
Macrophages and the maintenance of homeostasis
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7491045/
2020 Sep 15
概要
マクロファージの分極については、これまでに多くの論文が発表されている。これらの章では一般的に、マクロファージが免疫反応を指揮する役割に焦点を当て、これらの分極反応を形成するT細胞由来のサイトカインを強調している。マクロファージが宿主の防御に重要であることを考えると、このような免疫への偏りは理解できる。しかし、マクロファージはどこにでも存在し、多くの異なる細胞プロセスに関与しており、マクロファージを免疫細胞として記述することは間違いなく単純化しすぎている。また、マクロファージは、発生、組織再構築、創傷治癒、血管新生、代謝などの過程で重要な役割を果たしているが、これらの過程を無視している。
本章では、マクロファージが生体内の変換器として機能していることを提案する。ウィキペディアによると、”変換器とは、エネルギーをある形から別の形に変換する装置である “とある。変換器という言葉は、さまざまなエネルギー形態を解釈できる「センサー」と、電圧や電流を切り替えて環境に影響を与える「アクチュエーター」の両方を表す言葉である。マクロファージは、環境からの無限ともいえる多様な入力を感知し、その入力をさまざまな反応結果に変換することができる。したがって、マクロファージは、免疫細胞として機能するというよりも、むしろ、微小環境の変化を解釈し、重要な組織反応を引き起こす細胞トランスデューサーとして広く考えるべきである。本章では、マクロファージが感知するいくつかの感覚刺激と、これらの刺激に対してマクロファージが行う反応について説明し、ホメオスタシスの維持というマクロファージの主要な指令を達成するための方法を説明する。
キーワード
マクロファージ、分極、サイトカイン、発生、炎症
テーマ用語
白血球生成、サイトカイン
はじめに
マクロファージは、メチニコフ1が適応免疫機構を持たない原始的な動物を研究していた時に発見された。この細胞は、ギリシャ語のphagein(食べる)とcytes(細胞)からphagocytesと呼ばれてた。当初、ファゴサイトーシスのプロセスは、組織の再吸収や栄養の獲得といった恒常的なプロセスに関与していると認識されていた。その後、メチニコフ1は、このプロセスが侵入者から体を守るためにも利用できると推論した。そして、メチニコフ1は、このプロセスが侵入者から体を守るためにも使われていることを発見した。メチニコフ2のファゴサイトーシス現象に関する先見の明は、微生物に対する細胞性免疫に関する今日の理解の基礎となった。このように、マクロファージが宿主の「バランス」を促進する(ホメオスタシス)という当初の概念はほとんど無視され、細胞性免疫反応へのマクロファージの関与が本質的に影を潜めていた。
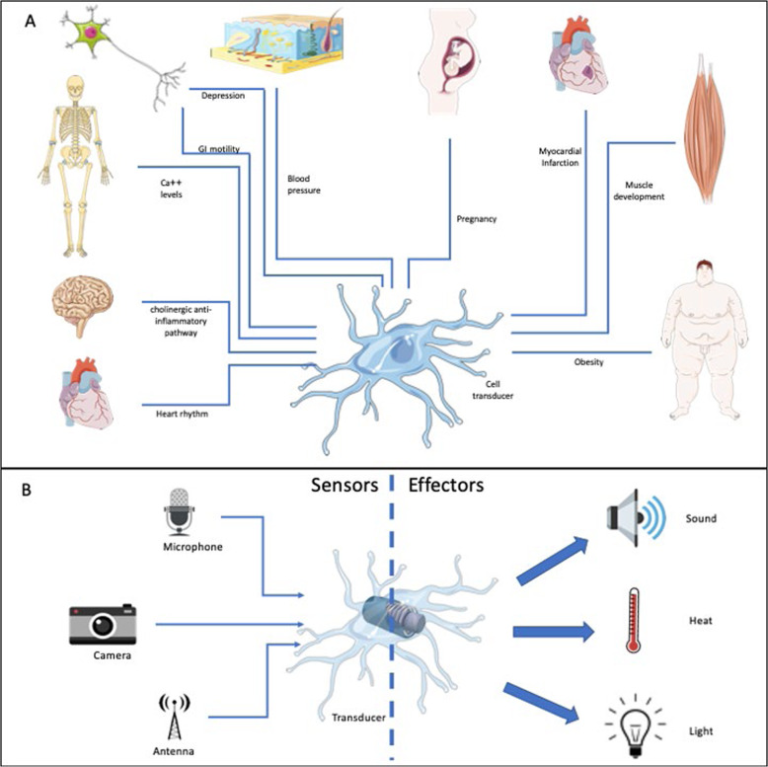
マクロファージは、体内のほぼすべての組織に存在し、臓器の機能を正常に維持している。マクロファージは、鉄、ビリルビン、カルシウム、脂質、アミノ酸などの代謝に関与し、体内のこれらの物質の濃度をほぼ一定に保つことに貢献している3-7。これらの恒常的な機能のほとんどは、100年以上前にヒトデで確認された原始的なプロセスであるエフェロサイトーシスに関連している。マクロファージの貪食作用は、膨大な数の死細胞や組織の残骸を除去し、再利用する。このようなクリアランスは、すべての生物で行われており、適応的な免疫反応がなくても、場合によっては血液がなくても、何の支障もなく進行する。組織のマクロファージが治癒に貢献することは、もともと血液を持たない無脊椎動物のMetchnikoff1によって報告された。しかし、高等脊椎動物の初期胚では、血管が発達する前に、組織マクロファージが治癒や組織再生のプロセスに貢献することができる。