにはどのような欠陥があるのか?
非常に複雑なモデルを使用すると、常に、それが現実をどれだけよく表しているかという多くの疑問が生じる(Scrieciu, 2007)。しかし、フードシステムのモデルは、生物物理学的、社会的、経済的、および制度的な要素を統合しようとするため、特に複雑である。したがって、このようなモデルに対して時々なされる批判は、栄養の尺度としてカロリーを使用していることである(例えば、Herforth, 2015)。栄養学者も、食料安全保障のより広範な定義を求める人々も、カロリーは1996年の世界食料サミットで採択された食料安全保障の定義に大きく欠けることを指摘している。「食糧安全保障は、すべての人々が、いつでも、食事上の必要および食物の嗜好を満たすのに十分な、安全で栄養のある食糧を物理的および経済的に入手できるときに存在する」(例えば、Burchi et al.、2011)。したがって、カロリー充足の達成は、最終的には無意味なことかもしれない。
このような多様な批判に対応するため、定量モデルの主要な問題は、技術的不確実性、方法論的不確実性、認識論的不確実性に分けられることがある(Funtowicz & Ravetz, 1990)。
より狭い技術的な懸念としては、Reilly and Willenbockel(2010)やWise(2013)が提起した「知識のギャップと優先順位」がある。このレベルでは、これらの研究者は、データの質に非常に大きな問題があることに同意している。多くの国では、貧困、GDP、水の利用可能性、単純な人口など、モデルの最も基本的な入力値さえも定量化されている。疑問のあるデータの問題は、ガーナの事例で明らかになった。ガーナの統計局は2010年、将来のGDP予測を60%以上上方修正することを発表した。これにより、ガーナは文字通り一夜にして低中所得国に転落したのである(Jerven, 2012)。国連食糧農業機関のFood balance sheets: a hand-book (国連食糧農業機関, 2001)では、このような困難さが認められている(最終的には否定されているが)。
また、データポイント間の関係をモデル化する際にも、同程度の問題がある。ReillyとWillenbockelによれば、以下のとおりである。「統合評価研究で使用されるモデル・コンポーネントの妥当性検証に関するさらなる研究が必要である。特に、気候変動やその他の突然の変化がある場合、モデルを過去の経験に合わせて校正することは、一般に「オーバーフィット」として知られる問題を引き起こすからである。これらの「技術的」なレベルの問題のいくつかは、モデル作成者自身も認めている(例えば、Bruinsma, 2003)。
さらに重要な問題は、多くの異なる国から送られてくるため、モデルの別々の部分のデータセットが、異なるスケール、時間軸、概念的スキームに基づいていることである(国連食糧農業機関、2001年)。しかし、このような不一致を調整する方法はない。当然のことながら、これらの限界を論じた上で、ReillyとWillenbockelは、「モデル出力は、明確に定義された信頼区間を持つフォアキャストとして誤解されるべきではない」と結論づけている。むしろ、それらは、高度に相互依存的なシステムにおける複雑な相互作用と、一般的な大きさの効果の可能性について、定量化された洞察を提供することを意味している」(Reilly & Willenbockel、2010年)。このコメントは、いくつかの重要な問題を提起している。第一に、これらの数字を引用している人たちは、この制限を一般に見逃しているようである。第二に、信頼区間がなければ、モデラー自身を含め、誰もこの「効果の一般的な大きさの順序」が何であるかを知ることはできない。
このような不確実性が予測の質にどのような影響を及ぼすかについては、Thomas Hertelが米国農業応用経済学会の会長講演で行った幅広い批判を検証することで知ることができる(Hertel, 2011)。Hertelは、国連食糧農業機関やその他のモデルの主要な前提は、価格、需要、供給をどのように関連付けるかに関係していると指摘している。国連食糧農業機関の定量モデル(GAPS以前)に焦点を当てると、価格上昇に対して農業供給がほとんど反応しないことを前提としていることが分かる。この仮定が導入されたのは、農業において食糧供給が需要にどのように反応するかという測定が、ほとんど短期的に行われたからである。しかし、これらのモデルの目的は、より長期的なものである。長期的な測定は、弾力性の図式がまったく異なることを示唆している。Hertelは、将来の需要の伸びが食糧価格の上昇に十分見合うものであれば、それが供給を刺激することになると論じている。したがって、農産物価格が上昇すれば、高い収量が見込まれ、地価が上昇し(既存の土地を保護し、より多くの土地を生産に回す)、農業文化研究が刺激され、廃棄物が削減されると期待でき、これは実際に長期的な証拠が示すとおりである(Hertel、2011年)。国連食糧農業機関 によれば、将来の食料供給力の主要な決定要因である世界の作物収量の伸びの減少傾向でさえ、価格の関数である可能性がある。この点について、Hertel は経済学者の Robert Herdt の言葉を引用して、「収量を大幅に上げる経済学は魅力的ではない」(International Rice Research Institute, 1979)、と述べている。この関連で、Hertel は 国連食糧農業機関 の経済学者 Jelle Bruinsma の言葉も引用している。「適切なインセンティブがあれば、2050 年の穀物と油糧種子の需要増加の大部分は既存の技術で満たすことができるだろう」。したがって、Hertelは、よく指摘される長期的な 「収量の伸びの鈍化は、単に純デマンドの伸びの鈍化によるものかもしれない 」と結論付けている。そして、彼はこうまとめている。「得られたモデルが、ここで想定しているような長期的な持続可能性の分析に適していることは明らかではない。」 とまとめている。
Hertelを要約すると、農家やその他の意思決定者に作用するインセンティブが農業生産性を説明する鍵であるという強い証拠があるが、コムモディティ価格が長期的に下落しているため、国連食糧農業機関は低インセンティブシステムをモデル化してきたということだ。
このことを背景に、次節では予測モデルの基礎となる4つの仮定を追加し、GAPSを例にとって分析することにする。
5.2 GAPSの前提条件
GAPS の主要な前提条件は、将来の生産ニーズの予測に強く影響するもので、次のとおりである。
前提 1:バイオ燃料は “需要 “によって駆動される。
2002 年以降、液体バイオ燃料(エタノールとバイオディーゼル)の分野は急速に発展している。米国農務省の数字によると、米国のバイオ燃料産業は2011年に1億2700万トンのトウモロコシを消費し、これは世界のトウモロコシ生産量の15.6%にあたる。さらに、ブラジルではサトウキビの50%以上がバイオ燃料に利用されており、2009年にはEUが900万トンの植物油(主に菜種)と900万トンの穀物をバイオ燃料として消費している(Alexandratos & Bruinsma, 2012)。特にエタノールについては、2016年の世界の生産量は、数百万ガロン単位で、米国(15250)、ブラジル(7295)、欧州連合(1377)、中国(845)、カナダ(436)である(Mohanty & Swain, 2019)。

バイオ燃料ブームの最初の帰結は、グローバルなサプライチェーンを通じて伝播し、ほとんどの商品に影響を及ぼす価格スパイクであった(de Gorter, Drabik & Just, 2015; McMichael, 2009)。こうした価格高騰は、暴動、飢餓、財政難を誘発し、一部の国では政治的混乱が生じた。大量の農産物がバイオ燃料に転用されるという混乱は、しかし、最終的には物理的なサプライチェーンによって吸収された。現在、コモディティはほとんどスパイク前の価格に戻っている(OECD-国連食糧農業機関, 2016)。
国連食糧農業機関のモデルでは、バイオ燃料は需要とみなされている。つまり、入手可能性を減らすが、人口を養うことには貢献しない(特定の残渣を動物に与える場合を除く)。国連食糧農業機関 はさらに、将来を見据えて、バイオ燃料は面積を拡大し続け、2020 年にプラトーになると想定している(Alexandratos & Bruinsma, 2012, p.97)。
バイオ燃料を需要として扱う正当な理由は、バイオディーゼルやバイオエタノールは、意図的な政策選択による環境的に有益な産物であるというものである。この立場には大きな疑問がある。第一に、パーム油や大豆からのバイオ燃料やトウモロコシからのエタノールは、生物多様性に甚大な損失をもたら す。さらに、温室効果ガスの純排出量は、数百年にわたって測定した場合、何の便益ももたらさないか、あるいはプラスにしかならない(例えば、Germer & Sauerborn, 2008; Danielsen et al, 2009)。第二に、バイオ燃料政策の発展は、気候への懸念ではなく、農業・文化需要を高めてより多くの投入物を販売できるようにしようとするロビー活動の複雑な凝集体によって推進されてきたため、必要性の正当化は疑わしい(Baines, 2015; ActionAid, 2013)。真の需要ではなく、ロビー活動によって動かされる市場についての重要なポイントは、予想通り食 料需要が増加した場合(および/または食料価格が上昇した場合)、バイオ燃料に対する関心は消滅する ことだ。なぜなら、需要を下支えすることがバイオ燃料の重要な政治的魅力であるからだ(Baines、2015 年)。言い換えれば、バイオ燃料に充てられた土地は、必要であれば人口を養うために利用できるが、GAPS などのモデルでバイオ燃料を需要として分類すると、この機会が見えなくなってしまうのである。バイオ燃料に転換される食料の量は非常に多い。例えば、ActionAid は、G8 諸国(ブラジルを除く)が年間消費するバイオ燃料の量は、4 億 4100 万人分に相当すると結論付けている(ActionAid, 2013)。今日、この数字が測定されれば、間違いなくもっと大きくなっているはずである。
国連食糧農業機関のモデラーがバイオ燃料の転用の倫理に触れていないのは不思議なことだ。第一に、飢餓の世界では、食糧の入手可能性を犠牲にしてバイオ燃料を転用することで生じるジレンマは、かなり明白なものであるため(Chakrabortty, 2008)、第二に、国連食糧農業機関エコノミストは、そのモデルを論じる際に、様々な行動の環境および倫理への影響について脱線し、非典型的スタンスを取っている(例えば、Alexandratos & Bruinsma, 2012, p. 131)ためである。第三に、GAPS のようなモデルの前提が、生産の増大が倫理的要請であることを考えると、輸送のために燃やすことによって食料の入手可能性を減らすことが、なぜ同様に倫理的に問題でないのだろうか。国連食糧農業機関のモデラーたちは、非典型的アプローチの適用において、またしても一貫性のない態度をとっている。
国連食糧農業機関 が、バイオ燃料を何億人もの人々への潜在的な食糧供給源として取り込まず、「デマンド」としてモデルに組み込んだことは、一面では誤りである。しかし、これはまた、規範的なモードの間を行ったり来たりする矛盾の一例でもある。国連食糧農業機関のエコノミストたちが、強力な利益団体と協調するときだけ明確な規範的スタンスを取るのは偶然だろうか。同時に、このシャッフルは、GAPSがその基本的なパラダイムとして、生産がすべてに優先するという規範的な期待を持っていることによって、非常に促進されている。
前提2:現在の農業生産システムは生産性に最適化されていること
国連食糧農業機関の統計によれば、もし私たちが本当に世界の収穫量(1日当たりのカロリーで測定)を最大化しようと思ったら、みんなサツマイモ(70×10(3)kcal/ha/day)を食べることになるだろう。あるいは、赤道から離れた場所に住んでいれば、ジャガイモ(54 × 10(3) kcal/ha/day)である(国連食糧農業機関, http:// www.国連食糧農業機関.org/docrep/t0207e/T0207E04.htm)。もちろん、これらの作物だけを食べない理由もある。その理由は、輪作による農学的な利点から、多様な食事に対する身体的・文化的な必要性まで、多岐にわたる。しかし、農家は、コーヒーやワイン用のブドウなど、人々を養う能力が非常に低い作物も多く栽培している。その結果、世界の農業は作物代替によって生産性を大きく向上させる可能性を持っている。
この可能性は、米国や欧州連合などの先進国には明らかに存在する。そこでは補助金や市場の独占が、カロリーや栄養よりもはるかに強力な生産推進力となっている。しかし、バングラデシュ(ニューヨーク州ほどの面積に1億6千万人の人口を抱える)のような国は違うかもしれない、と考える人もいるかもしれない。バングラデシュは世界で最も人口密度が高く、貧困と食糧難の割合が最も高い国の一つである。しかし、バングラデシュの条件下では、小麦の収量は冬期米の約半分だが、小麦の市場価格は高く、投入コストははるかに低い(J. Duxbury, 私信)。そのため、バングラデシュの農家は415,000haで小麦を栽培している(国連食糧農業機関STAT, 2017)。そのような農家は、栄養ではなく市場を追い求めているのである。
