
Why is USG rewriting the history of the mRNA vaccines?
https://rwmalonemd.substack.com/p/why-is-usg-rewriting-the-history
ロバート・W・マローン医学博士、MS
著者 ジル・グラスプール・マローン医学博士
Nature Biotechnologyは2022年7月、「The COVID-19 vaccine patent race 」と呼ばれる論文を発表した。この論文の中で、Nature Biotechnologyは、mRNAワクチン接種の最初の実用化が2000年に行われたと主張している。 これは明らかに誤りである。 この科学雑誌の偶発的な見落としとは考えにくい。 このmRNAワクチン接種を実用化した最初の生体内試験(動物)実験は1989年に行われた。最初の実験は、HIVウイルスのgp120タンパク質を産生するマウスのmRNAワクチン接種と、その後のmRNAワクチン接種マウスにおけるgp120抗体の産生であり、2番目の実験は、ヒト幹細胞保有SCIDマウスのHIV nef mRNAワクチン接種とそれに続くHIVチャレンジであった。この2つの実験は米国特許庁に提出され、これらの請求項は9件の発行済み特許として承認されたが、その優先日はいずれも1989年3月21日であった。 つまり、これらの実験は1989-90年に行われたことになる。 これら2つの例と9つの特許は、この記事の末尾に掲載した。 なお、これらの特許には、技術に関する広範な説明も含まれている。
ネイチャー誌が、マローン博士と特許に記載された他の共同発明者の初期の研究を、mRNAワクチンのプラットフォームと技術の実際の発明者として適切に引用し、認めなかったのは、今回が初めてではない。2021年9月、Nature誌は、Eli Dolginが執筆した、最初の原理証明実験がずっと後の2005年に行われたと主張する論文を発表した。 その論文は、Dr. Maloneが著者のEli Dolginに誤りについて通知したにもかかわらず、訂正されることはなかった。ドルジン氏は、特許を読むことさえ拒否し、その論文に記載されている時系列が誤りであるという事実を考慮しようとしなかった。 このため、主要な新聞、雑誌、ジャーナルは、マローン博士は実際の原理証明のmRNAワクチン実験とは何の関係もないと主張する連鎖を引き起こしたのである。
その後、米議会調査局(CRS)は、議会向けに「mRNAテクノロジー」と題する本を出した。A Primer」 2022年5月に出版された。これは、mRNAワクチンの歴史と技術に関する政府の記録である。繰り返すが、発明と年表(上記のNatureの記事から引用したCRS)は間違っているだけでなく、明らかな虚偽である。CRSに問い合わせたところ、彼らはその出版物を訂正することを拒否した。CRSからの返信のスクリーンショットはこちら。

問題は、なぜなのか?何が起こっているのだろうか? 特許記録は、米国特許商標庁が裁定し、発明者資格を付与する公的な政府機関であることから、極めて明確である。 それとも、この誤った時間軸にもっと意味を持たせるべきなのだろうか?mRNAワクチンの真の発明者を議会の記録から削除することを政府が選択したことについて?これは見た目通りなのだろうか?政府は文字通り、ロバート・マローン博士がこの発明で果たした役割を抹消し、彼と彼の業績、そして彼の共同発明者たちを歴史から抹消しようとしているのだろうか?