マクロファージは定常状態では重要な役割を果たしており、通常、マクロファージは唯一の組織に常駐する「免疫」細胞である。目8,関節9,乳腺10,卵巣11などに存在するマクロファージは、組織からの入力信号を統合し、隣接する間質細胞に指示を伝えることで、組織の健全性を維持している。マクロファージが介在する恒常性は非常に重要であり、マクロファージはヒトの母乳中にも存在し、乳児の消化管バランスの制御に寄与していると考えられる12。また、マクロファージは、新生児の炎症の制御や治癒反応の促進に重要な役割を果たす調節機能を発揮する13。マクロファージは、表皮や角膜、血管が存在しない関節の内側などに存在している。マクロファージは、組織から情報を得て、それを翻訳して反応を引き起こすトランスデューサーとして機能する重要な細胞である。これらの反応は、通常、その器官の日々の活動に不可欠な生理学的機能に関連している(図1a)。
図1 トランスデューサーとしてのマクロファージ
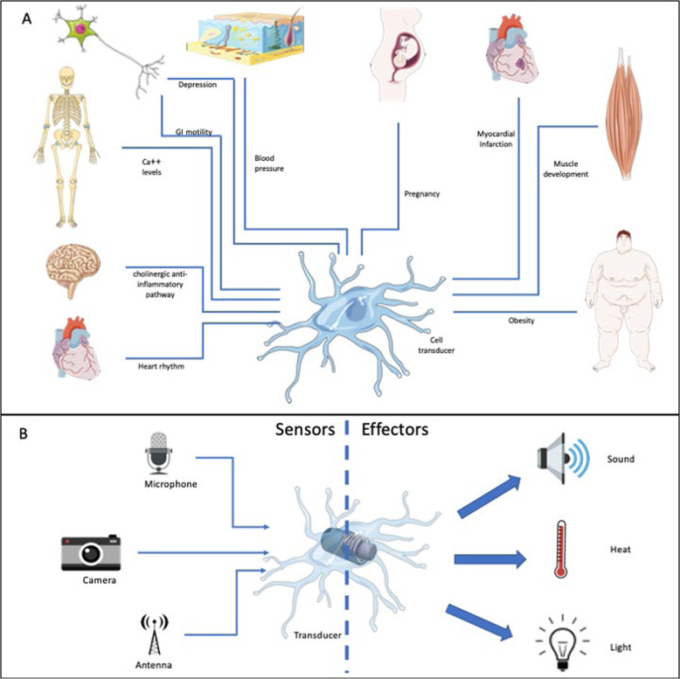
a 異なる組織のマクロファージは、微小環境から情報を収集し、この情報を処理し、個々の器官の特定の機能のために重要な化学反応を生成するために情報を伝達することによって機能する。
b 異なるエネルギー形態として描かれた異なる刺激または刺激の組み合わせが、マクロファージによって受け取られ、異なる出力に変換される。この画像はMedical ART(https://smart.servier.com/)を使用して作成した。
マクロファージは、微小環境を感知し、臓器のニーズに応えることができる細胞として、様々な場面で説明されてきた。メチニコフが述べているように、マクロファージは常にバランスを求めているが、同時に臓器がその固有の機能を発揮するのを助けている。例えば、妊娠中には、ホルモンからのシグナルを感知し、それを伝達して免疫寛容な環境を構築し、本来なら拒絶されるはずの胚の発育を可能にする。また、胎盤の構築にも貢献しており、これがなければ胎児の発育はあり得ない14。心臓では、マクロファージが心筋細胞と相互作用し、心筋細胞を感知してその再分極を促進し、それによって心臓の伝導を維持する。最近の興味深い研究では、マクロファージが組織からの浸透圧シグナルを伝達し、高塩分食を与えられたマウスやラットの血圧を制御できることが明らかになった16。実際、マクロファージはNaCl高張性から生じるシグナルを認識し、塩分濃度の高い方向に移動することができる17。これらは、恒常性の維持というマクロファージの最も重要な機能を示すいくつかの例にすぎない。マクロファージは、トランスデューサーとして機能することで、微小環境を解釈し、バランスを保つために近隣の細胞に指示を与える(図1b)。これらの「出力」は、マクロファージの刺激と関連する炎症性サイトカインよりもはるかに複雑である。細胞外マトリックスを再構築するマトリックスメタロプロテアーゼ、血管新生を誘導する血管内皮成長因子(VEGF)やトロンボスポンジン、創傷治癒を促進する成長因子などが含まれる。このような背景から、マクロファージを単に「免疫」細胞と考えるのは、あまりにも単純化しすぎている。
マクロファージが環境を感知する能力は、単なる理論上の可能性ではなく、がんなどの病気の診断や治療を改善するために利用することができる。がんの治療における大きな課題の1つは、早期の転移を検出することである。Gulatiたち18は最近、アルギナーゼ-1プロモーターの制御下でルシフェラーゼを発現する、人工的なマクロファージレポーターを開発した。したがって、この細胞が腫瘍微小環境に引き寄せられると、腫瘍に関連するプロファイルを持つマクロファージ(腫瘍関連マクロファージ)となり、ルシフェラーゼを発現し始めるので、転移の可能性を容易に検出することができる18。マクロファージのトランスデューサー機能について理解が深まるにつれ、マクロファージの単純なM1/M2分類は現実を反映しておらず、入力刺激を変えるだけで、これらの細胞を予測可能な方法で操作できることがわかっていた。