しかし、農業における最適でない栄養パフォーマンスの最も顕著な例は、おそらく肉の消費である。歴史的に、肉と酪農の生産の多くは、他の形態の作物栽培にあまり適さない、あるいは適さない限界的な土地を利用していた。しかし、特に多くの「先進」国では、優良な土地を家畜の飼料用に作付けすることが増えている。この飼料の最も効率的な変換者(魚と鶏)でさえ、ヘクタール当たりのカロリーリターンは、最も栄養価の低い野菜よりも悪く、最も効率の悪い(牛肉)からは、またおよそ4倍もの収穫がある(Cassidy et al.、2013)。したがって、肥育システムにおける牛肉の飼料変換効率(カロリーで測定)は、3%(鶏肉は 12%)と推定されている(Cassidy et al.、2013)。米国のトウモロコシ作物の約 35%が家畜飼料に使用されていることから、このことは明らかに代替の大きな機会を意味する(Baines, 2015)。

こうした多様な機会にもかかわらず,高収量作物の低収量作物への代替は,GAPS では無視されている可能性である。これに対して,GAPS は逆方向の代替を認めている。これは,GAPSが所得上昇に伴う肉の消費量の増加を許容する場合である。このように、GAPSは再び矛盾している。
この代替を認める標準的な正当化理由は “消費者需要 “である。人口が豊かになると、「自然に」肉を食べるようになる、という理論である。ベジタリアンの多い国を除けば、確かに豊かさと肉の消費量には相関がある。注意しなければならないのは、OECD諸国は年間3180億ドルを農業補助金に費やしていることだ。これらは、圧倒的に食肉かバイオ燃料の支援に使われている(OECD, 2002)。野菜や果物への助成は皆無に等しい。したがって、「消費者に帰属する選択の力と自由には疑問がある」し、他国も食肉生産への助成を決定しない限り、この路線に忠実に従うという単純な期待もできない(Rivera-Ferre, 2009)。
GAPSが見落としている作物代替によって、どれだけの食料が増えるのだろうか。どのような作物を代用し、何を代用するかに大きく依存するため、どのような計算も困難である。最も集中的に研究されているのは、食肉消費による影響である。最近の推定では、世界のフィードチェーンから動物がいなくなれば、40億人がさらに養えるようになる(Cassidy et al.、2013)。しかし、この研究では、動物の飼育を最も熱量の高い作物に置き換えてはいない。この結論は、主に欧米式の飼育と消費パターンを想定していることは注目に値する。したがって、遊牧民や半遊牧民のシステムが主流である世界の地域には当てはまらない。
もうひとつの代替は、混作、アグロエコロジー生産システム、保全農業の利用が拡大していることだ。これらはGAPSで想定される単収をさらに上回り、時には劇的に収量を増やすことができる(Sampson, 2018; Kassam et al.2009)。2015/16年、保全農業は世界で約1億8000万ヘクタールの農地を占め、2008/09年以降、毎年1000万ヘクタール以上拡大している(Kassam et al.、2018)。
結論として、一般的なモデル(GAPSだけでなく)は、何十億人もの人々を養う可能性を持つ作物代替のための明確な機会を無視している。
前提3:最大 “収量ポテンシャル “の存在
GAPSやその他のモデルに見られる重要な前提は、”収量ポテンシャル “という概念を使用していることだ。収量ポテンシャルとは、作物には遺伝的な収量の上限があり、それを超える作付けシステム(通常は肥料や化学物質の投入が想定される)は作物を持ち上げることができないという理論である。この上限があるため、GAPSでは、生産性の向上は2つの方法でしか実現できないと仮定している。長期的な研究と育種努力によって各作物の収量ポテンシャルを引き上げるか、あるいは、「最適でない」種子や方法を用いているためにこの上限をはるかに下回っている農家を説得して、現在の収量ポテンシャルに近づけるか、である。このように、GAPSでは収量ポテンシャルが重要な位置を占めている。国連食糧農業機関はこの仮定を多くの箇所で述べている。「新たな緑の革命や、一度限りの飛躍的な収量増加の可能性は、かなり限られているという認識がある」(Alexandratos & Bruinsma, 2012, p.125)。
しかし、こうした収量の可能性はあくまで理論的なものである。存在することが証明されておらず、存在しないかもしれない。その代表的な例が、世界で最も重要な作物である米であろう。コメの潜在収量は、標準的に 1 ヘクタールあたり 8~10 トンと見積もられている(Peng et al.) このような高い収量は、肥料や化学物質の投入量が非常に多く、土壌や散水体制が理想的な農学的条件のもとでのみ発生すると想定されている。しかし、米の生産量の世界記録は 22.4 トンである(Diwakar et al.、2012)。この記録は、SRI(System of Rice Intensification)と呼ばれる方法を用いて、ある農家が少ない投入量で達成したものである。この記録が意味するのは、専門家によるSRIの研究でも裏付けられているように、標準的な「最適」条件で栽培した場合、コメはその想定収量をはるかに下回る(あるいは収量ポテンシャルは現実世界では意味を持たない)ことである(Kassam他、2011; Taylor & Bhasme、2019)。

SRIによって達成された収量の意味は、持続可能な収量と生産性が、すべての定量的なグローバルモデルによって想定された収量の数倍を上回るということだ。理論上の収量が実際の農場で満たされることは通常ほとんどないため、22.4 tは実際には標準的な予想の3倍の収量の可能性を示している。SRIの手法は他の作物にも適用され、ここでも大幅な収量の向上が見られた(Abraham et al.、2014)。2013年、SRIの実践者は950万人と推定された(Uphoff, 2017)。2019年には、この数は少なくとも2倍になっている(N. Uphoff, pers.) 収量ポテンシャルの概念を受け入れ、SRIを無視することで、定量モデルは農業で最も急速に広まっている開発の一つを見過ごしている(Stoop et al.、2017)。
米は地球の半分(35億人)の主食であるため、収量が3倍になれば、特にSRIはより持続可能な方法であるため、おそらくさらに70億人を養える可能性があることは容易に理解できる(Fageria, 2007)。
前提 4:世界の食糧生産は、世界の食糧消費とほぼ等しい。
経済のほとんどの分野とは異なり、農業生産は、世界規模でも地域規模でも、消費量を上回ることができる。穀物(小麦、米、大麦、キビ、ソルガム、オート麦)を例にとると、インドや中国のような人口密度の高い国でも過剰生産が起こっている。2017年、国連食糧農業機関は世界の穀物貯蔵量を7億6200万トンと推定しているが、これは世界の穀物総生産量(2017年)2億5900万トン(国連食糧農業機関STAT)に対するものである。
この備蓄は、災難に対する保険となる。しかし、この7億6200万トンは、世界の需要に対する供給過剰を意味するものでもある。これらのストックの重要な特性は、その腐敗性である。気候や貯蔵の質、作物の種類によっては、腐ったり、ネズミや虫に食われたりする可能性があるため、ストックが育っていなくても、作物は高い確率でそこに入っていく。第二の性質は、年に何度も収穫がある場合、失われる在庫の量は、年換算した在庫の定常量の何倍にもなる可能性があるということだ。例えば、米の収穫の33%が貯蔵中に失われ、3回の収穫に対応する3期分の米の貯蔵がある場合、年間総在庫の100%が毎年失われることになる。このような性質から、必要なときに必要なだけ使えるように、ストックに入る量をカウントすることが非常に重要である。また、上手に貯蔵しても、いずれは劣化してしまう。中国では、小麦の在庫は最大で3〜4年、平均で2年程度と分析されている。このため、米と小麦のストック量が多い中国は、小麦の余剰ストックを消費するバイオ燃料政策を開始した。これは着実に成長し、現在では年間8億4500万ガロンのエタノールを生成している(Mohanty & Swain, 2019)。このプログラムにもかかわらず、中国の小麦在庫はまだ増え続けている。
このことが最終的に意味するのは、毎年どれだけの在庫がエンドレスになっているかを理解することが重要だということだ。残念ながら、モデル化のために、国連食糧農業機関は「世界レベルでは生産量と消費量が等しい」と仮定している(Alexandratos & Bruinsma, 2012; 国連食糧農業機関, 2001も参照)。この仮定は、2016年のGAPSの説明文に再掲されている。「このモデルは、閉じた世界経済を仮定しており、すべてのシミュレーション期間の終了時に、世界の需要が供給と等しくなる」(Kavalleri et al.、2016)。在庫に入る量はカウントされるが、開始在庫と終了在庫の差としてのみカウントされる(国連食糧農業機関, 2001)。正味のところ、昆虫、カビ、ネズミ、経年変化による損失は無視される。
この方法がいかに損失や劣化した資源を無視しているかを示すには、例を挙げるのが簡単である。1月1日に7億6200万トンの在庫があり、翌年にネズミや虫で全部失われ、翌年に全部入れ替えた場合、国連食糧農業機関のカウント法では在庫に変化はなく、在庫も追加されていないことになる。つまり、7億6,200万トンも在庫があるにもかかわらず、何も在庫がないように見えるのである。

世界の穀物供給のうち、貯蔵中に失われるものはどのくらいか?収穫後のロスを推定することは難しく、優先順位も低いようである。さらに、多くの推定値は必ずしもストックに適用されるものではなく、むしろポストハーベスト一般に適用されるものである。国連食糧農業機関は、低中所得国における穀物のポストハーベスト・ロスは約6.4%と推定している。ほとんどの穀類とパルスの損失推定値はもっと高いが、非常に変わりやすく、多くの不確実性を認めている(Boxall, 2001; Kumar & Kalita, 2017; Sharon et al, 2014)。推定値には、アフリカのトウモロコシの20%~30% (Tefera et al., 2011); 西カメルーン高地のトウモロコシの12%と44% (Tapondjou et al., 2002 ); インドのコメの11%~17% (Alavi et al., 2012); そして35% (Scrimshaw, 1978) が含まれる。例えば、サハラ以南のアフリカでは90日後に59%など、非常に高いレベルを推定している報告もある(Kumar & Kalita, 2017)。このように、国連食糧農業機関のポストハーベスト・ロスの数値は、特に貯蔵が最も劣化する段階と考えられることが多いため、非常に低い水準にある(Kumar & Kalita, 2017)。
GAPSは、モデルのバランスをとるために、互いに打ち消し合う2つの誤りを犯していると言えるだろう。それは、ストックに入るものを無視し(変化しない限り)、ストック内の損失を無視することだ。これは、閉じたモデルを構築するという観点からは良いのだが、食糧の入手可能性を推定するという目的からすると、この誤差の意味は非常に大きい可能性がある:数えきれないほどの年間過剰生産があり、それは貯蔵で失われるが、将来の需要を満たすために利用可能である。マルサス的な言い方をすれば、人口は現在の生産量を下回り、人口(少なくとも需要)は両者が等しくなる前に増加する余裕がある、ということだ。これは、ストックや備蓄が望ましくないとか不要だということではなく、非常に大きな知られざるギャップが存在する可能性が高いということだ。
現在の推定では、ストックもストックロスも非常に不確かである。しかし、国連食糧農業機関 の数字を額面通りに受け止めれば、より腐りやすい(非穀物)作物の損失を考慮しなくても、おそらく10億から20億人の穀物需要に相当する食料が過小評価されていることになる。
要約すると、ここで論じた4つの仮定は、(国連食糧農業機関のGAPSの推定値を上回る)潜在的に利用可能な余剰食料の推定値を以下のように算出する。仮定1:5億人、仮定2:40億人、仮定3:70億人、仮定4:10億-20億人。仮定1はデータが古いため、現在のバイオ燃料を過小評価している。仮定2は、食肉から作物への代替のみを含み、高カロリー作物から低カロリー作物への代替は含まない(研究が存在しないため)。仮定3はコメのみを含み、SRIは他の収量上限も同様に欠陥がある可能性を示唆している。それにもかかわらず、125 億人分の予備の食糧は非常に大きな過小評価である。
5.3 モデルは何のためにあるのか?