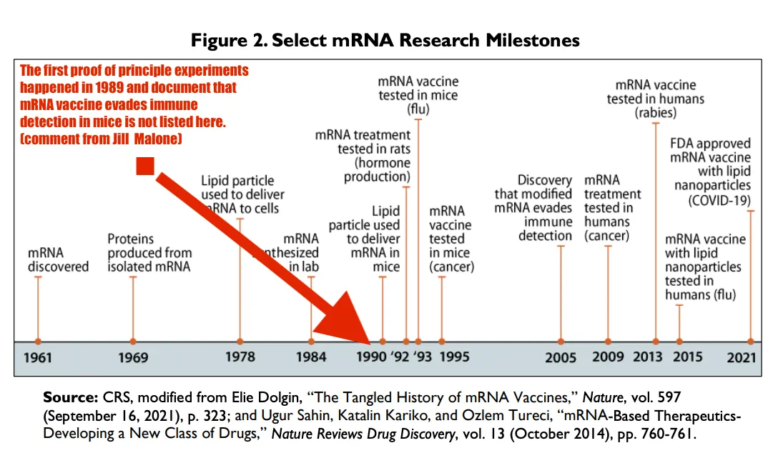
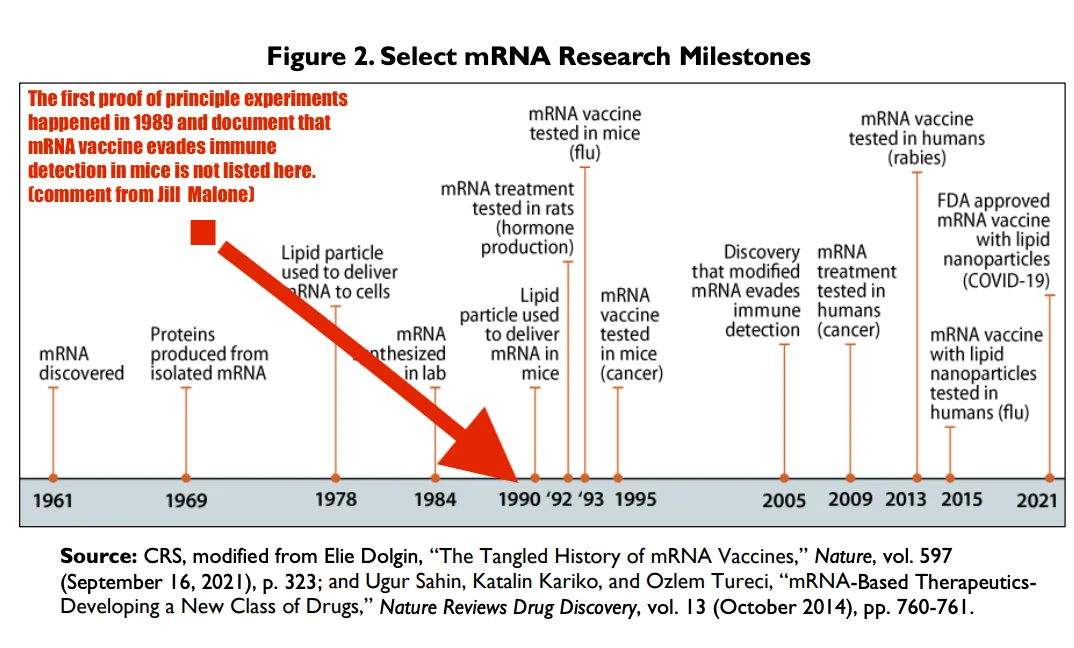
以下は、CRSが捏造した発明のバージョンである。USPTOによって文書化され公開された最初の原理証明の実験が、CRSの文書にあるこの図から欠落していることに注意してほしい。
CRSは、以下の彼らの出版物から、図に記録されているような原理証明実験を完全に書き抜いている。これは現在、米国議会の記録、つまり米国政府の出来事に関する記録である。
では、正しい歴史とは何だろうか?