マクロファージの起源
最近、いくつかのグループがマクロファージの発生に関心を寄せている。19-27年には、卵黄嚢由来の組織に留まっているマクロファージは、造血系の入力とは無関係にほとんど維持されていることが明らかになった。これらの観察結果から、新たに移動してきた単球由来のマクロファージは炎症を起こすものであり、組織内に留まっている細胞は炎症を緩和し、ホメオスタシスを回復する機能を持っているという考えが生まれた28-30。組織に存在する細胞を含むすべてのマクロファージは、パターン認識受容体を発現しており、活性化刺激に反応することができる。従って、卵黄嚢由来の組織常駐マクロファージを含む全てのマクロファージは、この章の後のセクションで説明される様々な刺激入力に反応することができる。さらに、すべてのマクロファージは、微小環境の変化に応じてその表現型を変化させる可塑性を持っている。したがって、すべてのマクロファージは、アデノシン、PGE2,あるいは組織内で産生される他の調節分子に反応して、炎症反応を弱め、組織修復に向けた反応に偏らせる可能性を持っている。組織内に存在するマクロファージに起因するとされる抗炎症反応のいくつかを解釈する別の方法があるかもしれない。我々の考えでは、組織内に存在するマクロファージを免疫寛解の表現型に偏らせる組織由来の2つの主要な影響があると思われる。第一に、定常状態では、組織常駐マクロファージは、正常な組織形成を維持するために細胞成長を促進する組織由来のモジュレーターに長時間さらされている可能性が高い。これらの調節因子には、アデノシン、PGE2,および後述するレゾルビンが含まれる。第二に、マクロファージの成長因子であるマクロファージコロニー刺激因子(M-CFS)は、多くの細胞によって産生され、マクロファージを成長促進と血管新生の表現型に偏らせる(Hamidzadeh er al)。定常状態ではM-CFSが優勢であるが、炎症時には顆粒球M-CFSが一過性に産生され、それに曝されたすべてのマクロファージの炎症反応を促進する。したがって、以下に述べる入力と反応は、マクロファージの起源にかかわらず、すべてのマクロファージに共通するものである。
マクロファージの生理機能を変化させる感覚的入力
病原体や損傷に関連する分子パターン
Metchnikoffは、炎症なくして治療はあり得ないと考えてた35。現在、我々は炎症とそれに関与する分子について多くのことを知っている。また、炎症を制御しないと組織の病理を引き起こす可能性があるため、治療プロセスを開始するだけでなく、炎症をしっかりと制御しなければならないこともわかっている。本章では、炎症の制御に焦点を当てる。しかし、まず始めに、炎症を引き起こす病原体や損傷に関連する分子パターン(PAMPsやDAMPs)について述べておきたい。簡単に言えば、マクロファージは、危険な兆候を示す分子パターンを認識する多様な受容体を備えている。マクロファージは、これらのシグナルが受容体に結合すると、遺伝子発現に大きな変化が生じる。例えば、ナノモル濃度の細菌性リポ多糖(LPS)にさらされたヒトのマクロファージは、安静時のマクロファージと比較して、約4500の遺伝子を異なる形で発現している36。このような遺伝子発現の変化により、M1マクロファージと呼ばれる炎症性のマクロファージが形成される(注:「古典的に活性化されたマクロファージ」という用語は、通常、トール様受容体(TLR)リガンドやサイトカインであるインターフェロン(IFN-γ)にさらされた抗菌性のマクロファージに対して用いられる)。定常状態にごくわずかな乱れが生じただけでも、刺激された炎症性(M1)マクロファージの集団が形成される。これらの細胞は、様々な炎症性サイトカインやケモカインを発現する。これらの炎症反応の誘導は、TLRシグナルの様々な負の制御因子によって慎重にバランスがとられている。これらの制御因子は、転写因子、アダプター複合体、シグナル伝達経路、および受容体リガンドの結合自体を阻害する働きをする。これらの制御因子については、文献37を参照のこと。37.