以上の議論から、国連食糧農業機関 の GAPS のような定量的モデルは、疑わしい仮定に大きく依存していることが浮き彫りになった。簡単にまとめると,第一に,GAPS は,高収量の食料種を低収量の食料種で代用する可能性を十分に考慮していない。第二に、GAPSは、必要であればバイオ燃料の代わりに食料を栽培することが可能であり、またそうすることができることを無視している。第三に、GAPS は、既存の作物、特に世界人口の半分が主食とする米の収穫量を、不当に低く見積もっている。第四に、GAPSは、貯蔵で失われた年間余剰分を無視している。
GAPSと関連する食糧モデルに対するこの批判には、Thomas Hertelの批判も加えるべきである。彼は、国連食糧農業機関が、食糧の有効性が価格とともに上昇する可能性を過小評価していると主張している(Hertel, 2011)。
したがって、本章の執筆のきっかけとなった、生産過剰を示す長期的な世界価格(下落)と、迫り来る欠乏を予測する定量的モデルとの間の不一致は、容易に解決することができる。モデルには欠陥がある。実際の、あるいは潜在的な供給を過小評価し、需要を誇張しているのだ。
科学、技術、経済における定量的モデルは、かつてないほど普及している(Porter, 1995)。この上昇を説明するために、2 つの異なる考え方が生まれた。一つは、定量的モデルは現実を客観的に近似したものであり、コンピュータなどの発展により、より強力で有用なツールとなったため、より普及した、というものである。
もう一つは、定量モデルが客観性を装うために普及したというものである。1933年にイギリスの科学者ホグベンが書いたように、モデルによって「事実無根の仮定を、完璧な代数学の印象的なファサードの背後に隠す」ことが可能になるのである。(別の表現では、生態系モデラーであるディック・レビンス(Dick Levins)が言っている。「定量的モデルはすべて質的モデルを装っている」。) このように、モデルは意識的あるいは無意識的な組織的偏見や目的のための煙幕(偽装)として機能することができるのである。セオドア・ポーターの言葉を借りれば、「定量化とは、決定しているように見せずに決定を下す方法である」(Porter, 1995)のである。
モデリングの台頭に関する標準的な説明には、少なくとも部分的に欠陥があることは確かである。すべてのモデリングは、複雑さを均一性に、品質を量に還元する現実からの後退である。データを削り、相互作用を単純化する際には、選択をしなければならない。おそらく、すべての定量的モデリングの中で、その構成要素が生物学的、気候学的、水文学的、経済的、社会的である食料システムの表現ほど単純化が問題となるものはないだろう、つまり、それらは非可算である。したがって、モデル作成者は、無数の潜在的な寄与要因のうち、どれを含め、どれを除外するかという難しい選択に迫られるのは必然的である。同様に、これらの要因の間の関係を数学的に単純化する方法も考えなければならない。このような選択は、客観性に対する挑戦である。
食料モデルに適用されるこの客観性の問題の好例が、国連食糧農業機関の年次会計の時間枠である。国連食糧農業機関の農業年は、1月初旬から12月末までである(国連食糧農業機関, 2001)。このため、ヨーロッパと北米のニーズに合わせて調整されており、南半球や熱帯のニーズには対応していない。この選択は、西洋の価値観、国際貿易の価値観、そして金融の価値観が、このようなモデルの中に微妙に存在し、これらのモデルが奉仕すると主張する貧しい国々(主に南半球と熱帯)の、あまり目立たないか定量化できる優先順位を犠牲にしていることを例証するものである。

したがって、客観的なモデラーという主張は成り立たない。このことは、すべてのモデルが必ずしも無価値であることを意味するものではない。しかし、モデルの作者と資金提供者の誠実さと無関心さについて、より微妙な問いを投げかけることを示唆している。このようなモデルの作成者は、無意識のバイアスの可能性を最小限にするためにどのようなステップを踏んでいるのだろうか?
この観点から、国連食糧農業機関が「可能な限り」社内の専門家に頼っていると主張するのは心もとない(Alexandratos & Bruinsma, 2012)。また、国連食糧農業機関が特に著名な場において、そのデータを高い信頼性で示すこともない(例えば、Diouf, 2008, Tomlinson, 2011に引用されている)。一方、国連食糧農業機関の文書では、仮定、限界、不確実性についての言及は少なく、中ページや裏ページ、付属文書に追いやられている。時には、これらが説明なしに割り引かれていることさえある(Reilly & Willenbockel, 2010; Wise, 2013)。
第三に、国連食糧農業機関の報告書では、規範的な言葉(「べきである」)に頻繁に移行し、また元に戻っていることは、制度的な選好が存在するにもかかわらず、うまく管理されていないことを示唆している。おそらく、「無関心」問題に対する最も有益な定式化は、(レトリックに過ぎないとしても)こう問うことであろう。もし、国連食糧農業機関のモデルが、提示された答えと二律背反する答え(すなわち、食糧供給は十分である)を出したら、国連食糧農業機関はそれを支持するだろうか。
5.3.1 食料危機の中での利害の対立
この疑問は,本章で明らかにした4つの疑問のある前提について最も顕著なこと,つまり,その影響が中立的でないことを考察することにつながる。それぞれが、現在あるいは将来の需要を誇張したり、供給を過小評価したりする結果となっている。したがって、これらはすべてマルサス的なシナリオを誇張することになる。Hertelが指摘した、国連食糧農業機関が生産に対する価格の影響を適切に考慮していないという仮定についても同じことが言える(Hertel, 2011)。これも将来の食糧供給を過小評価している。この影響は、最低でも何十億人もの人々が必要とする食糧に相当する。
マルサス的誇張の説明として考えられるのは、国連食糧農業機関が、食糧/学術/慈善事業の結びつきのほとんどすべての大規模機関と同様に、利害の対立を抱えていることだ。もし、ある種の食糧危機の脅威がなければ、国連食糧農業機関の組織的存在意義(モットー:Fiat Panis)は消滅してしまうだろう。さらに、このような対立がある以上、国連食糧農業機関の立場からすれば、ある種の食糧危機の方が他より明らかに望ましい。国連食糧農業機関、IFPRI、世界銀行といった機関が、農業の問題を土地の偏在、民主主義の欠陥、貧困、あるいはアグリビジネスの過剰な権力に起因するものとして捉えようとすれば、アグリビジネス、政府、裕福な土地所有者、まさに直接的、間接的にこれらの機関に出資、支配している人々と対立してしまうだろう。政治的にはるかに安全なのは、飢餓を生産不足のせいにすること、つまり農民のせいにすることである(Food First, 2016; Sampson, 2018)。
この文脈では、国連食糧農業機関自身のモデルによる、食糧生産の増加は飢餓に苦しむ人々の数にほとんど影響を与えないという、一見無視されたような予測を思い出すことが有用である(Alexandratos & Bruinsma, 2012; Bruinsma, 2003)。この予言は的中した。2018年、その後食糧生産は増加し、食糧価格は下落したにもかかわらず、栄養失調者の数は8億2100万人に増加した。国連食糧農業機関が飢餓を解決したいのであれば、彼ら自身のモデルは、生産量の増加以外のところに目を向けよと言っている。これはGAPSのような生産主義を前提としたモデルにとって、非常に有害な知見である。しかし、2003年から17年経った今でも国連食糧農業機関は生産性をターゲットにしており、ほぼすべての主要な食糧安全保障機関や慈善事業も同様である(Food First, 2016)。
5.3.2 最終的な考え
GAPS のようなモデルが生産性の問題を前提にすることで、特に利益を得ている利益団体がある。「世界を養う」ことは、国際的なアグリビジネスの主要な広報活動である。貧しい人々や飢餓に苦しむ人々を救うことができるのは、アグリビジネスだけである、というのが彼らの主張である(Peekhaus, 2010; Stone & Glover, 2011)。希少性の物語が真実であるならば、この主張は強力である。それは、農業を道徳的な問題へと変容させる(Dibden et al.2013 ; Latham, 2015)。農薬、遺伝子組換え生物(GMO)、単一栽培は否定的な結果をもたらすかもしれないが、それらは飢餓に代わる必要なものである、というのがこの物語の趣旨である。したがって、唯一の代替案は、特権階級のための贅沢品にすぎず、政策立案者の関心を引くことはない。あるいは、欠乏が神話であるとするならば、すべての農薬は散布され、すべての遺伝子組み換え作物は利益のためだけに存在する。主に農業のために行われる生態圏の破壊は、事実上無駄である(IAASTD, 2009; IPBES, 2019)。出る杭は打たれる。

アグリビジネスはこの点に立脚しているかどうか。その理由は、ジョージ・ラコフが論じたように、人間は道徳的な言葉で考え、行動するからである。彼らは自分たちを「良い」と思いたいのである(Lakoff, 2004)。したがって、道徳的本質を主張する生産性への定量的モデルの焦点は、アグリビジネスへの贈り物となるのである。
そのため、世界の農業需要と生産を推定・予測するために、GAPSやその他のモデルの作成に非常に大きな努力と注意が向けられてきたが、生産性の欠如が、それを刺激するとされる飢餓の蔓延に極めて重要な役割を果たすという明確な証拠は、事実上存在しない(Lappé & Collins, 2015; Sen, 1981)。対照的に、農業、特に工業的単一栽培は、生態系に優しく、私たちを養ってくれる個人とコミュニティにとってより有益になるように再編成する必要があるという証拠が豊富にあり、圧倒的でさえある(HLPE, 2013; IAASTD, 2009)。しかし、これは欠乏の物語を捨てない限り、実現しない。
別の道を歩む必要性は、国連食糧農業機関のモデラーもところどころで認めている。国連食糧農業機関 の最新の主要モデリング・レポート「食料と農業の未来」の序文には、次のように書かれている。2050年までの代替経路は、「長期的に食糧と農業システムの持続可能性を確保するためには、迅速かつ目的にかなった行動が必要である」(国連食糧農業機関, 2018)と述べている。しかし、その報告書でさえ、依然として希少性の物語を永続させている。それは、探索する代替経路の究極の決定者として、量的生産主義的な焦点を特権化し、それによって、そのようなすべての可能性を損なっている。国連食糧農業機関 やその他の機関が代わりに必要としているのは、農業と食糧システムの広範な結果、つまり多機能性に焦点を当てることだ。
多機能性とは、農業は他のシステムに深く組み込まれており、それゆえ農業がどのように行われるかは、作物生産量という単一の指標をはるかに超える結果をもたらすという考え方である(IAASTD, 2009; Kremen & Merenlender, 2018)。例えば、工業的農業は、気候変動やその他の大気汚染の唯一最大の原因となっている(Bauer et al., 2016; Goodland & Anhang, 2009; Steinfeld et al., 2006)。農業はまた、生活を提供し、水(帯水層、地表水、海洋)を浄化または汚染し、生物多様性を保全または劣化させ、景観価値と文化的に適切な食物を提供するなどしている(IUCN Task Force on Systemic Pesticides, 2017)。農業のこれらの貢献は、熱量生産の定量的モデルではすべて事実上無視されているが、生産性とは異なり、これらの機能はより危険にさらされることが多いという理由だけでも、意思決定者は少なくとも生産性と同じだけの配慮を受ける必要がある。
比較的単純なアウトプットである農業生産性のモデリングの限界について述べてきたが、世界レベルでの農業のモデリングが建設的な役割を果たすことができるかどうかは、それなりに疑問が残るところであろう。おそらく最終的には、モデリングのより有益な役割は、よりローカルなスケールで、より明確な性質の問題を問うモデリングになる。パリ流域の窒素モデリングに関する研究は、モデリングがより控えめな目的でありながら、政策立案者やその他の人々にとって有用であることを示す一例である(Billen et al.、2012)。国連食糧農業機関は今、農薬などの投入が生物多様性や人間の健康に及ぼす影響を、さまざまな農業システムから評価することに焦点を移すべきかもしれない。これは、現在の農薬規制が対応していない分野である(Vandenberg etal.) このような分析は、長い間無視されてきた外部性が今後中心的な役割を果たすようになることを、農業に知らせる有益な手段となるはずである。
13. 遺伝子編集作物やその他のGM作物は、持続可能な食糧システムを破綻させるだろうか?