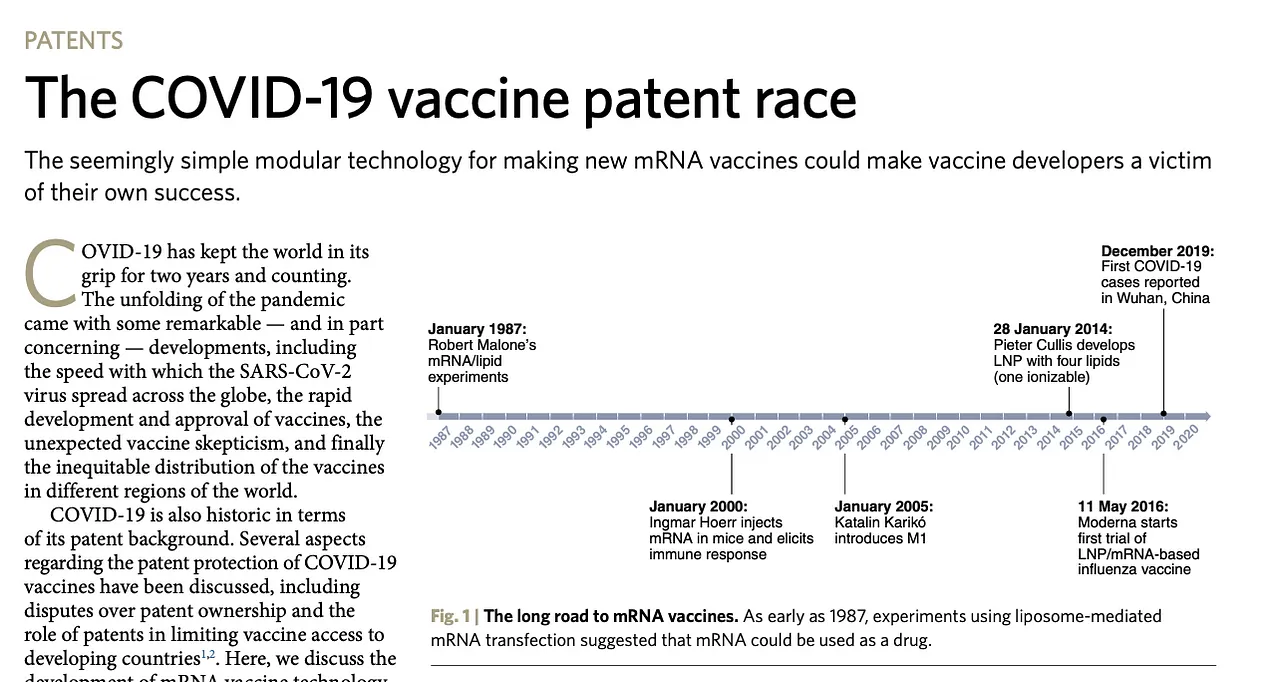
mRNAワクチンの歴史は、ロバート・マローン博士が1987年と1988年に大学院生としてソーク研究所に在籍していたときに始まった。そこで彼は、in-vitro RNAトランスフェクション、さらにin-vivo RNAトランスフェクション(カエルの胚やマウスを使ったもの)のパイオニアとなったのである。
その結果、彼の代表的な論文となった。カチオン性リポソームを介したRNAトランスフェクション RW Malone, PL Felgner, IM Verma. 米国科学アカデミー紀要(PNAS)86 (16), 6077-6081
このScienceの論文は、生体内試験のDNAとRNAのデータを並べて示した最初のものである(生体内試験のDNAに関する最初の論文)。その論文は1990年に発表された。
マウスの筋肉への生体内試験での直接遺伝子導入。Wolff JA, Malone RW, et al. Science. 1990;247(4949 Pt 1):1465-8. 4,750の論文に引用された、その研究成果である。
Malone博士が出願した特許とSalkからの開示資料には、in-vivo RNAトランスフェクションや、mRNAの安定化のための方法(現在、他の発明としてクレームされている)が含まれている。これらは、博士のウェブサイトで閲覧することができる。
彼の研究は1989年にVical社で継続され、最初の生体内哺乳類ラット実験が彼によってデザインされた。mRNA、コンストラクト、試薬はSalk研究所とVical社でMalone博士が開発し、これには生体内実験のための投与量も含まれていた。RNAとDNAはFedexでDr. Jon Wolffに送られた。ウィスコンシン大学のWolff博士は、マウスとラットに注射をした。RNAとDNAのワクチン接種に関する最初の特許開示は、1988年から1989年にかけてMalone博士によって書かれたものである。
この一連の研究は、9件以上の特許と多数の出版物を生み出し、この特許の研究は約7000件引用された。
1989年には、mRNAワクチンとトランスフェクションに関する9件のブレイクスルー特許を生み出す研究が行われ、その優先日はすべて1989年3月21日だった。これは、ソーク特許の出願と同じ優先日であり、2つの研究機関の弁護士が協力していたことがわかる。これらの特許は、mRNAワクチン接種に関する最初の公開研究である。特許のタイトルとリンクは、以下の文献に記載されている。これらの特許には、mRNAワクチンに関する原理実証実験があり、この発明が機能したこと、そしてこれがそれを示す最初の実験であることが明確に記録されている。
明らかに、不都合な歴史は、政府、大手製薬会社、新聞社、そしてネイチャーのような雑誌でさえ、一般の人々にその全歴史を知られたくないものである。 しかし、最近のネイチャー・バイオテクノロジー誌が書いたことは衝撃的に不正確であり、ネイチャー誌がこれらのワクチンを誰が発明したかを改竄したのはこれが初めてではない。
発見とアイデアが哺乳類で実用化された最初のmRNA実験-実際に発行され公開された特許に示されている-は以下にリンクされている。
以下は、発行された9つの特許の中の原理証明実験(例8と9)の文章だ。発明の詳細な説明は、特許の中に含まれている。 Dr. Maloneは、この技術を発明し、特許明細書を書き、初期の実験を主導し、それに関与してきた。Vical社を辞めた後も、Gary Rhodes博士とこの技術に関する共同研究を続けている。 つまり、米国政府、『ネイチャー』誌、さまざまな新聞、雑誌が現在歴史として書いていることは、真実ではないのだ。この特許には、他に4人の発明者がいて、実験に貢献している。 しかし、実際の発明とアイデアはマローン博士によるものであり、そのことを示す膨大な文書がある。 ジル・マローン博士が書いた詳細な歴史は、ここをクリック。
