
コンテンツ
What Has Neuroimaging Taught Us on the Neurobiology of Yoga? A Review
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7362763/
要旨
ヨガは世界的に人気が高まっており、いくつかの身体的、精神的な効果が示唆されている。ここでは、ヨガの実践者を対象とした研究における既存の神経画像学的文献から生成された研究の包括的かつ批判的なレビューを提供する。
磁気共鳴画像法(MRI)陽電子放出断層撮影法(PET)単一光子放出コンピューター断層撮影法(SPECT)を用いたヨガに関する34の国際的な神経画像学的研究をレビューした:11の形態学的研究と26の機能的研究があり、そのうち3つは形態学的および機能的研究に分類されていた。
一貫した所見としては、島と海馬における灰白質体積の増加、前頭前皮質領域の活性化の増加、主にデフォルトモードネットワーク内での機能的接続性の変化などが挙げられる。神経画像所見にはかなりのばらつきがあり、これは部分的にはヨガのスタイルやアプローチの違いやサンプルサイズの制限を反映している。
身体活動のような直接的な比較対象グループは今のところほとんど使われていない。最後に、画像所見から導き出された根底にある神経生物学に関する仮説を、ヨガの潜在的な有益な効果に照らして議論する。
キーワード
ヨガ、神経イメージング、PET、SPECT、MRI、神経生物学、連結性
序論
ヨガという言葉は、サンスクリット語のyujに由来し、「結合 」または 「精神的な結合の方法 」を意味する。ヨガはインドで生まれたもので、サマディ(純粋な意識)を達成することを究極の目標とし、健康の改善と個人の変容を促進することを目的とした、肉体と精神のトレーニングを統合した行動練習である。
パタンジャリの『スートラ』によると、ヨガは次の8つの側面(四肢)すなわちヤマ(倫理的指針、不道徳な行動からの禁欲)ニヤマ(自己規律)アーサナ(身体的姿勢)プラナヤマ(呼吸のコントロール)プラティハラ(感覚の引き出し)ダーラナ(集中)ディヤーナ(瞑想)そしてサマディ(純粋意識)を包括している(Villemure er al 2014, 2015; Birdee et al 2016; Eyre et al 2016; Khalsa et al 2016)。)
長い年月をかけて、ヨガの哲学やスタイルのバリエーションが何十も生まれていた。ヨガのスタイルの中には、身体的なワークアウトとして構成されているものもあれば、瞑想に重点を置いているものもある。瞑想は、心の中の思考の流れを集中させ、減速させる方法である(Brewer et al 2011)。瞑想のためのサンスクリット語であるディヤーナは、パタンジャリの経典によると、ヨガの八つの肢の一つであり、ヨガと瞑想が相互に関連しているという事実を強調している。いくつかの瞑想法はヨガに由来しており、ヨガと本質的に結びついている。また、瞑想に集中して準備する心を鍛えるために、身体の動きと呼吸法の練習が用いられ、ヨガのトレーニングのトライアンドから一緒に行われている(Birdee et al 2016)。
ヨガは東洋では何千年も前から実践されていたが、現在では西洋世界でも急速に人気と関心が高まっている(Ivtzan and Jegatheeswaran, 2015)。最近の国民健康インタビュー調査のデータによると、米国成人の14.3%が過去12ヶ月間にヨガを行ったことがあるとされている(Clarke et al 2018)。さらに、ヨガを行っている人は、女性、若年層、非ヒスパニック系白人、大学教育を受けている人、高収入の人、健康状態が良い人が多いとされている(Cramer et al 2016)。多くの健康な人にとっての目標は、健康的でアクティブなライフスタイルの一環として、フィットネスと柔軟性を達成し、日々のストレスを軽減し、エネルギーを向上させることである(Barnes et al 2008; Ivtzan and Jegatheeswaran, 2015; Cramer et al 2016)。西洋社会ではいくつかのヨガスタイルが実践されているが、多くのセンターではヨガのアーサナ(物理的なポーズ)に主眼が置かれているが(Ivtzan and Jegatheeswaran, 2015)ほとんどは上述した身体的なポーズ、呼吸法、瞑想の3つの要素を含んでいる。また、ヨガの練習には、純粋に肉体的なものからよりスピリチュアルなものまで、さまざまな意図がある。最近の研究では、ヨガを実践する動機はダイナミックであることが示されている。練習を続けることで、より肉体的なものからよりスピリチュアルなものへと、練習者が設定する意図にシフトがある(Ivtzan and Jegatheeswaran, 2015)。