アリソン・K・ウィルソンバイオサイエンス・リソース・プロジェクト(ニューヨーク州イサカ、米国
13.1 はじめに
従来の農業は、気候変動の主要な要因である(Foley et al., 2005)。天然資源、合成肥料、農薬の集中的な使用は、大気、土壌、水質をさらに悪化させ、大規模な生物多様性の損失を引き起こす(Foley et al., 2005; Horrigan, Lawrence, & Walker, 2002; Kremen & Merenlender, 2018; Pretty et al., 2000; Sánchez-Bayo & Wyckhuys, 2019; Tilman, 1998)。こうした評価により、従来の農業がより持続可能なものにならなければならないという幅広い合意が形成された。また、持続可能な農業への移行は、持続可能な食料システムの不可欠な要素である。しかし、食料安全保障の目標を達成しながら農業の持続可能性を向上させる方法については、非常に異なる見解がある(Godfray & Garnett, 2014; Holt- Giménez & Altieri, 2013; Kremen & Merenlender, 2018; McMichael & Schneider, 2011; Mercer, Perales, & Wainwright, 2012; Perfecto & Vandermeer, 2010; Zaidi et al., 2019)。
バイオ作物の持続可能性への影響は、依然として重要な論争分野である。良きにつけ悪しきにつけ、この影響は相当なものであると思われる。最もよく引用されるソースによると、2017年に遺伝子組み換え(GM)作物は24カ国の1億8980万haに植えられた(ISAAA, 2017b)。商業的に栽培されているGM作物の大半は、除草剤耐性(HT作物)か、昆虫病原体である細菌バチルス・チューリンゲンシスに由来するGM農薬を生産する(Bt作物)かのいずれかである。世界で栽培されている主なHT作物および/またはBt作物の商品作物は、ダイズ(9410万ha)、トウモロコシ(5970万ha)、綿(2410万ha)、カノーラ(1020万ha)である(ISAAA、2017)。
持続可能性の狭い視点は、農業の役割を食料安全保障の提供として捉えている。さらに、食料安全保障を高収量と同一視している(Latham, this volume)。その収量を得るために、従来の農業は、大規模なモノカルチャー作付けシステムと、農場外のインプット(ハイブリッド種子、農薬、水、燃料など)の多用とを組み合わせている。労働力は機械化によって代替される。土壌の肥沃度や害虫は、合成肥料、殺虫剤、殺菌剤、除草剤で管理される。
- a 慣習的農業(工業的農業とも呼ばれる)には、投入量の多い大規模なモノカルチャー商品作物栽培システム、閉鎖型動物飼育場が含まれる。
- b ISAAA(International Service for the Acquisition of Agri-biotech Applications)は、遺伝子組み換え作物を含む農業バイオテクノロジー製品の導入を促進するために、政府組織とバイオテクノロジー産業の両方から資金援助を受けている。
ハイブリッド種子は毎年購入する必要がある。その結果、持続可能性の向上は、単作物の収穫量/エーカーの増加や、それに伴う農地外投入資材の使用量の減少という枠組みになっている。バイオテクノロジー、特に遺伝子組み換え作物(遺伝子操作作物、バイオエンジニアリング作物とも呼ばれ、トランスジェニック、シスジェニック、RNAi、遺伝子編集作物など)の利用は、こうした持続可能性の目標達成に必要だと考えられている(Ammann 2005;Dibden、Gibbs、& Cocklin. 2013;Godfray & Garnett. 2014;Pretty 2001)。このパラダイムでは、収量を増やすことによって、GM作物は零細農家や持続可能な農業システムにも利益をもたらすことができると主張される(Holt-Giménez & Altieri, 2013; Mercer et al., 2012; Shelton, Hossain, Paranjape, & Azad, 2018).
これに対し、持続可能性の広義の視点は、農業を本質的に多機能であり、多様で相互に関連する環境および社会的影響を有すると見なす(McIntyre et al., 2009)。したがって、持続可能であるためには、農業は、(1) 食料安全保障と健康的な食生活を提供し、(2) 貧困と不平等を軽減しつつ、農村の生活と文化を支え、(3) 気候変動の軽減に重点を置いて、生物多様性と環境の健全性を高めなければならない(Horrigan et al., 2002; Kremen & Merenlender, 2018; McMichael & Schneider, 2011)。このパラダイムの中では、低投入の小規模農業、食料・種子主権、地域の食料システムを支援する技術や実践が不可欠と考えられている(Adhikari, 2014; Holt-Giménez & Altieri, 2013; Kremen & Merenlender, 2018; McIntyre et al., 2009)。しかし、持続可能な農業システムには、さまざまな可能性がある。これには、オーガニック、アグロエコロジー、アグロフォレストリー、伝統的なシステムなどがある。それらに共通するのは、複数の農業、環境、社会的目標を達成するために生物多様性を利用することである(Altieri, 1999; Thrupp, 2000; Wickson, Binimelis, & Herrero, 2016)。例えば、土壌の健全性と害虫の両方は、多品目輪作,カバークロップ,間作を含む実践,プッシュ/プル,SRIc,不耕起技術,家畜や木の取り込みによって、農場内の生物多様性を高めることで管理されている(Anderson, 2015; Hailu, Niassy, Zeyaur, Ochatum, & Subramanian, 2018; Kremen & Miles, 2012; Midega, Pittchar, Pickett, Hailu, & Khan, 2018; Pretty, 2001; Thakur, Uphoff, & Stoop, 2016; Zhang, Postma, York, & Lynch, 2014). このような技術は、農地外投入物の必要性を大幅に削減し、完全に排除することを目標とすることができる(Nicholls, Altieri, & Vazquez, 2016)。このような広範なアグロエコロジーの持続可能性のパラダイムの中で、GM作物はしばしば相容れないものとみなされる(Adhikari, 2014; Altieri, 2005; Barker, 2014; Fischer, 2016; Garibaldi et al., 2017; Kesavan & Swaminathan, 2018; Kremen & Miles, 2012; McIntyre et al., 2009; Pengue, 2005; Schütte et al., 2017; Wickson et al., 2016).
作物や栽培品種の幅広い多様性を利用できることは、持続可能な農業に不可欠な要素である。歴史的には、農民の種子保存と選択により、特性や適応性が大きく異なる作物品種の膨大な多様性が生み出された(Villa, Maxted, Scholten, & Ford-Lloyd, 2005)。植物育種の目的は、有益な新しい形質の組み合わせや特性を持つ作物を生産することで、この多様性に貢献することである。従来の植物育種では、性的に適合する植物を遺伝的に交配し、所望の形質を持つ子孫を選抜することでこれを実現している。
時には、従来の植物育種家は、意図的な突然変異誘発、体細胞突然変異、あるいは幅広い交配を利用して、新規形質を導入することもある(Wilson, Latham, & Steinbrecher, 2004)。一方、遺伝子組み換え作物の開発者は、実験室ベースの技術(例えば、組換えDNA(rDNA)技術、組織培養、植物形質転換(例えば、アグロバクテリウム感染または粒子砲撃)を組み合わせて使用し、新規形質を特定する特定のrDNA配列を作物植物に導入する(Barampuram & Zhang, 2011)。GM技術は、あらゆる生物(例えば、ウイルス、細菌、真菌、哺乳類、非食用植物)からDNAを作物のゲノムに結合、改変、および転送する能力を付与することにより、利用できる形質の範囲を拡大する(Wickson et al., 2016)。さまざまな「遺伝子編集」システムを含む一連の新しいGM技術(nGM)が、植物で使用するために開発されている(Eckerstorfer, Heissenberger, Reichenbecher, Steinbrecher, & Waßmann, 2019)。標準的なGM技術とは対照的に、遺伝子編集技術は、ゲノムの特定領域へのDNAの組み込みおよび/またはその他の改変を対象とすることができる(Fichtner, Castellanos, & Ülker, 2014)。GM および nGM 技術の利点は、(1) DNAの特定の配列を移植または変更する能力、および (2) 通常の植物育種では導入できない新規のDNA 改変および形質を導入する能力にあると主張されている。
13.2 HT作物とBt作物の影響
理論的には、HTd作物とBte作物は、(1) 農薬の使用全体を減らし、(2) 安全な農薬(例えば、植物が生産するBt毒素やグリホサート除草剤)をより有害な農薬に置き換えることで持続可能な農業を促進することを目的としていた(Ammann 2005;Andow. 2010;Koch et al., 2015). HT作物は、不耕起農業の導入を促進することで、持続可能性を向上させるとも言われている。
しかし実際には、Bt作物やHT作物の普及により、害虫抵抗性、「スーパーウィード」、二次害虫の発生が問題となっている(Benbrook, 2018; Bonny, 2016; Carrière et al., 2016; García et al., 2019; Gould, Brown, & Kuzma, 2018; Kilman, 2010; Kranthi, 2016; Mortensen, Egan, Maxwell, Ryan, & Smith, 2012; Stone & Flachs, 2018; Tabashnik & Carrière, 2017)。これらの問題に対して、農家は殺虫剤と除草剤の両方の使用を増やした。また、耕起やその他の機械的な方法で雑草を駆除することを増やしたところもあった(Bonny, 2016; Green, 2014)。種子業界の対応としては、各品種に複数のBt農薬および/またはHT形質(スタック形質およびピラミッド形質f)を追加し、新しい植物産生農薬(VIPタンパク質毒素やRNAiベースの殺虫剤など)を開発した(Bøhn & Lövei, 2017; Carrière et al, 2016; Chakroun, Banyuls, Bel, Escriche, & Ferré, 2016; Gould et al., 2018).
さらに、BtやHT作物とそれに付随する農薬の導入は、農家の実践に様々な変化を促し、それ自体が非常に有害な環境影響を及ぼしてきた。まず、Bt作物やHT作物の導入は、総合的病害虫管理(Gray, 2010)や持続可能な技術の利用を損なってきた。それらは、GM作物が生産する農薬や化学農薬を、耕起、短期作物、カバークロップ、輪作、生物的防除を含む農薬を使わない防除手段で代用することによって行う(Brainard, Haramoto, Williams, & Mirsky, 2013; Gutierrez, Ponti, Herren, Baumgärtner, & Kenmore, 2015; Kesavan & Malarvannan, 2010; LaCanne & Lundgren, 2018; Lang, Oehen, Ross, Bieri, & Steinbrich, 2015; Schutte et al. 2017; Stone & Flachs, 2018; Tooker, 2015)。第二に、Bt作物の栽培(実際の昆虫の圧力が判明する前の決定)は、対象となる害虫が脅威だろうかどうか、Bt毒素が有効な保護を提供するかどうかにかかわらず、景観、および消費者を農薬にさらす。したがって、Bt作物の幅広い取り込みは、予防的な農薬使用である(Douglas & Tooker, 2015; Gray, 2010; Stone & Flachs, 2018; Tooker, 2015)。最後に、HT作物は、簡略化された作物ローテーションおよび/または農業システムを奨励することによって、生物多様性を減少させてきた。また、栽培期間中、広範囲な除草剤の無制限散布を許可し、生物多様性の損失をさらに悪化させている(Schreiner, 2009; Schütte et al., 2017)。もう一つの結果として、米国市場のHT大豆はグリホサート汚染が高い(Bøhn et al., 2014)。
このようにBtおよびHT形質は、農薬の踏み絵を悪化させ、拡大してきた(Altieri, 2000; Binimelis, Pengue, & Monterroso, 2009; Douglas & Tooker, 2015; Mortensen et al., 2012; Pengue, 2005; Stone & Flachs, 2018)。結果として生じる「技術促進型農薬トレッドミル」については、Douglas and Tooker (2015)が説明している:
ネオニコチノイド種子処理剤は、農家にとって魅力的な他の技術とともに「タグ・オン」された可能性もある。ネオニコチノイド種子処理剤は、通常、より大きなパッケージの一要素であり、例えばトウモロコシでは、生殖質(すなわち作物品種)、最大8つのトランスジェニック、最大6種類以上の種子処理(殺菌剤、殺線虫剤、殺虫剤)を含むことができる。
これらのよく知られた結果は、HTおよびBt作物の採用が、ネオニコチノイド、グルホシネート、2,4-d、ジカンバといった極めて毒性の高いことが知られている農薬の使用を含む、長期にわたる農薬使用の劇的な増加につながることを示している(Douglas & Tooker, 2015; Mortensen et al, 2012; Schütte et al, 2017; Tooker, 2015)。HT作物による除草剤使用の増加は、科学文献で繰り返し実証されている(Schütte et al., 2017)。しかし、Bt作物は農薬の使用を減らすことができると主張する著者もいる(例えば、Klümper & Qaim, 2014; Naranjo, 2009)。短期間の研究(例えば、抵抗性が発現する前)、あるいは植物が生産したBt毒素および/または種皮殺虫剤の量を定量化しなかったことが、これらの矛盾を説明できる(Benbrook. 2012;Douglas. 2016)。適用農薬、GM作物生産殺虫剤(例えば、Benbrook, 2012; Clark, Phillips, & Coats, 2005; Nguyen & Jehle, 2007; Saxena, Stewart, Altosaar, Shu, & Stotzky, 2004; 米国環境保護庁、2010; van der Hoeven, 2014)、および種皮農薬を完全に考慮すると、BtおよびHT作物はいずれも農業システムでの農薬使用量が増える(例えば、、Benbrook, 2012; Bøhn & Lövei, 2017; Bonny, 2016; Capellesso, Cazella, Schmitt Filho, Farley, & Martins, 2016; Douglas & Tooker, 2015; Heinemann, Massaro, Coray, Agapito- Tenfen, & Wen, 2014; Kranthi, 2016; Perry, Ciliberto, Hennessy, & Moschini, 2016; Yang, Iles, Yan, & Jolliffe, 2005).