余談だが、この特許の所有者であるVical社という会社は、この仕事、原理証明の実験は発表しないことに決めていた。 その代わり、Vical社はこの技術をMerck社にライセンスし、Merck社はmRNA技術を放棄し、DNAワクチンの開発に取り組んだが、あまり成功しなかった。 発明者たちは、バイカルの雇用契約下にあったため、バイカルの許可を得ずに研究成果を発表することはおろか、それについて話すことさえできなかった。
以下は、特許に記載された最初の実験のうちの2つで、優先日は1989年である。
実施例8:HIVウイルスのgp120タンパク質を産生するためのマウスのmRNAワクチン接種
HIVウイルスのgp120タンパク質をコードするmRNAを含むリポソーム製剤は、gp120の遺伝子(Aids Research and Reagent Program, National Institute of Allergy and Infectious Disease, Rockville, Md. 20852からのpIIIenv3-1)が、例4の手順でプラスミドpXBGに挿入される以外は例1〜5に従って調製される。実施例6に従って調製し、10%ショ糖中の200μg/mlのgp120 mRNAおよび500μg/mlの1:1のDOTAP/PEを含む製剤の200alの容量を、1日に3回マウスの尾静脈に注射する。最後の注射から約12〜14時間後に、注射部位から筋肉の一部を取り出し、実施例7に従って、細胞溶解液として調製する。HIV特異的タンパク質gp120は、実施例7の手順に従って、ライセートにおいても同定される。
mRNAワクチン接種マウスの血清中に存在するgp120抗体がHIV感染を防御する能力は、以下のようにHT4-6Cプラーク減少アッセイによって決定される。HT4-6C細胞(CD4+ HeLa細胞)は、ブルース・チェスブロ博士、(ロッキーマウンテン国立研究所、モンタナ州)から入手し、RPMI培地(BRL、ゲイサーズバーグ、メリーランド州)中で培養して成長させる。次に、この細胞群をバッチに分割する。一部のバッチは、約107個のHT4-6C細胞に約105〜106個の感染単位のHIVを加えることによりHIVに感染させる。他のバッチは、HIVとgp120 mRNAを接種したマウスの血清約50μlの両方を加えることによって、HIV感染に対するgp120免疫血清の保護効果を試験する。3日間のインキュベーションの後、全てのバッチの細胞を洗浄し、固定し、クリスタルバイオレットで染色し、プラークの数をカウントする。gp120免疫血清の保護効果は、HIV単独で処理したバッチにおける数と比較して、gp120 mRNAワクチン接種マウス血清とHIVの両方で処理した細胞のバッチにおけるプラークの数の減少として決定される。
実施例9:nef mRNAによるヒト幹細胞保有SCIDマウスのmRNAワクチン接種とそれに続くHIVチャレンジ
重症複合免疫不全マウス(SCIDマウス(Molecular Biology Institute, (MBI), La Jolla, Calif. 92037))を、Mosierの方法(Mosier et al., Nature 335:256 (1988))に従って腹腔内注射により成人ヒト末梢血リンパ球と再構成した。その後、400〜4000感染単位のHIV-1を腹腔内注射した。マウスは、P3レベルの動物封じ込め施設において、密閉されたグローブボックス内で維持された。
HIVのnefタンパク質をコードするmRNAは、プラスミド(pGM92、NIAID、Rockville、Md.20852から)の形態でnef遺伝子を入手し;プラスミドからnef遺伝子を除去し;転写のためにpXBGプラスミドにnef遺伝子を挿入し;転写産物のnef mRNAを例2〜5で述べたように精製することによって調製された。次に、このnef mRNAを実施例6にしたがって製剤に組み込んだ。実験動物に、200μg/mlのNEF RNAと500μg/mlの1:1のDOTAP:DOPE(RNA/リポソーム複合体形態)を含む10%ショ糖溶液の200μl尾静脈注射を毎日行い、対照動物には同様に200μg/ml酵母tRNAと500μg/ml 1:1DOTAP/DOPEリポソームを含むRNA/リポソーム複合体を注射した。注射後2,4および8週目に、注射したリンパ系器官から生検標本を得て、免疫組織化学のために調製した。