サイトカイン
マクロファージが局所的な微小環境で産生されるサイトカインに非常に敏感であるかどうかについては議論の余地はない。マクロファージの生理機能に影響を与えるサイトカインとして最も広く研究されているのは、IFN-γとインターロイキン4(IL-4)である。実際、この2つのサイトカインは、マクロファージの生理機能に劇的に異なる影響を与えるため、マクロファージはこれらのサイトカインのどちらかに曝されるかによって、M1とM2に分類される。マクロファージは、IFN-γ38の高親和性受容体を発現し、IFN-γに応答して、遺伝子発現に無数の変化を与え、Mackaness39,Cohnら40,Nathanら41が最初に述べた強力な抗菌細胞になる。簡単に言えば、IFN-γを産生してマクロファージを活性化し、細胞内の生物を殺すために、細胞媒介性の免疫反応が行われる。
マクロファージの生理機能におけるIL-4の役割についても広く研究されている42,43が、なぜかまだよくわかっていない。TH2サイトカインであるIL-4は、主に蠕虫感染やアレルギー反応に反応して産生されるが43,マクロファージに大きな影響を与え、根本的に異なる活性化状態(当初は「交代活性化」状態と呼ばれていた)を引き起こす44。一つ目は、オルターナティブ活性化マクロファージ(AA-Mφ)を同定するバイオマーカーに関するものである。マウスのシステムでは、外因性IL-4に対するマクロファージの反応は非常に劇的である。我々の研究では、IL-4投与後4時間以内に、活性化されたネズミのマクロファージは、休息中のマクロファージと比較して、23の転写産物を25倍以上に増加させた(表1)。これらのアップレギュレートされた遺伝子には、Ym1 (Chitinase-like 3), Retnla (Fizz1, RELMα), Mrc1 (元々AA-Mφの同定に使用されたマンノース受容体)が含まれてた44。ヒトのIL-4処理マクロファージの同定はそれほど容易ではない。マウスのIL-4処理マクロファージによって発現される転写産物のいくつかは、マウスのAA-Mφに関連するキチナーゼのものを含めて、マウスのマクロファージに特有のものである。IL-4がヒトのマクロファージの遺伝子発現に影響を与える程度もより緩やかなようで、バイオマーカーの同定をより困難にしている。我々の手では、マウスのIL-4誘導遺伝子トップ23のうち、ヒトのマクロファージで広範囲に発現が増加したのは1つだけであった。このように、組織中のヒトAA-Mφの同定は些細なことではなく、多くのグループが単にヒトAA-Mφに関係のないネズミのマーカーを使用してこれらの細胞を誤って同定した。いわゆるM2マクロファージに関する混乱の第二のポイントは、創傷治癒におけるIL-4の役割である。Wynnら45-48は一連の優れた論文の中で、マウスシステムを用いて、IL-4処理マクロファージにおけるアルギナーゼ発現の誘導について述べている。アルギナーゼはアルギニンをポリアミン生合成の前駆体であるオルニチンに変換するため、IL-4で処理したマクロファージは創傷治癒反応を示した。これらの研究に続いて、多くのグループがM2マクロファージを創傷治癒を促進する細胞として暫定的に同定した。IL-4が創傷治癒にどの程度寄与しているのか、また創傷治癒マクロファージの産生にIL-4が必要なのかは、マクロファージの生物学上、混乱を伴う問題であると思われる。一見すると、基本的な治癒反応が、IL-4のように主に適応免疫細胞によって産生される単一のサイトカインに大きく依存していることは意味をなさないかもしれない。実際、適応免疫反応とは無関係に創傷治癒が起こる実験例は数多くある。例えば、成熟したT細胞を持たないSCIDマウスでは、創傷治癒が正常に行われる。49 IL-4R-ノックアウトマウスでは創傷治癒マクロファージが観察され、創傷治癒マクロファージの表現型はIL-4やIL-13を必要としなかった50。さらに最近の研究では、M1マクロファージからM2マクロファージへの移行には、IL-4やIL-13とは無関係に他の経路が関与していることが示されている51。以下では、免疫サイトカインであるIL-4の明らかな寄与がないにもかかわらず、マクロファージが組織再生を促進する例をいくつか紹介する。
表1 マウスマクロファージのIL-4処理により誘導される遺伝子発現
| シンボル | 名前 | ログFC 2 | 調整 P値 | 人間(FC 2)* |
|---|---|---|---|---|
| Chil3 ** | Chitinase-like 3 (Ym1) | 8.7 | 9.1 − 13 | バツ |