一方で、ヨガは身体的・精神的な問題を抱えている人たちにも定期的に実践されている。Cramer et al 2016)は 2012年の国民健康面接調査の横断的データを用いて分析を行った。この調査では、ほとんどのヨガ実践者がヨガの実践から得たポジティブな結果を報告しており、主にストレスの減少、全体的な健康状態の改善、感情的な健康状態の改善、睡眠の改善、健康をコントロールする感覚の増加などを挙げている。これらの主観的な自己申告による効果に加えて、ヨガの有益な医学的・心理学的効果に関する臨床研究も増えてきている。ヨガの生理学的効果としては、心拍数や血圧の低下、筋力の増加などが挙げられる(Hagins et al 2014;Vardar Yağlı et al 2015;Chu et al 2016)。さらに、抑うつ状態、不安状態、ストレス状態の改善および痛みの状態の緩和は、いくつかの研究で実証されている(Woolery et al 2004;Cramer et al 2013;Danucalov et al 2013;Riley and Park et al 2015;Wieland et al 2017)。したがって、ヨガはまた、うつ病、不安、心的外傷後ストレス障害(PTSD)などの様々な中枢神経系(中枢神経系)障害だけでなく、統合失調症や認知機能の低下においても、補完的な補助的アプローチを提供する可能性がある(Vancampfort et al 2012;Cramer et al 2013;Riley and Park et al 2015;Brenes et al 2018)。ほとんどの場合、これらの効果は、ヨガ介入の前、中、後の自己申告のアンケートで測定されており、そのようなアプローチは偏りや主観が生じやすいかもしれない。非侵襲的で簡単にアクセスできる(バイオ)マーカー、例えば血圧、薬の使用、心拍数(変動性)は関心のある測定値として使用されていたが、まだ特定のものであることに変わりはない。
ヨガの神経生物学的基盤を解明することに関心が高まっている。人間の脳の神経生物学を非侵襲的に調査する方法には、磁気共鳴画像法(MRI)陽電子放出断層撮影法(PET)単光子放出コンピューター断層撮影法(SPECT)脳波検査(EEG)脳磁図検査(MEG)経頭蓋直流刺激法(TDCS)経頭蓋磁気刺激法(TMS)血液バイオマーカー分析など様々な方法がある。これらの非侵襲的モダリティは、高空間分解能(MRI)と高感度(PET)で生体内の脳の構造と機能、ネットワーク機能効果を調べることができるため、本レビューでは、MRをベースとした手法、PET、SPECTを用いた神経イメージング研究に焦点を当てている。
本レビューの主な目的は、報告されている大脳効果の大きさと一貫性を検討することで、批判的な観点からヨガの既存の構造的・機能的神経イメージング研究の概要を提供することである。我々は矛盾に対処し、現在の知識のギャップを強調し、より良い研究デザインとイメージングプローブがどのようにしてこれらに対処できるかを議論する。
生体内の脳イメージング技術
MRI、PET、SPECTは、ヨガの脳への影響を調査するために選択された医学的な神経画像技術である。
MRIは、強い磁場、磁場勾配、電波を利用して組織内の水素原子を励起し、これらの組織の特性に応じて信号を読み取るイメージング技術の一群を指する(Hendee and Morgan, 1984)。MRIの主な利点は、(サブ)ミリメートルの空間分解能、非侵襲性、組織の物理的・生化学的性質を利用して組織を識別できることである。一方で、感度が比較的低いことが主な欠点である(Catana er al)。 MRIは、脳の構造的影響と機能的影響の両方を評価するために使用することができる。構造的MRI(sMRI)は、解剖学的情報を提供し、異なる組織や脳構造を線引きし、体積的変化や集団間の違いを評価するために使用することができる。脳の白質(脳白質)の変化の可能性を評価および調査するために、拡散テンソルイメージング(DTI)が一般的に使用されている(Sasson et al 2010)。DTIは水素の拡散特性に敏感である。拡散が制限されていない場合、水素は全方向に同じ量が拡散し、”等方性 “と呼ばれている。一方、水素の拡散が制限され、一方向に支配されている場合、拡散は “異方性 “と呼ばれる。脳白質では、軸索を取り囲む軸索壁やミエリン鞘によって拡散が異方的になる傾向があり、それをDTIで拾うことができるため、脳白質路の完全性を調べるのに適している。sMRIに加えて、安静時fMRI、タスクfMRI、機能的接続性fMRI、磁気共鳴分光法(MRS)、動脈スピンラベリング(ASL)などの機能的MRI(fMRI)測定が可能である。In vivo MRSはMRIに付随する非侵襲的な特殊技術であり、様々な化学物質の共鳴周波数の違いを測定・観察してMRSスペクトルを同定する。このMRSスペクトルは、関心領域(またはボクセルベースの測定による脳全体)の生化学的組成を反映しており、各代謝物はその固有の位置によって同定される(Henning, 2018)。