同じ時点で、血液サンプルを得て、ELISAキット(Abbott Labs, Chicago, Ill.)によりp24レベルについて、および実施例8のプラークアッセイによりウイルス力価についてアッセイした。HIV-1に対する免疫染色は、HIV感染患者からのポリクローナル血清を使用して、記載されているように(Namikawaら、Science 242:1684(1988))実施した。陽性細胞を数え、高倍率視野(400×)あたりの感染細胞数を決定した。これらのアッセイを用いると、8週間後には陽性染色細胞数の少なくとも2倍の減少が観察され、力価およびp24発現は少なくとも50%減少していた。これらの結果を合わせると、(生体内試験)処理による中程度の抗ウイルス効果があることがわかる。nef mRNAを200μg/ml、10%ショ糖中の1:1 DOTAP:DOPEを500μg/ml含む製剤を、ヒト幹細胞含有SCIDマウスの尾静脈に1日に3回注入する。免疫化後、マウスは有効量のHIVウイルスで感染させることによりチャレンジされる。尾静脈から定期的に血液サンプルを採取し、ELISAキットアッセイ(Abbott Labs, Chicago, Ill.)により、特徴的なHIVタンパク質p24の産生をモニターする。
以下は、発行された9件の特許である。
- 生物学的に活性なペプチドを送達し、細胞性免疫応答を誘導するための脂質媒介ポリヌクレオチド投与(mRNAを含む)。P Felgner, JA Wolff, GH Rhodes, R Malone, D Carson. Vical, Incに譲渡され、Merckにライセンスされた。No. 7,250,404, 発行日:2007/7/31 引用数:105件優先日:1989年3月21日。
- 被験者が感染する可能性を減少させるための脂質媒介ポリヌクレオチド投与(mRNAを含む)。P Felgner, JA Wolff, GH Rhodes, Robert W Malone, D Carson. Vical, Incに譲渡され、Merckにライセンスされた。US Pat. Ser.No.6,867,195 B1。発行日:2005年3月15日優先日:1989年3月21日。
- 病原体に対する免疫反応の生成(mRNAを含む)。P Felgner, JA Wolff, GH Rhodes, Robert W Malone, D Carson. Vical, Inc.に譲渡され、Merckにライセンスされた。US Pat. Ser.No.6,710,035. 発行日:2004年3月23日引用文献 39件の論文。優先日:1989年3月21日。
- 脊椎動物、哺乳類、魚類、鳥類またはヒトにおける外来性ポリヌクレオチド配列の発現(mRNAを含む)。P Felgner, JA Wolff, GH Rhodes, Robert W Malone, D Carson. Vical, Incに譲渡され、Merckにライセンスされた。 US Pat. Ser.No.6,673,776. 発行日:2004年1月6日優先日:1989年3月21日。
- 生理学的に活性なポリペプチドを哺乳類に送達する方法(mRNAを含む)。P Felgner, JA Wolff, GH Rhodes, Robert W Malone, D Carson. Vical, Incに譲渡され、Merckにライセンスされた。US Pat. Ser.No.6,413,942。発行日:2002年7月2日。(150の論文に引用)。優先日:1989年3月21日。
- DNA配列(mRNAを含む)の注射による哺乳動物における保護的免疫応答の誘導。P Felgner, JA Wolff, GH Rhodes, Robert W Malone, D Carson. Vical社に譲渡され、Merck社にライセンスされた。US Pat. Ser.No.6,214,804, 発行日: 2001年4月10日。360の論文に引用される。優先権主張日:1989年3月21日。