| Cd209e | CD209e抗原 | 6.9 | 1.9 −6 | 3.16 |
| Itgb3 | インテグリンベータ3 | 6.8 | 7.8 −9 | – |
| Ear11 | エオシン関連リボヌクレアーゼA | 6.3 | 8.5 −6 | – |
| Flt1 | FMSチロシンキナーゼ | 6.3 | 4.9 −7 | – |
| Serpina3g | セリンペプチダーゼ阻害剤 | 6.21 | 6.2 −7 | – |
| Chi3l4 | キチナーゼ3-like4 | 5.9 | 6.5 −6 | – |
| Pdcd1lg2 | プログラムされた細胞死1リガンド2 | 5.7 | 2.3 −10 | 1.25 |
| Slc7a2 | 溶質キャリアファミリー7 | 5.7 | 3.3 −8 | – |
| Cdh1 | カドヘリン1 | 5.4 | 1.8 −7 | – |
| Ntrk1 | 神経栄養性TKRタイプ1 | 5.4 | 1.9 −4 | – |
| Tmem26 | 膜貫通タンパク質26 | 5.3 | 7.8 −4 | 2.72 |
| Tslp | 胸腺間質リンホポイエチン | 5.3 | 1.0 −4 | – |
| Il4i1 | IL-4誘発性1 | 5.2 | 1.9 −9 | – |
| Il31ra | IL-31受容体A | 5.1 | 1.0 −4 | – |
| En2 | 巻き込まれた2 | 5.0 | 7.8 −6 | – |
| シッシュ | サイトカイン誘発性SH2 | 5.0 | 7.7 −6 | 4.79 |
| Mrc1 ** | マンノース受容体 | 5.0 | 3.6 −7 | 2.37 |
| Apo7c | アポリポタンパク質L7c | 5.0 | 1.8 −6 | バツ |
| Socs1 | サプレッサーサイトカインシグナル伝達 | 5.0 | 7.7 −8 | 3.45 |
| Retnla ** | レジスチン様アルファ(Fizz1) | 4.8 | 1.6 −9 | バツ |
| Ccl7 | ケモカイン(CC)リガンド7 | 4.6 | 4.3 −3 | – |
| Ccl12 | ケモカイン(CC)リガンド12 | 4.6 | 4.6 −4 | – |
*IL-4で処理したヒトのマクロファージにおける倍数変化。「x」はマウスに特異的でヒトには見られない遺伝子を示す。
**太字の記号は、マウスM2aマクロファージで最も広く使われている3つのバイオマーカーを示す。
我々の意図は、マクロファージの生物学に対するサイトカインの貢献度を最小限に抑えることではない。実際、マクロファージは多くの異なるサイトカインの受容体を持っている。組織内のマクロファージは複数のサイトカインに遭遇し、その反応は当初のM1/M2という呼称からは想像できないほど複雑なものになっていると思われる。例えば、IL-4は単独ではマクロファージによる腫瘍壊死因子(TNF)の分泌を促進しない。52 IL-6はマウスのAA-Mφのマーカーの発現を増強するが、IFNγとの併用では炎症性サイトカインであるIL-1βとTNFの産生を増加させる 53 サイトカインは多くのマクロファージの機能を調節し、マクロファージ自身がサイトカインの供給源となりうることを理解すべきである。IL-33のオートクラインシグナルは、ラット肺胞マクロファージにおいてMMP2やMMP9などのマトリックスメタロプロテアーゼのアップレギュレーションを引き起こす54。LPSで刺激されたマクロファージの腫瘍活性の多くは、オートクラインIFNαおよびIFNβシグナルに依存している55。腫瘍関連マクロファージによる自律的なIL-10の産生は、これらのマクロファージが炎症性のIL-12を産生する能力を低下させる56。IL-10はマクロファージの抗炎症プログラムという点でよく知られているが、唯一の制御サイトカインではない57,58。IL-21は、マウス腹膜マクロファージにおけるLPS誘導性の炎症性サイトカインであるIL-1β、TNF、IL-6の発現を抑制する60。IL-35は、マクロファージのトランスフォーミング成長因子(TGF)βを活性化し、細胞外マトリックスの沈着を誘導することで、マクロファージの創傷治癒機能を促進する61。したがって、サイトカインは、マクロファージの炎症反応を刺激することも、炎症を鎮めて組織修復機能を促進することもできる。