13.2.1 GM作物に関連する農薬の毒性
GM作物の開発者と米国の規制当局は、Bt毒素とグリホサートは低毒性の農薬だと主張している(Koch et al., 2015; Williams, Kroes, & Munro, 2000)。しかし、Bt作物については、Bt毒素やBt植物が哺乳類、益虫、水生無脊椎動物などに対して有害な標的外影響を及ぼすことを示す証拠が増え続けている(Andreassen et al., 2015; Hilbeck & Schmidt, 2006; Latham et al., 2017; Paula et al., 2014; Venter & Bøhn, 2016)。多くの研究者が、さらなるバイオセーフティ研究、特に植物体研究、Bt作物への曝露による亜致死影響および長期影響に関する研究の必要性を指摘している(例えば、Andow, 2010; Arpaia et al., 2017; Hilbeck & Otto, 2015; Latham et al., 2017; Sanchis, 2011; Wolfenbarger, Naranjo, Lundgren, Bitzer, & Watrud, 2008)。同様の懸念は、グリホサートベースの除草剤にも当てはまる。例えば、グリホサートおよび/またはその製剤は、土壌や腸内細菌叢の組成に影響を与え、ミミズ、益虫、水生生物に悪影響を及ぼす(Schütte et al., 2017; Sharma, ジャー, & Reddy, 2018)。また、ヒトのがんや慢性腎臓病にも関連している(例:Jayasumana, Gunatilake, & Senanayake, 2014; McHenry, 2018; Myers et al., 2016)。それらの有害なオフターゲット影響を文書化した多くの証拠により、重大な研究ギャップと相まって、Bt毒素とグリホサート系除草剤が低毒性農薬であるという科学的合意はない(Ardekani & Shirzad, 2019; Hilbeck et al., 2015; Krimsky, 2015)。
13.3 GM作物における意図しない形質(unintended traits/UT)
意図した形質とは関係なく、GM技術はその精密さでしばしば称賛される。特に、従来の植物育種を複雑にする問題である「イールドドラッグ」の問題なしに、新規形質を導入できることが挙げられる(Gepts, 2002)。しかし、このような主張とは裏腹に、GM作物における予期せぬ有害な意図しない形質(UT)の報告が、定期的にメディアで取り上げられる。例えば、2012年、ウォール・ストリート・ジャーナル紙を含む様々な報道機関が、GMトウモロコシと大豆の茎が従来の作物よりはるかに丈夫であると報じた(Tita, 2012)。

丈夫なGMの茎は、トラクターのタイヤをパンクさせる。この予想外の特性は、農家が高価な強化トラクターのタイヤを新たに購入したり、タイヤを頻繁に交換したりしなければならなくなるため、経済的・環境的コストがかかる。このような報告の多くがそうであるように、「強靭な茎」の形質は、科学的な文献で十分に追跡調査されたり、認められたりしていない。しかし、この報告は、次のような重要かつ無視されがちな問題を提起している。
- (1) GM技術はどれほど精密で予測可能なのか?
- (2) 開発者や規制当局は、有害なUTを持つGM作物が市場に出回るのを防いでいるのか?
- (3) UTは持続可能性にとって過小評価されている障害なのか?
UTとは、意図しない効果と呼ばれることもあるが、ここでは、GM作物と非GMの同系統の作物との間で、意図したGM形質以外の有意な差異があるものと定義する。したがって、UTには、例えば、種子の発芽、雑草の抑制、害虫抵抗性、乾燥耐性、高さ、収量、開花時期などの特性における統計的に有意な差異が含まれる。さらに、UTには、栄養素、毒素、その他の生化学物質における組成の違いも含まれる。このようなUTは、トランスクリプトーム、プロテオーム、メタボロームプロファイリング研究によって明らかになることが多い(Cellini et al., 2004)。複数の査読者が、科学文献に記録された多種多様なUTの例を集めている(Cellini et al., 2004; Haslberger, 2003; Kuiper, Kok, & Engel, 2003; Nature Institute, 2019; Ricroch, Bergé, & Kuntz, 2011)。とはいえ、文書化された例はすでに収集したものよりはるかに多く、包括的なレビューを行うことはできない。
13.3.1 精度と予測可能性
遺伝子組み換え技術が精密で予測可能であれば、バイオテクノロジストは1つの遺伝子組み換え植物を作ればよく、その植物は意図した新しい形質を除いて親植物と同一であるはずだ。しかし、バイオテクノロジーの専門家たちは、何百、何千もの初期形質転換体を作り出す。同時に、どの遺伝子が小麦に最も効果的だろうかが明確でなかったため、多くの異なる遺伝子を試験した。最初の集団から1300以上のグリホサート耐性植物を選び出し、残りの植物は廃棄した。その後の選抜では、ラウンドアップ耐性と基本的な農作業能力を評価した。このような選抜を4世代続けた結果、6つの品種が残った。そして、3年間の「大規模圃場試験」を経て、ようやく1品種が商品化され、米国農務省(USDA)に提出され、承認された。この品種はその後、規制緩和の請願が取り下げられた。
Btイネ(Shu et al., 2002)、Btまたは病害抵抗性ジャガイモ(Davidson et al., 2004; Felcher, Douches, Kirk, Hammerschmidt, & Li, 2003)、ウイルス耐性タバコ(Xu, Collins, Hunt, & Nielsen, 1999)、ウイルス耐性大麦(Bregitzer、Halbert、& Lemaux, 1998)の開発で作られた初期形質転換体の大規模集団に関する研究により、他の遺伝子組み換え作物と形質に同様の問題があることが指摘された。収量、高さ、茎、葉の形態などの基本的な農学的形質の欠陥は、再生したGM植物に頻繁に見られ、多くの初期形質転換体が複数のUTを示す。数種類(2~22種)の形質しか評価されない場合でも、初期形質転換体の比率は、UTを持つ形質転換体とは、外来DNAが導入された細胞または植物などの生物を指すがUTを持つ割合は、通常20%からほぼ100%の間である(例えば、Bregitzerら, 1998; Dale & McPartlan, 1992; Davidsonら, 2004; Felcherら, 2003; Hoekema, Huisman, Molendijk, van den Elzen, & Cornelissen, 1989; Kumar, Rakow, & Downey, 1998; Shu et al., 2002; Vickers, Grof, Bonnett, Jackson, & Morgan, 2005)。
遺伝子組換え作物開発の後半では、何度も選抜を繰り返してもなお、多様なUTが頻繁に確認される。イネをサンプル作物として取り上げると、収量、種子サイズ、または活力の低下がすべて異なるBtイネ系統で報告されている(例えば、Bashir et al., 2004;Chen、Snow、Wang、& Lu 2006;Jiang et al., 2018;Shu et al., 2002;Tu et al., 2000;Wangら、、2012; Wei-xiang, Qing-fu, Hang, Xue-jun, & Wen-ming, 2004; Wu, Shu, Wang, Cui, & Xia, 2002)、穀物とわらの品質への変化(例えば、Bashir et al., 2004; Li et al., 2008; Wei-xiang et al., 2004;Wu et al., 2002)などが挙げられる。また、グルホシネート耐性イネの系統では、高さ、収量、発育のUTが記録されている(Oard et al., 1996)。
その他のUTの例は、本総説の表 13.1 および表 13.2にまとめられている。これらは、他の査読者が集めた例と合わせて、UTが特定のGM技術、形質、または植物種に限定されないことを確認している(Cellini et al., 2004; Haslberger, 2003; Kuiper et al., 2003; Nature Institute, 2019; Ricroch et al.)
13.3.2 商業用GM作物におけるUT
商業用GM作物は、GM植物育種が提供できる「最高の中の最高」と考えられている。それらは、「厳格な成分、栄養、安全性評価」、場合によっては環境リスク評価(Larkin & Harrigan, 2007)に先立ち、何年もの選択と開発を経る。最後に、現在栽培されているほとんどの遺伝子組み換え作物は、商業的な発売や輸入が許可される前に、何らかの規制プロセスを経ている(Davison, 2010; Pelletier, 2005)。とはいえ、表13.1には、商業用GM作物で確認されている多くのUTのいくつかを例示している。これらのUTは、査読付き論文や、米国で特定の作物や事象を規制緩和するためにUSDA/APHIS(米国農務省/動物・植物衛生検査局)に提出した請願書に記録されている。表13.1に示した例は、持続可能性に影響を与えるものであるため、選択されたものである。
表13 1からは重要な結論が導き出される
まず、UTを持つ商業作物の例を見つけるのは難しくない。実際、多くの商業用遺伝子組み換え作物には複数のUTが存在する。例えば、Mon810トウモロコシについて記録されたUTには、多数の組成の違い(リグニンの増加やアレルゲンの存在を含む)、水分含量の増加、有益な土壌生物への悪影響が含まれる。第二に、UTにはさまざまな起源がある。cry1Ab Btトウモロコシに関連するリグニンレベルの増加のようなものは、多くの独立したcry1Ab事象で見られるように、導入遺伝子(すなわち形質自体)の多面的効果であると考えられる。また、Btペイマスターコットンにおける根瘤線虫に対する抵抗性の喪失のように、事象に特異的で、導入遺伝子の挿入部位またはその近傍での突然変異によるものと考えられるものもある(Colyer et al., 2000)。第三に、商業用遺伝子組み換え作物で一般的に記録されている多くのUTは、持続可能性に明白な悪影響を及ぼしている。例えば、MON810やBt11 トウモロコシで記録されているように、水分含有率が高いため、乾燥時に追加エネルギーが必要となる(Ma & Subedi, 2005)。害虫抵抗性の喪失や収穫可能量の減少などのUT は、特に農家が収量を維持するために外部からの投入物(例:燃料、農薬、肥料、水)を多く使用する場合に、持続可能性を損なう。表 13.1には、RR 大豆(7%~11%減)、グリホサート HT トウモロコシ(5.98 ブッシェル/エーカー減)、トウモロコシ根こぶ病予防 Bt トウモロコシ(12.22 ブッシェル/エーカー減)について記録された大きな収量減少を含む(ゴードン 2007;シー他、2013)。表13.1に記載されている害虫や病原菌に対する抵抗性の喪失には、Btトウモロコシ品種におけるアブラムシやアザミウマの数の増加、アトランティックニューリーフBtジャガイモとペイマスターBtワタにおける線虫抵抗性の喪失、様々なBtワタ品種で観察されたフザリウム菌類病に対する抵抗性の減少がある(Bourguet et al., 2002; Brodie 2003; Colyer et al., 2000; Faria et al., 2007; Li et al., 2009)。表 13.1 は、商業用遺伝子組み換え作物(多くは世界中で数百万エーカーで栽培)が、HTやBt 作物の「丈夫な茎」に加えて、有害なUTを頻繁に持ち、その大規模かつ負の環境影響を助長するUTを持つことを示すものである。
それにもかかわらず、多くのバイオテクノロジストたちは、商業品種においてUTが確認されても、重要でないとして却下する傾向がある(例えば、Fox, Morrison-Saunders, & Katscherian, 2006; Larkin & Harrigan, 2007; Ricroch et al., 2011; Shepherd, McNicol, Razzo, Taylor, & Davies, 2006; Sidhu et al., 2000)。これは、RR 大豆について記録された収量損失 UTのケースである。RR 大豆の開発者は、米国農務省への規制に関する請願の中で、さらなる品種改良によって収量を向上させると主張していたが、それが実現することはなかった(USDA/APHIS、Petition P93-258-01, 2020)。同様に、CZW3カボチャの大きく統計的に有意な組成UTも、さらなるリスク評価の必要性を示す赤旗として、あるいはその特定の事象や作物を拒否するために解釈されるのではなく、却下された(USDA/APHIS Petition 95-352-01p )。しかし、代謝物、栄養素、または潜在的な毒素に対する定量的または定性的な変化などの大小の組成上のUTは、生態学的相互作用(Arpaia et al., 2017; Li et al., 2009; Mesnage et al., 2016; Venter & Bøhn, 2016)や食品または飼料安全(Haslberger, 2003; Pelletier, 2005; Schubert, 2008)といった持続性に必須の特質に対し負の影響を与えることがある。また、ある環境下またはある野外試験で見られた統計的に有意な差異が、他の環境では見られないということも日常的に行われている。しかし、導入遺伝子は、導入遺伝子×環境の相互作用が大きい場合がある(Zeller, Kalinina, Brunner, Keller, & Schmid, 2010)。したがって、特定の条件下でのみ見られるUTは、環境応答の欠陥やその他の重大な問題を示す可能性もあるため、よりリスク評価の出発点として考慮する必要がある(Agapito-Tenfen、Guerra、Wikmark、& Nodari. 2013)。
表13 1の例は、商業用GM作物で記録されたUTを網羅したものではないことを強調することが重要である
さらに多くの要因が重なり、公開されているデータは、UTを持つ商業系統の数と各商業系統のUTの数を著しく過小評価している可能性がある。アウトクロッシングの増加、種子休眠性、シードバンクの持続性など、多くの重要な持続可能性形質は、悪影響を及ぼす可能性があるにもかかわらず、評価されることはないかもしれない(例えば、Altieri, 2005; Bergelson, Purrington, & Wichmann, 1998; Linder, 1998; Linder & Schmitt, 1995)。
GPT-4:
アウトクロッシング:
アウトクロッシングとは、植物の交配方式の一つで、異なる個体間で遺伝子が交換されるプロセスを指す。これに対して、自家受粉(セルフィング)は、同じ個体内での交配を指す。アウトクロッシングは、遺伝的多様性を高め、植物の適応能力や持続可能性を向上させることができる。しかし、一方で、アウトクロッシングが増加すると、遺伝子プールに不適合な形質が広がることがあり、悪影響を及ぼす可能性もある。
シードバンクの持続性:
シードバンクとは、土壌中に存在する種子の貯蔵庫を指す。これらの種子は、環境条件が適切になるまで休眠状態で待機し、条件が整うと発芽する。シードバンクの持続性は、植物が長期間にわたって環境変化に対処し、生存戦略を維持する能力を指す。シードバンクの持続性が高いと、植物個体群は不適な環境条件による影響を受けにくくなる。ただし、シードバンクの持続性が高すぎると、一時的な環境変化に適応する機会を逃すことがある。
商業用GM作物に関するその他の制限としては、組成研究の標準化の欠如、ゲノム分析の欠如、多くのラボおよびフィールド試験における不十分または不適切な試験条件の使用(例:毒性試験を行う生物またはライフステージが不適切、フィールドプロット間の距離が不適切)、GM作物と製品の長期および亜致死影響に関する研究の欠如が挙げられる(Arpaia et al., 2017; Booij, 2014; Hilbeck, Meier, & Trtikova, 2012; Pelletier, 2005; Schubert, 2008).