- DNA配列(mRNAを含む)の注射による哺乳類の保護的免疫応答の誘導。P Felgner, JA Wolff, GH Rhodes, Robert W Malone, D Carson. Vical, Incに譲渡、Merckにライセンス。 US Pat. Ser.No.5,589,466. 発行日:96年12月31日。899の論文に引用される。優先日:1989年3月21日。
- 哺乳類における外来性DNA配列の送達(mRNAを含む)。 Vical, Incに譲渡され、Merckにライセンスされた。P Felgner, JA Wolff, GH Rhodes, Robert W Malone, D Carson. US Pat. Ser.No.5,580,859. 発行日:96年12月3日。1244件の論文に引用される。優先日:1989年3月21日。
- 脂質を媒介としたDNAデリバリーによる抗体の生成(mRNAを含む)。P Felgner, JA Wolff, GH Rhodes, Robert W Malone, D Carson. Vical, Incに譲渡され、Merckにライセンスされた。US Pat. Ser.No.5,703,055. 発行日:1997年12月30日。419の論文に引用される。優先日:1989年3月21日。
- RNAトランスフェクションによるレトロウイルスRNAのパッケージングを研究するための新規アプローチ(要旨)。RW Malone、P. Felgner、I. Verma. RNA Tumor Viruses、1988年5月17-18日。Cold Spring Harbor (RNA transfection に関する一連の論文/要旨の最初のもの)。
- カチオン性リポソームを用いた培養真核細胞および胚の mRNA トランスフェクション。Malone RW. Focus. 1989; 11:61-8
- 1989年3月21日にUSPTOに提出されたSALK特許。カバーレターには、これを隠して、1989年3月29日に出願されたと書かれていることに注意。
- 最初のmRNA VACCINEの実験データ 1990年(VICAL社から特許庁へ)
- カチオン性リポソームを介したRNAトランスフェクション。Malone RW, Felgner PL, Verma IM. Proc Natl Acad Sci (PNAS) U S A. 1989;86(16):6077-81. 749の論文に引用されている。
- マウス筋肉への生体内試験直接遺伝子導入。Wolff JA, Malone RW, et al.科学. 1990;247(4949 Pt 1):1465-8. 4,750 件の論文に引用されている。なお、RobertはNorthwesternの学生であり、University of Wisc.とは無関係である。
- カチオン性リポソームを介したトランスフェクション組織培養細胞による高レベルのメッセンジャーRNA発現。Malone R, Kumar R, Felgner P. NIH Conference: 「抗HIV剤としての自己切断型RNA(要旨)。ワシントン DC 1989 年 6 月。
- カチオン性リポソームを介したRNAトランスフェクション。Dwarki VJ, Malone RW, Verma IM. Methods Enzymol. 1993;217:644-54. 引用された 102件の論文がある。
- 哺乳類における外来性DNA(mRNAを含む)配列のデリバリー P Felgner, JA Wolff, GH Rhodes, R Malone, D Carson. バイオテクノロジー・アドバンス 1993: 15 (3-4), 763-763
- 粘膜免疫反応を惹起するためのDNAワクチン(mRNAを含む)。US Pat. Ser.No.6,110,898。発明者発明者: RW Malone,Jill Glasspool Malone.発行日:8/29/00。40の論文に引用される。優先日:1996年。
- 活性なシトフェクチン:ポリヌクレオチドトランスフェクション複合体を生成するための製剤および方法 ロバート・W・マローン(Robert W Malone)、他、US Pat. Ser.No.5,925,623 7/20/99.