マクロファージの重要な機能の多くは、適応免疫反応に関連するサイトカインが存在しなくても正常に進行する。マクロファージは、筋肉の発達において、よく知られた役割を果たしている62。ウエイトトレーニング中、同心円状の収縮により筋繊維が損傷する。マクロファージは、組織の残骸を除去するために筋内マクロファージは、死んだ細胞や死にかけている細胞から放出されるDAMPsに反応して、炎症の表現型をとる。ウェイトトレーニング後に生じる筋肉痛は、このプロセスの結果である。筋マクロファージはその後、生理的変化を起こし、筋に存在する幹細胞であるサテライト細胞にシグナルを送り、新しい筋線維に分裂・分化させる。この新しい筋繊維は、回復後の筋肉に強度を与える。発育中の筋肉にリンパ球が移動したという報告は知らないし、我々の知る限り、T細胞とB細胞を遺伝子的に欠失させた実験動物は正常に筋肉を発達させている63,64。したがって、運動中に損傷した筋肉からIL-4が放出される未知の生得的な源がない限り、マクロファージによる筋肉の発達促進におけるIL-4の決定的な役割を決定することはできない。心血管疾患からの回復期にも同様の修復プロセスが起こる可能性がある。マクロファージは心筋梗塞に反応して心臓内に移動し、壊死組織の除去を促進するようになる。その後、マクロファージはその表現型を変え、幹細胞の分化と治癒を促進する。65 自己反応性T細胞は発生過程でクローン的に削除されているため、抗原特異的リンパ球の寄与は最小限であると考えられている。このように、心臓の創傷治癒過程は、IL-4が存在せず、適応免疫の寄与もない状態で行われる。
免疫複合体
免疫複合体(IC)は、マクロファージの生理機能に大きな影響を与える感覚刺激物質である。マクロファージが高密度のICの存在下で刺激されると、マクロファージは複数の炎症性メディエーターの産生を抑制し、抗炎症性サイトカインや成長促進因子、血管新生因子の産生を増加させる。このように、IC存在下での刺激は、IC非存在下での刺激と比較して、マクロファージの生理機能に劇的な変化をもたらす。当初、このような遺伝子発現の変化が観察されたとき、66-69は、抗原特異的な免疫反応を促進する上でのIgGの重要性が十分に認識されていることを背景に、その解釈がやや難しかった。現在では、マクロファージはIC存在下での刺激に応じて生理機能を変化させ、体液性免疫反応を停止させ、必要な組織修復プロセスを開始させると考えられている。すべての免疫反応は組織に損傷を与える可能性があるため、治癒反応を開始するためには、すべての免疫反応に恒常性の調節が必要である。そのため、外来抗原に反応してIgG抗体反応が産生される。ICはFcγ受容体を架橋して貪食を開始するが、マクロファージはICを免疫反応を終了して組織修復を開始するシグナルとして解釈する。
このようなICに対する反応は、過剰な体液性免疫反応による組織の損傷を防ぐことができるが、宿主にとっては有害な場合もある。リーシュマニア症では、大量のICがマクロファージによる細胞内寄生虫の殺傷能力の低下と関連しているとされる。びまん性皮膚リーシュマニア症と呼ばれる最も重篤な皮膚リーシュマニア症では、IgG ICの海の中を文字通り泳いでいる真皮マクロファージの中で、寄生虫が制御不能な状態で増殖している70。呼吸器系ウイルスも、このマクロファージの生理機能の変化を利用している可能性がある。71 呼吸器合胞体ウイルスや一部のネココロナウイルスは、体液性免疫反応の後に発症する「ワクチン関連呼吸器疾患強化」と呼ばれる現象と関連している72。
内因性調節因子
マクロファージ自身は、恒常性の維持に機能する多くの分子を産生している(図2)。一般的に、これらの分子は、TLR刺激などの活性化シグナルに反応して産生される。TLR刺激は、炎症反応を促進するだけでなく、炎症を抑制し、恒常性を回復するための調節機構を活性化する。炎症が長引くと組織の破壊につながるため、これは重要なプロセスである。ここでは、アデノシン、プロスタグランジンE2(PGE2)レゾルビン、リポキシンなど、いくつかの内因性調節因子が、炎症の積極的な解決に寄与していることを説明する。
図2 内因性および外因性の調節因子に対するマクロファージの反応
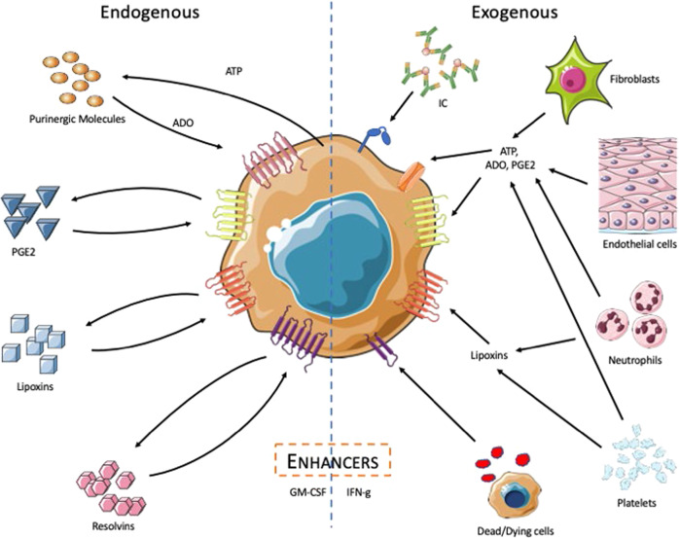
左:刺激を受けたマクロファージはATPを産生・放出する。マクロファージの外部酵素であるCD39とCD73は、ATPを速やかにアデノシンに変換し47,アデノシンの高親和性受容体を介して、炎症性メディエーターの産生を停止させ、成長関連因子や血管新生因子の産生を誘導する。刺激されたヒトのマクロファージは、PGE2の合成とPGE2の受容体をアップレギュレートし、PGE2の調節作用に非常に敏感になっている。マクロファージはまた、脂質をリポキシンやレゾルビンに異化し、炎症反応を弱める。右:繊維芽細胞、内皮細胞、好中球、血小板などの近隣の細胞も、これらの調節分子の供給源となりうる。これらの近隣細胞は、初期の活性化反応を増幅させる「エンハンサー」分子を産生するように刺激することもできる。この画像はMedical ART(https://smart.servier.com/)を使用して作成した。