さらに、GM作物の所有権は、独立した研究を制限する作用がある(Waltz, 2009)。大学や資金提供者も、GM作物による害を発見する可能性のある研究を奨励せず、害の発見はしばしば激しく論争されるか、あるいは抑制される(Fagan et al., 2015; Peekhaus, 2010; Waltz, 2009)。その結果、商業化されたほとんどのGM作物は、UTを特定できる独立した試験やリスク評価をほとんど受けていない(Diels, Cunha, Manaia, Sabugosa-Madeira, & Silva, 2011; Séralini, Mesnage, Defarge, & de Vendômois, 2014).
要約すると、市販の遺伝子組み換え品種を含め、標準的な遺伝子組み換え技術を用いて開発されたすべての作物において、ユビキタスではないにしても、UTは頻繁に発生している。これらのUTは持続可能性に悪影響を及ぼすことが多く、GM作物は持続可能な農業と食料システムにとって不適切な選択となっている(Kesavan & Swaminathan, 2018)。
13.3.3 標準的なGM 技術がUTに寄与する
UT は、導入遺伝子(または付随する選択可能マーカー遺伝子)の意図しない影響から生じることがある。しかし、GM作物を生産するために使用される技術もUTを生じさせる可能性がある(Wilson, Latham, & Steinbrecher, 2006)。
導入遺伝子は、通常アグロバクテリウムの感染によって植物細胞に導入されるか、あるいは「遺伝子銃」を使ってDNAでコーティングした粒子を植物細胞に浴びせる。導入遺伝子はその後、植物の自然なDNA修復機構によって、植物ゲノムの損傷領域に統合される。したがって、導入遺伝子が組み込まれるゲノム上の位置は制御不能であり、独立した組み込みイベントごとに異なる。改変された植物細胞は、組織培養によって植物全体に再生される。1つ以上のトランスジェニックイベントを持つ再生植物は、さらなる分析のために選択される(多くの場合、抗生物質または除草剤耐性を指定する共移植マーカー遺伝子の助けによって)。このように、トランスジーンの挿入は、必然的に内在性の植物ゲノムを破壊する。
さらに、アグロバクテリウムの感染、粒子砲撃、組織培養は、いずれも高い変異原性を持つことが示されている。これらを合わせると、何千もの変異が生じる可能性がある。これらの変異は、導入遺伝子が組み込まれた部位に存在することもあれば、それに連なることもあり、またゲノム全体に広がることもある(Wilson et al., 2006)。このような変異には、塩基対の変化、大小のDNAの挿入と欠失、大規模なゲノム再編成のほか、細菌染色体DNA、ベクターDNA、複数の導入遺伝子、導入遺伝子断片の意図しない統合がある。このように、GM技術の変異原性は、GM作物で記録されたUTの頻度と多様性に寄与している(Wilson et al., 2006)。場合によっては、遺伝子組換え(アウトクロッシングまたはバッククロッシング)によってUTを除去することができる。しかし、導入遺伝子の挿入部位と遺伝的に結びついたUTは、目的の形質から切り離すことは不可能ではないにせよ、困難であろう。UTは、クローン的に増殖する、あるいは交配が困難または不可能な商業作物にとって、さらに大きな問題となる。これにはジャガイモ、バナナ、キャッサバ、そしてほとんどの樹木作物が含まれる。
13.4 新しい遺伝子組み換え形質と技術
前のセクションで述べたUTは、害虫抵抗性、ウイルス抵抗性、除草剤耐性という非常に限られた形質のために作られた遺伝子組み換え植物で確認されたものである。これらは単純な形質であり、単一の導入遺伝子によって特定され、その新規生成物は通常の植物機能、構造、または生化学的経路を変更することは意図されていない。乾燥、塩分、熱、または洪水に対する耐性の向上、特定の栄養素のレベルの意図的な変更、または収量の増加など、持続可能な農業に利益をもたらす可能性のある複雑な形質については、UTはさらに大きな障害になると考えられる(例えば、Flowers & Yeo, 1995; Kollist et al, 2019)。表13.2は、このような複雑な形質を持つGM作物で、UTを示す多くの文書化された例の一部を列挙したものである。
13.4.1 複雑なGM形質には失敗の歴史がある
メディアから頻繁に好意的に注目されているにもかかわらず、複雑なGM形質を持つ商業用GM作物を開発する試みのほとんどは失敗している。せいぜい、実行可能な製品を生産する上で、従来の植物育種に大きく遅れをとった程度である(例えば、Barker, 2014; Gilbert, 2016; McFadden, Smith, Wechsler, & Wallander, 2019; Stone & Glover, 2011, 2017)。
複雑な形質を持つ極めて限られた数のGM作物が商業的に利用できるようになったのは、ごく最近のことである(例:McFadden et al., 2019; Waltz, 2015)。しかし、UTのリスクが高いにもかかわらず、これらの新しいGM作物は、これまでの商業化されたGM作物よりもさらに独立した研究、リスク評価、規制の精査を受けていない(Camacho, Van Deynze, Chi-Ham, & Bennett, 2014; Waltz, 2016, 2018). 実際、「干ばつに強い」ダイズHB4については、開発者自身がその形質の背後にある作用メカニズムを理解していないと認めている(Waltz, 2015)。
13.4.2 ゴールデンライスの事例
ゴールデンライスは、栄養強化型GM作物の例として広く引用されている(Bollinedi et al., 2014)。珍しいことに、ゴールデンライスに関するかなりの量の研究が科学文献に掲載されている。したがって、ゴールデンライスは、理論的に持続可能な農業に利益をもたらす可能性のあるGM形質のケーススタディとして有用である。

ゴールデンライス品種は、β-カロテン生合成経路の酵素を指定する2つの導入遺伝子を含んでいる(Bollinedi et al., 2014; Ye et al., 2000)。理論的には、米の胚乳でこれらの酵素を標的として生産することで、β-カロテン(プロビタミンA)の穀物レベルを高めることができる(Dubock, 2014)。
2000年に最初のゴールデンライス論文が発表されて以来、官民の研究者がゴールデンライスを何度も繰り返し生産し、それぞれが米粒中のβ-カロテンのレベルをさらに高めることを目的としている(Bollinedi et al., 2014)。シンジェンタは、公共部門が使用するために、GR2の6つの事象を寄贈した(Bollinedi et al.) 国際稲研究所(IRRI)は、ビタミンA欠乏症の人口が多いとみなされる国を対象とした育種活動でこれらを使用した(Bollinedi et al., 2014; IRRI, 2019; Stone & Glover, 2017)。2つのゴールデンライス事象、GR2-R1およびGR2Eは、最も多くの研究開発の対象となっている(例えば、Bollinedi et al., 2017,2019;Paine et al., 2005;Schaub et al., 2017)。
13.4.2.1 GR2-R1
イベントは、劇的な収量低下を含む農学的欠陥を引き起こす長年にわたり、GR2-R1イベントはゴールデンライス育種努力の中心であった(Bollinedi et al., 2017; Stone & Glover, 2017)。しかし、GR2-R1系統は、一貫して低い収量を与えた(Dubock, 2014; Stone & Glover, 2017)。さらに、インドの研究者は、GR2-R1において、小人症、多毛、淡い緑色の葉、根の欠陥、遅い開花、および低い稔性を含む他のUTを記録した(Bollinedi et al., 2017)。
GR2-R1イネで観察されるUTには、少なくとも2つの根本的な欠陥が寄与している。1つ目は、導入されたDNAそのものに関連するものである。GR2-R1植物では、導入遺伝子によって指定された酵素が、穀物とは別の組織で活性化する(Bollinedi et al., 2017)。これは、少なくともGR2-R1では、GR2導入遺伝子制御配列が意図したとおりに機能しないことを示している(Paine et al., 2005)。2つ目の欠陥は、インドの研究者がGR2-R1の統合部位を配列決定した際に発見された。GR2-R1では、統合された導入遺伝子は、オーキシン輸送タンパク質を規定するOSAux1と呼ばれるネイティブ遺伝子を破壊している(Bollinedi et al., 2017)。オーキシンは、成長と行動に重要な機能を持つ植物ホルモンである。
制御欠損と挿入部位欠損の両方が、植物の成長と発達に重要な役割を持つ、いくつかの追加の植物ホルモンに影響を与えると予測されている。アブシジン酸、ジベレリン、サイトカイニンなどである。実際、研究者らは、GR2-R1イネの葉、茎、開花部において、非GMの同系統と比較して、これら3つのホルモンのレベルが変化していることを発見した(Bollinedi et al., 2017)。多くのUTと固有の分子欠陥を考慮し、GR2-R1をさらに開発する努力は放棄された。
13.4.2.2 GR2E:穀物中のβ-カロテンのレベルが低い。
GR2-R1の欠陥が明らかになるにつれて、2番目の事象であるGR2EがIRRIのゴールデンライス育種プログラムに組み込まれた(Dubock. 2014)。しかし、ゴールデンライス形質がプロビタミンAを生産する効果は、イベントによって大きく異なる(Bollinedi et al., 2019; Paine et al., 2005)。シンジェンタの6つのGR2イベントのうち、GR2Eはβ-カロテンレベルが最も低い(Bollinedi et al., 2014; Paine et al., 2005)。
科学的および規制的な文献には他の、時にはより高い測定値が存在するが(Bollinedi et al., 2019; FSANZ, 2017; Paine et al., 2005; Schaub et al., 2017)、世界中の規制当局に提出したデータでは、GR2E米のβ-カロテンレベルは精米時わずか 3.5 μg/g、無洗米時 0.5-2.35 μg/gとされていた。(FDA, 2018a, 2018b).
13.4.2.3 ゴールデンライス:β-カロテンは貯蔵中に急速に劣化する
2つの異なる研究グループは、GR2E米穀のβ-カロテンレベルが貯蔵中に急速に減少することを報告している(Bollinedi et al., 2019;Schaub et al., 2017)。3週間の保管後、ゴールデンライスGR2Eは元のレベルの60%しか保持していなかった。10週間後には13%しか残っていなかった(Schaub et al., 2017)。2つ目の論文では、今度はGR2EとGR2-R1の両方について同様の結果が報告された(Bollinedi et al., 2019)。GR2-R1については、いくつかの異なる遺伝的背景で急速な分解が起こることが示された。調理は、β-カロテンレベルをさらに低下させることが示された(Bollinedi et al., 2019)。これらの結果を合わせると、通常の保管および調理条件下でのβ-カロテンの急速な分解は、ゴールデンライス品種の一般的な問題であることが示唆される。
13.4.2.4 ゴールデンライス: 利益とリスク評価が欠けているにもかかわらず商業化?