アデノシンはA1r、A2Ar、A2Br、A3という4つのGタンパク質共役型受容体を介して感知され、そのうちの2つ、A2ArとA2BrはGαsシグナルを介して細胞内cAMPレベルを上昇させる75。A2ArとA2Brのシグナルは、炎症性サイトカインの産生を抑制し、マクロファージによるIL-10の産生を増加させることで、アデノシンの調節作用を増強する76-78。アデノシンの感知はまた、VEGFの産生を引き起こす79-81。VEGFは血管新生と創傷治癒に重要な成長因子である82。
PGE2は、細胞膜のアラキドン酸に由来する内因性脂質である。PGE2は、最も広く作用し、最も研究されているプロスタグランジンである。PGE2は、マクロファージがPAMPsに応答して、また、アポトーシス細胞を取り込んだ際に産生される。プロスタグランジンやロイコトリエンの合成からレゾルビンやリポキシンの産生へと切り替わることが、炎症の解決促進段階であると広く考えられている86。しかし、マクロファージの活性化に伴い、PGE2は抗炎症反応を誘導し、恒常性の回復に積極的な役割を果たしていることを理解すべきである87。PGE2が感知されると、TNF、IL-1β、IFNβなどのマクロファージによる炎症性サイトカインの放出が抑制される88-90。PGE2の抗炎症作用のほとんどは、EP2およびEP4受容体を介して産生され、細胞内cAMPの放出につながる91。 -また、マクロファージ由来のPGE2は、好中球の動員に影響を与えることで、組織損傷部位における炎症の解決に重要な役割を果たしていることが示されている95。
レゾルビンは、マクロファージのホメオスタシスに貢献するもう一つの脂質メディエーターファミリーである。レゾルビンは、オメガ3系多価不飽和脂肪酸であるエイコサペンタエン酸とドコサヘキサエン酸に由来しており、それぞれEシリーズ、Dシリーズのレゾルビンと呼ばれている96。現在までに、レゾルビンの受容体として知られているのは、Gタンパク質共役型受容体GPR32,GPR18,ChemR23である97。ヒトのマクロファージは、大腸菌や黄色ブドウ球菌の刺激に反応して、レゾルビンD2(RvD2)とD5(RvD5)を産生することが示されている98。100 RvD1およびRvD2は、ヒト肺胞マクロファージが産生するIL-6およびTNFのレベルを低下させ、ヒト単球由来マクロファージが産生するTGFβのレベルを上昇させることも示された。
リポキシンは、マクロファージの活性化を抑制するアラキドン酸由来の脂質メディエーターの一種である。LXA4とLXB4は、リポキシゲナーゼ(LO)酵素である15-LO、5-LO、12-LOによって産生されるリポキシンである102。これらの安定したアスピリントリガー物質であるAT-LXA4とAT-LXB4は、アスピリンの存在下でCOX-2によって産生される103。ALX/FPR2は、LXA4およびAT-LXA4のGタンパク質共役型受容体である104 LXA4は、ヒト肺胞マクロファージでピコグラムレベルで産生され、LPS刺激に反応して増加することが示されている105,106。LXA4は、IκBおよび核内因子-κBのリン酸化を抑制することにより、マクロファージのLPSに対するTNFレベルを低下させる107。また、LXA4は、RAW264.7マクロファージにおける活性酸素種の産生および顆粒球CFS(G-CFS)の分泌を抑制することが示されている108。
上述の内因性分子は、炎症性サイトカインの産生を抑制することで、マクロファージの活性化を一過性のものにしている。その一方で、マクロファージを組織修復プロセスの調整役に再プログラムする。例えば、レゾルビン、特にRvD2の機能の一つは、感染部位への多形核(PMN)細胞の浸潤を停止させ、貪食を増加させることで、感染部位から破片やアポトーシス細胞を除去することであると推測される109,110。111 外因性のアデノシン、PGE2,レゾルビン、リポキシンは、マクロファージに感知され、その表現型を変化させる。114-118 血小板と好中球はリポキシンの主な供給源である。119,120 したがって、マクロファージは組織環境中の異なる供給源から受け取ったシグナルを伝達し、恒常性を促進することができる。
マクロファージと発生
卵黄嚢27の前駆細胞に由来するマクロファージは、発生の初期に胚に現れる。彼らは、胚発生において重要な役割を果たしている。発生過程では、胎児が完全に形成された後に、最終的な形になるように組織を再構築する必要がある。この再構築には、細胞を除去して新しい空間を開いたり、器官の大きさや形を変更したりすることが含まれる。この再構築は、主にプログラムされた細胞死とマクロファージによるアポトーシス細胞の除去によって達成される121。その最も良い例の一つが、発生過程における指の個別化である。マクロファージは、この過程で細胞の破片を再吸収し、組織の再編成を促進するのに欠かせない細胞である122。胚発生期にマクロファージの機能が変化すると、組織や臓器の発生に不具合が生じることがあるが123,B細胞とT細胞を欠損したマウスでは、明らかな発生の不具合は見られない。124 マクロファージによるアポトーシス細胞の除去は、炎症を抑制するだけでなく、自己抗原に対する寛容性の発達を促進する表現型と関連している。
神経系への入力
マクロファージは、中枢神経系(中枢神経系)の発達と維持に重要な役割を果たしている126,127。これらの細胞には、ミクログリアだけでなく、いわゆる中枢神経系関連マクロファージも含まれる。マクロファージは、生命維持の基本となる栄養因子を分泌することで、神経細胞の分化・生存を促進し、神経細胞の機能を維持している。中枢神経系におけるマクロファージ制御の欠如は、うつ病や炎症性腸疾患などの心身症と関連している。128 「うつ病のマクロファージ理論」と呼ばれる画期的な論文129が、最近のレビューで再評価されている。130 2000,Tracey et al 131は、迷走神経刺激が炎症性サイトカインの産生を抑制することを示し、「コリン作動性抗炎症経路」を特定した。その後まもなく、Tracey132は、神経系がどのように炎症反応を調節して、反射的に炎症をコントロールしているかについて、重要な総説を書いた。この研究は、神経系との相互作用のトランスデューサーとしてマクロファージが参加していることを補強している。さらに最近では、Chiu et al 133が、バクテリアが侵害受容器の感覚ニューロンを直接刺激できること、そして侵害受容器ニューロンと免疫細胞の間には広範なクロストークが存在することを明らかにした。CGRP、ガラニン、ソマトスタチンなどの神経ペプチドの放出は、TNFの転写を抑制することができる133。マクロファージと神経系の間には、消化管運動を制御する上で重要な関係が存在する134。マクロファージは、神経線維に沿って密接に配置され、消化管運動を制御するために神経細胞に継続的なシグナルを提供することが示された。さらに、神経細胞はマクロファージのホメオスタシスの維持に重要な役割を果たしている134。De Schepper et al 135は、血管系、腸管神経細胞、腸管運動に関連するいくつかの重要な機能を制御しているため、ホメオスタシスの維持に重要な腸管マクロファージの集団について述べている。最近の研究では、腸内感染により神経細胞が急速に失われること、マクロファージがβ2アドレナリン受容体シグナルを介して神経細胞死を防ぐ神経保護的な役割を果たすことが明らかになっている135。
結論
本研究では、マクロファージがトランスデューサーとして機能するという概念を提唱した。マクロファージは、体のほぼすべての器官の発生と機能に不可欠な細胞であり、胎児の発生初期に存在する。マクロファージは、内因性または外因性のさまざまなシグナルを組織内で感知し、それに迅速に反応することができる。したがって、マクロファージをM1またはM2に分類することは適切ではない。これは、マクロファージがサイトカインにしか反応しないことを意味するだけでなく、2つのサイトカインにしか反応しないことを意味するからである。我々は、マクロファージの主要な機能は「バランス」または「ホメオスタシス」を達成することであるというメチニコフの視点を好んでいる。炎症はバランスを大きく崩すものなので、マクロファージの主要な役割の一つは、炎症を抑制し、修復を促進することである。このようなマクロファージの役割は、なぜかあまり評価されていない。マクロファージは本当に特別な細胞だ。マクロファージは、その可塑性により、環境の無数の変化に対応し、特定の瞬間の生理的ニーズを満たすことができる。マクロファージが知覚する感覚入力とその結果としての生理的反応をより深く理解することで、我々はマクロファージを病気の治療のために操作することができるようになる。