ゴールデンライスにおける農学的およびバイオセーフティ的なUTは、この形質の2つの側面から生じると考えられる。第一に、GR2-R1 で示されたように、β-カロテン生合成経路と交差する多くの生合成経路の意図しない改変から UTが発生する可能性がある。これらの交差する経路は、植物ホルモンに加えて、揮発性物質、他のカロテノイド、未知のシグナル伝達分子など、さまざまな化合物を生成する(DellaPenna & Pogson, 2006)。このような穀物中のUTは、例えば、栄養、毒性、種子休眠、発芽、稔性に影響を与えるかもしれない。
第二に、穀物中のβ-カロテンの急速な分解は、分解産物のレベルとバイオセーフティについて疑問を投げかける(Schaub et al., 2017)。
しかし、UTの可能性が高いにもかかわらず、現在、GR2Eに関するゲノムデータまたはその他の適用可能な研究が全くない。さらに、対象となる集団について、重要なヒトでの有効性および安全性の研究がまだ不足している(Schubert, 2008; Stone & Glover, 2017; Then & Bauer-Panskus, 2018)。しかし、オーストラリア、米国、カナダの規制当局は、GR2Eに関する開発者のバイオセーフティ主張を受け入れている(IRRI、2019)。
したがって、ゴールデンライスに関するデータは、ゴールデンライスの継続的な失敗の原因は評論家と過剰規制であるという推進派の主張と矛盾する(Dubock, 2014; Lee & Krimsky, 2016; Stone & Glover, 2017)。むしろ、入手可能なデータは、ゴールデンライスの商業化が、GM植物育種に固有の技術的な困難によって一貫して妨げられてきたことを示唆している。さらに、現在の有力候補であるGR2Eは、対象集団のビタミンA欠乏症の緩和を支援するという表明された人道的目標に有用な貢献をする可能性は低い。これは、GR2Eの穀物中のβ-カロテンの初期レベルが低く、その後急速に分解されることが一因である。しかし、GR2Eを商業化することは、食糧システムに不必要な農学的リスクとバイオセーフティリスクをもたらすことになる。β-カロテンの分解を遅らせることが示唆されているゴールデンライスの真空包装は、食品システムの持続可能性をさらに損なうことになる(Bollinedi et al., 2019)。
13.4.2.5 ゴールデンライス:
GM植物育種の失敗を照らす複雑なGM形質であるゴールデンライスの開発は、すべてのGM植物育種家が直面する固有の技術的課題の多くを例示している。これには、(1) GM形質を導入するために使用される技術の不正確さと変異原性、(2) 導入遺伝子とゲノムの構造および機能との関係の基礎となる生物学的プロセスの不十分な科学的理解、(3) 遺伝子と形質の関係およびそれらが発生および/または環境要因によってどのように影響されるかの限られた科学的理解、がある。
これらの技術的な困難が重なり、遺伝子組み換え植物の育種はUTに陥りやすく、最終的には失敗に終わる。ゴールデンライス開発の歴史は、持続可能な農業に役立つ可能性の高い複雑な遺伝子組み換え形質をうまく表現した安全で丈夫な作物品種の生産が、さらに問題である可能性が高いことを示唆している。
ゴールデンライスはさらに、GM作物に対する適切なリスク評価と規制を実施するための規制当局の制度的失敗を全体的に浮き彫りにしている。多くの研究者がすでに指摘しているように、より厳格な規制が必要である(1) GM形質や作物がその目的を確実に果たすため、また(2) 食料システムと環境を保護するため(Fox et al. 2006; Freese & Schubert, 2004; Heinemann, Agapito-Tenfen, & Carman, 2013; Heinemann, Kurenbach, & Quist, 2011; Hilbeck et al., 2015; Hilbeck & Otto, 2015; Latham et al., 2017; Mandel, 2003; Modonesi & Gusmeroli, 2018; Pelletier, 2005, 2006; Schubert, 2008; Venter & Bøhm, 2016; Wilson et al, 2006).
13.4.3 新しい遺伝子組み換え技術はより正確か?
バイオテクノロジストたちは現在、新世代のゲノム改変技術(nGM)が食料生産を持続的に増加させるために不可欠であると主張している(Stone, 2017; Zaidi et al., 2019)。これらのnGMには、シスジェネシス/イントラジェネシスとRNAiが含まれるi。イントラジェニック、シスジェニック、およびRNAiベースの形質から生じる特定のリスクについては、読者は他のレビュー(Casacuberta et al., 2015; Eckerstorfer et al, 2019; Gelinsky & Hilbeck, 2018; Heinemann et al, 2013; Lundgren & Duan, 2013; Senthil-Kumar & Mysore, 2011)に言及すること。
最近開発された植物用のnGMで、圧倒的に議論されているのは、「遺伝子編集」と呼ばれる技術である(Agapito-Tenfen, Okoli, Bernstein, Wikmark, & Myhr, 2018; Casacuberta et al., 2015; Eckerstorfer et al, 2019; Hou, Atlihan, & Lu, 2014; Lusser & Davies, 2013)。遺伝子編集は、オリゴヌクレオチド指向性突然変異誘発(ODM)、jおよび/またはメガヌクレアーゼ、TALEN、k ZFN、lおよびCRISPR/Cas9mなどの部位指向性核酸酵素の使用を含む技術のばらばらなファミリーであるこれらのうち、CRISPR/Cas9は最も広く用いられている(Fichtner他、2014;Lusser & Davies. 2013;Sauer他、2016)。遺伝子編集の主張する利点は、ゲノム修飾を特定のゲノム位置に正確に狙いを定めることができることである。
非常に多様なゲノム改変を遺伝子編集によって意図的に導入することができる(Ahmad, Rahman, Mukhtar, Zafar, & Zhang, 2019; Eckerstorfer et al, 2019; Fichtner et al, 2014; Lusser & Davies, 2013; Puchta, 2017)。これらは、1塩基対の変化から、DNAの大規模な挿入または欠失に及ぶ。挿入には、トランスジェニック、シスジェニック、RNAiベースの形質、制御配列、または複数のトランスジェニックが含まれる可能性がある。CRISPR/Cas9はさらに、単一の遺伝子またはゲノムの複数の部位に、同時に複数の変化を作り出すために使用することができる。マルチプレックスと呼ばれるこの技術は、例えば、遺伝子ファミリーの複数の異なるメンバーを変異させたり、ノックアウトしたりするために使用できる(Fichtner et al., 2014)。
持続可能性にとって重要な問題は、標準的な遺伝子組み換えよりもはるかに精密であると主張される遺伝子編集が、UTの導入なしに有益な形質を導入できるかということである。編集された作物植物について公開されているデータはまだ極めて限られているが、植物の遺伝子編集法もUTsを導入しやすいと予想される理由が存在する。
まず、事実上すべての遺伝子編集プロトコルは、標準的なGM技術、すなわち組織培養とアグロバクテリウム感染または遺伝子銃のいずれかを利用している(Ahmad et al., 2019; Ding, Li, Chen, & Xie. 2016;Eckerstorfer et al., 2019)。これらの技術は、DNA、RNA、タンパク質、またはオリゴヌクレオチドを含むことができる遺伝子編集試薬を、植物細胞に導入する役割を果たす。例えば、アグロバクテリウムの感染は、CRISPR RNAガイド配列およびCas9ヌクレアーゼを規定するDNAを、一過性またはDNA統合を介して導入するために使用され得る。しかし、先に述べたように、組織培養、アグロバクテリウム感染、および遺伝子銃は非常に変異原性が高く、ゲノム全体に何千もの変異を導入することができる(Wilson et al., 2006)。DNAを含まない試薬をプロトプラストに直接取り込むような代替法は、変異原性が低く、したがってUTを導入する可能性が低いかもしれない。しかし、この仮定はまだ実験的に検証されていない。さらに、このような方法は、現在、ほとんどの作物種で利用できない(Ding et al., 2016)。
第二に、動物と植物の両方から得られた新たな証拠は、遺伝子編集自体が、標的部位またはその近傍で意図しない変異を引き起こす可能性があることを示している。これらには、ベクター、細菌、およびその他の余分なDNAの挿入、および大きなDNA欠失および再配列の意図しない導入が含まれる(Biswas et al., 2020;Kosicki, Tomberg, & Bradley, 2018;Li et al., 2015;Norris et al., 2019;Ono et al., 2015)。
第三に、動物からの新しい研究は、正確で意図された編集であっても、オンターゲットのmRNAの誤制御を頻繁に引き起こす可能性があることを示唆している(Sharpe & Cooper, 2017; Tuladhar et al., 2019)。これには、「エクソンスキッピング」と意図せず変更されたRNAスプライシングが含まれる。どちらも、UTをもたらす可能性のある新しいタンパク質コード配列を生成する可能性がある。
第四に、遺伝子編集試薬がゲノムの意図しない部位で切断する可能性があることが、植物と動物の両方で示されている。これらの切断は、オフターゲット編集をもたらし、潜在的にUTをもたらす可能性がある(Ahmad et al., 2019;Biswas et al., 2020;Fichtner et al., 2014;Jin et al., 2019)。
第5に、遺伝子編集は、研究者が指針となる先行研究をほとんど持っていない状況に適用されている。したがって、一部の研究者は、遺伝子編集が「利用されていない種、あるいはおそらく野生種を広く適応した選択肢に発展させ、世界の食料安全保障の向上に役立てる」ために使用できると示唆している(Van Eck, 2018)。さらに、従来の育種では、通常ゲノムの変化から保護されているゲノムの領域をターゲットにするために適応されつつある(Kawall, 2019)。また、多重化の結果は、従来の植物育種を通じて導入することは不可能ではないにしても、困難であろう(Kawall, 2019)。このような遺伝子編集の新規利用は、すでに高い確率でUTを導入する可能性を高めると思われる。
これらの観察結果は、植物の遺伝子編集の結果は不正確で予測不可能であり、使用する技術の組み合わせによっては、遺伝子編集が高い変異原性を持つ可能性があるという結論を支持している。しかし、遺伝子編集は新しい研究分野であり、特に植物については、まだ多くの知識ギャップがある(Ahmad et al., 2019; Schindele, Wolter, & Puchta, 2018)。欠けているのは、遺伝子編集された作物植物と非編集の比較対象の全ゲノム配列である。また、遺伝子編集作物におけるUTの系統的な解析も欠けている。特に商業利用を目的に開発されている作物における、異なるタイプの遺伝子編集技術や異なるタイプの編集の意図しない影響に関して、知識のギャップは大きい。しかし、遺伝子編集された生物におけるUTが頻繁に起こるであろうことは、最近のニュース報道が示唆している。これは、斑点のあるフリースから大きな舌、余分な椎骨、病気、死に至るまで、様々なUTを持つ遺伝子編集動物について述べたものである(Rana & Craymer, 2018)。とはいえ、ほとんどの植物遺伝子編集の論文では、UTの有無を系統的に検証していない。しかし、あるグループは、乾燥ストレス下でのトウモロコシの穀物収量を改善することを目的とした、CRISPR-Cas9が生成したトウモロコシARGOS8遺伝子のプロモーター変異株が、草丈、穂丈、穀物水分において統計的に有意な差を含むUTを示すことを発見した(Shi et al., 2017)。
したがって、バイオテクノロジストたちは、「農業におけるCRISPRは、従来の方法と同一の結果を、より予測可能で、より速く、さらには安価に生み出すことができる単なる『新しい育種法』と考えた方がよい」(Gao, 2018)と頻繁に主張するが、DNAやその他の試薬を導入するための植物形質転換技術の使用、組織培養の使用、外因性DNA挿入の可能性などから、CRISPRやその他の遺伝子編集技術は従来の植物育成よりもGMと近いことは明らかだ(Eckerstorfer et al, 2019; Kawall, 2019).標準的なGMと同様に、遺伝子編集を用いて導入されるUTの数と種類は、一部は導入される新しい形質によって、一部は技術そのものによってゲノムにもたらされる意図しない効果に依存する。したがって、すべての利益、危険性、およびリスクは、独立して派生したnGM形質および作物ごとにケースバイケースで実験的に評価されなければならない(Biswas et al., 2020;Eckerstorfer et al., 2019;Gelinsky & Hilbeck. 2018;Hilbeck et al., 2015)。これには、同系統の系統との全ゲノム配列の比較やゲノム解析が含まれるはずだ。
13.5 持続可能な農業と植物育種
議論したように、GM作物の導入が従来の農業をより持続可能なものにしたわけではない。しかし、遺伝子組換え植物育種や、より広範な遺伝子組換え農業の失敗は、持続可能なシステムへ移行するために必要な変化についての洞察を与えてくれる。
重要な洞察は、再生農業に転換した農民の経験から得られる。これらの農家は、遺伝子組み換え作物とその高投入管理システムを、生物多様性を高め、表土の損失を減らし、自然の土壌肥沃度を高める持続可能な手法(例えば、複合被覆作物、間作、複数年の多作物輪作、遺伝子組み換え作物を使わない不耕起農業、家畜の再導入)と組み合わせている。研究によると、再生農業は財政的および環境的な持続可能性の向上につながることが示唆されている(LaCanne & Lundgren, 2018)。財政的な利益は、主に従来の種子のコストが下がることで発生し、土壌の改善や害虫の圧力の低下により、合成農薬や肥料などの高価で汚染性の高い投入物の必要性が減るからだ。つまり、新しい形質や品種の導入は、持続可能性を向上させるために必要な多くの構成要素の一つに過ぎない。逆に言えば、持続可能な農業を支えるためには、植物育種家が低投入の持続可能なシステムに特化した形質を開発し、その中で選択する必要がある(Murphy, Campbell, Lyon, & Jones, 2007; van Bueren et al, 2011)。
しかし、これまでのところ、有機栽培をはじめとする持続可能なシステムで使用される在来作物品種の大半は、在来システム用に育成され、在来システム内で選抜されている(Murphy et al., 2007; van Bueren et al., 2011)。しかし、慣行システムで高い収量や性能を発揮する品種が、必ずしも有機システムで最適とは限らない(Murphy et al., 2007; van Bueren et al., 2011)。これは、抵抗性育種の主な対象である害虫や病原菌が、2つのシステムで異なるためだ。また、有機栽培では除草剤を使用しないため、雑草の問題も異なる(van Bueren et al., 2011)。さらに、従来のシステム用に開発された非遺伝子組み換え品種は、持続可能性に対して極めてネガティブなトレードオフをもたらすことがある。例えば、宿根を防ぐために導入された半矮性穀物品種は、ミネラル栄養とタンパク質含有量の減少、根の大きさと深さの減少、耐病性と栄養利用効率の低下、雑草抑制力の低下など、さまざまなUTを持つ(Marles, 2017; van Bueren et al., 2011)。これらの欠陥は、高投入システムにおいてマスクされる可能性がある。したがって、持続可能なシステムのために特別に育種された作物や形質は、持続可能なシステムの収量と性能の両方に大きな利益をもたらす可能性がある。
13.5.1 持続可能なシステムのための形質
研究者は、有機農業やその他の持続可能なシステムのための品種を育種する際に、一般的に重要と思われる特性や形質を多数特定している(van Bueren et al., 2011)。これには、養分利用効率の向上(土壌微生物相との有益な共生を促進する活力ある根系または根滲出液)または取り込み(細根の増加)、真菌および細菌病に対する耐性、昆虫耐性(生活史、総形態、物理特性または代謝を変化させて抵抗を促進するなど;Carmona、Lajeunesse、& Johnson. 2011)、雑草に対する競争能力の向上、生物学的ストレス要因に対する耐性の向上および栄養価向上など品質の向上がある。現代の従来型作物品種、特にランドレースや農民品種、その野生親族は、持続可能なシステムのための育種を行う際に、いずれも貴重な変異源となる(Deb, 2014; Dwivedi et al, 2016; van Bueren et al, 2011)。
特定の形質の有用性と影響は、特定の作付けシステムと作物種に依存する(van Bueren et al., 2011)。例えば、不耕起栽培システムでは、雑草が潜在的な問題となり、植え付け深さや土壌水分が問題となる(Joshi, Chand, Arun, Singh, & Ortiz, 2007)。したがって、除草剤を使用しない不耕起栽培システムに有用な形質には、種子の出芽を早める(または雑草に対する競争力を高めるその他の形質)、残渣の分解を早める(または場合によっては遅らせる)、深く播種しても発芽する能力、機械除草に対する耐性がある(Joshi et al., 2007; van Bueren et al., 2011)。その他、作物残渣に生存する害虫や病原菌に対する耐性や、一部の残渣から放出される植物毒性有機酸に対する耐性など、有益と思われる形質もある(Joshi et al.)
ポリカルチャーシステム(例えば、アメリカ大陸の古代「三姉妹」トウモロコシ・豆・カボチャシステム、アフリカやアジアの穀物・マメ科植物システム、またはカバークロップポリカルチャー)での使用を目的とした品種の優先順位は、不耕起栽培の場合と異なっている。有用な形質は、作物間の競争ではなく、補完性を促進する傾向がある。例えば、ポリカルチャーで栽培されたトウモロコシ、カボチャ、豆類は、ニッチ補完性に依存した収量の優位性があることが研究者によって明らかにされた(モノカルチャーで栽培した場合と比較して)。これは、根の養分探索戦略の違いに起因すると考えられる(Chang et al.) したがって、根やシュートの可塑性を促進する形質をポリカルチャー作物用に探索することができる(Zhang et al., 2014)。一方、雑草抑制を促進するために不耕起栽培システムで示唆されるようなアレロパシー形質(van Bueren et al.
自然の生態系は、さらなる特性を示唆することができる。例えば、大草原は、多年生穀物(小麦など)やヒマワリを育種する努力を促してきた(Piper, 1993)。最近のレビューでは、多年生穀物は、発達した根系を必要とし、必要な投入量が少なく、多機能な持続可能システムをサポートできることが示唆されている。例えば、一年草や多年草との間作、および/または放牧システムや長期的な輪作において、傾斜した土地で土壌を保護し改善することができる(Ryan et al.)
13.5.2 持続可能な育種:
考察と戦略植物育種家がアグロエコロジー農法システムで具体化される種類の食糧および種子主権目標を支援するためには、農家やコミュニティのニーズに対応する必要がある。そのためには、環境、規模、持続可能な実践、市場の違いに柔軟に対応できる育種戦略が必要である。低投入で持続可能なシステムは多種多様であり、地域の食料システムのニーズも大きく異なることから、柔軟性と適応性は不可欠である。これらに加えて、気候変動がもたらす不確実性の増大が加わる。
育種家が農家(場合によっては消費者や従来の農家中心の育種会社など他の人々)と協力する参加型植物育種戦略は、これらの複数の成果を達成する有望な方法である(Ceccarelli & Grando, 2019; Cleveland, Daniela, & Smith, 2000; Mercer et al., 2012; Murphy, Lammer, Lyon, Carter, & Jones, 2005; van Bueren et al., 2011)。育種プログラムの開始時に農家が参加することで、理解が深まり、農家が好む形質を持つ品種がより確実になるはずだ。農家の畑やテストプロットで品種試験を行い、選抜プロセスに農家を参加させることも、参加型育種の構成要素である。参加型育種には、現地に適した品種を提供するだけでなく、他にもさまざまな利点がある。例えば、すべての関係者を教育し、権限を与える一方で、コストを削減することができる。また、新品種の取り込みも促進される(Najeeb et al., 2018)。
参加型植物育種は、適切な品種混合物の最適化と利用を促進することもできる。混合品種は、特に自家受粉作物やクローン作物において、レジリエンスを高める有望な戦略である。一つの圃場に異なる品種を植えることで、変動する生物学的または生物学的ストレス条件下での収量の安定性を向上させたり、変化する条件への継続的な適応を可能にしたりすることができる(Cleveland et al., 2000; Murphy et al., 2005; Phillips & Wolfe, 2005)。例えば、東アフリカの農家は、場所によって異なる病気に耐性のある多くの品種のコモンビーンズを混合して栽培している。アンデスの農家の中には、収穫と植え付けを一括して行うことで、多種多様なジャガイモ品種を維持しているところもある(Cleveland et al., 2000)。進化型植物育種は、持続可能な農業のために遺伝的に多様で回復力のある作物を提供できるもう一つの戦略である(Döring, Knapp, Kovacs, Murphy, & Wolfe, 2011; Murphy et al., 2005; Phillips & Wolfe, 2005; Raggi et al., 2017)。この場合、育種家は、例えば生物学的または生物学的ストレス要因に対して、高い固有の遺伝的多様性を持つ異質な複合交配集団を作り出す(Jackson、Kahler、Webster、& Allard、1978)。これらの個体群は、自然の作物栽培システムにおける低投入条件下で、多くの場合、参加型植物育種プログラムの中で順次選択される。時間をかけて育種家や農民が選抜することで、可変的な低投入作物システムによく適応した異質な集団や純系統を生み出すことができる。
植物育種のすべてに共通する最後の重要な考慮点は、支配力、つまり権力という点である。従来の植物育種や遺伝子組み換え作物では、農家が毎年種子を購入することを奨励または義務づけている。また、遺伝子組み換え作物には、研究、種子の保存、育種に使用するための特許制限がある。また、遺伝子組み換え作物の開発には、専門的な知識、設備、試薬が必要である。これらの要因により、種苗会社や大規模な機関の手に支配されている。食料主権と種子主権を支えるには、参加型手法に加え、持続可能な植物育種家は、農民の種子の保存と適応を容易にする品種を優先しなければならない。これらの品種は特許がなく、他の制限、特に種子の共有、育種、研究に対する制限を受けないものでなければならない。
13.6 結論 障害と機会
理論的には、持続可能な農業の広範な要件を満たす遺伝子組み換え作物を作ることがいつか可能になるかもしれないが、実際には、これが実現する可能性は極めて低いと思われる(Kesavan & Swaminathan, 2018; Wickson et al, 2016)。とはいえ、技術的、生態学的、社会的に数々の失敗があるにもかかわらず(例えば、Benbrook, 2018; Fischer, 2016; Wickson et al., 2016)、GM作物は商業的に成功しており、多くの国で特定の商品作物の市場を支配している。この成功は、様々な要因によるものとされている。農家調査によると、HT作物は時間を節約でき、散布の自由度が高く、Bt作物はリスクを軽減するための「保険」と考えられている。また、農家は種子の選択肢が限られていることが多く、代替品に関する知識の喪失や、ある技術が必然であるという思い込みもあって、技術的な踏み絵に閉じ込められてしまうことが多いという調査結果もある(Pechlaner, 2010; Stone & Flachs, 2018)。
しかし最終的には、GM作物の商業的成功は、技術的要因よりもむしろ政治によるものである。科学と技術は中立ではない(O’Brien, 1993)。GM作物は従来の農業を支え、その農業は広大な企業の農産業複合体を支えている(Lima, 2015)。経済的、イデオロギー的な理由から、米国政府、非政府組織、大学、学者はアグリビジネスと協力してGM作物や技術の取り込みを促進し、公的研究、政府の規制、補助金を調整して、その急速な受け入れと拡大を促進し、歓迎できない発見を抑制して代替案を締め出している(Binimelis et al. 2009; Cáceres, 2015; Capellesso et al., 2016; Foscolo & Zimmerman, 2013; Harsh, 2014; Peekhaus, 2010; Pelletier, 2005, 2006; Robinson, Holland, Leloup, & Muilerman, 2013; Schnurr, 2013; Schnurr & Gore, 2015; Schreiner, 2009; Vanloqueren & Baret, 2009; Waltz, 2009). 主流メディアは、GM作物や技術を有望で技術的に成功したものとして一貫して描写することで、こうした努力をさらに後押ししている(Barker, 2014; Stone, 2017)。
こうしたシステム的な偏見にもかかわらず、変化の兆しはある。米国の一部の商品作物農家は、GM作物を放棄し、より持続可能な再生手法に代えた(LaCanne & Lundgren, 2018)。一部のGM植物育種家は、GMから従来の参加型植物育種に切り替えている(Ceccarelli & Grando, 2019; Gilbert, 2016)。一方、有機生産を行っている土地の面積は増加している(Paull, 2017)。さまざまな分野の数多くの研究者が、農法と植物育種の両方がより社会的・生態的に持続可能になることを求め、GM作物を拒否している(Kesavan & Swaminathan, 2018; Kremen & Miles, 2012; McIntyre et al., 2009; van Bueren, Struik, van Eekeren, & Nuijten, 2018)。これらはすべて、トランスジェニックな踏み絵を終わらせ、持続可能な食料システム、すなわち健康な地球上で健康な人々を支えることができる食料システムを支えるために必要な、アグロエコロジーと再生の実践に移行するための科学と政治の機運が高まりつつあるという希望に満ちた兆候である(Anderson & Rivera Ferre, 2020; Valenzuela, 2016)。