GABA、主要な抑制性および興奮性神経伝達物質を反映するグルタミン酸+グルタミン(Glx)細胞膜の合成および破壊のマーカーであるグリセロホスホコリン+ホスホコリン(Cho)および神経細胞および軸索の生存率および密度のマーカーであるN-アセチルアスパラギン酸およびN-アセチルラスパーティルグルタミン酸(NAA/NAAG)である(Verma et al 2016)。安静時またはタスクfMRIでは、ニューロンの活動状態に応じて変化する脳組織の血中酸素濃度依存性(BOLD)変化に基づいて、脳の局所活動が検出される(Chen and Glover, 2015)。活性化された領域の相関パターンは、機能的接続性解析により同定することができる。いくつかのネットワークがロバストに同定されており、その中には、注意を必要とするタスクがないときに活性化するデフォルトモードネットワーク(DMN)(Fox and Greicius, 2010)や、中央幹部(前頭)ネットワーク(Trotta et al 2018)などがある。間接的なニューロン機能は、絶対的な脳灌流を測定するASLを用いて測定することができる。この手法では、流入動脈血は磁気的に標識され、標識された対照画像と標識されていない対照画像の間の信号差は、局所的な灌流に比例する(Haller et al 2016)。
PETは、非常に高い(ナノモル)感度を有する分子機能核医学イメージング技術である。PETイメージングは、トレーサー量で投与された放射性標識放射性リガンドを利用している。陽電子放射性ラジオアイソトープからなる放射性物質は、神経受容体、再取り込み輸送体、細胞内・細胞外タンパク質などの標的分子に結合する化合物に結合している。陽電子放出は、シンチレータまたは固体PET-scan検出器で検出された同時消滅ガンマ線の測定により位置が特定され(Sossi, 2018)受容体密度やグルコース代謝率などの定量的な生理的指標が得られる。空間分解能はsMRIに劣るが、現在の装置では3〜4mmの空間分解能が可能である(Vandenberghe et al 2016)。
同様に、SPECTイメージングは、分子的および機能的な脳のプロセスを可視化するためにも使用される。PETとは対照的に、トレーサーは単光子放出放射性同位体で標識される。SPECTイメージングは、回転シンチレータガンマカメラを用いて、放射性トレーサー分布の定量的な画像を提供する。脳機能研究では、SPECTは主に脳の局所的な灌流を研究するために使用されていた(Goffin and van Laere, 2016)。
レビュー方法と検索戦略
MEDLINE1を用いて2019年末までの論文を特定し、以下のキーワードでヨガと神経画像研究に限定して検索した。”yoga AND (magnetic resonance OR positron emission tomography OR single photon emission computed tomography OR MR OR PET OR SPECT or functional connectivity)” 得られた論文リストは、その後、要約に「磁気共鳴 OR MR」、「機能的磁気共鳴イメージング OR fMRI」、「陽電子放出断層撮影 OR PET」、「単一光子放出コンピュータ断層撮影 OR SPECT」、またはタイトルまたは要約に「脳」と「ヨガ」のうち少なくとも一つを含むかどうかを精査した(下記の研究の包含基準と除外基準を参照)。残りの結果のうち、すべての要旨
は、ヨガの脳への影響を調査するための神経画像法を記述していることが確認された。
異なるヨガスタイルの形態を調査するために構造的・機能的神経イメージングを用いた研究で、以下の包含基準を満たしているものはすべて検討した。(i) 英語、ドイツ語、オランダ語、フランス語で掲載された論文、(ii) 神経画像法を用いた論文、(iii) 査読付き科学雑誌に掲載された論文。
研究は、(i)個別の症例報告、(ii)一回限りの施術者の効果、(iii)レビュー論文、(iv)MRI、PET、SPECT以外の神経生理学的手法(例えば、脳波マッピング、MEGなど)を使用した場合は除外した。
研究成果
研究の選択と分類
合計93件の研究が特定され、レビューされた。すべての研究のうち約3分の2(93件中59件)は含まれなかったが、主にヨガそのものに焦点を当てていなかったり、神経細胞への影響を調査していなかったりしたため、最終的には34件の研究が含まれてた。
含まれた研究では異なるスタイルのヨガが用いられており、で簡単に説明されている。アシュタンガ、アイアンガー、ヴィンヤサ、クリパル、クンダリーニ、ニドラ、サハヤ、シバナンダ、ハタヨガである。選ばれた34論文は、sMRIによる脳の構造変化を評価する「構造・形態学的ニューロイメージング研究」(n=11件、表1)と、脳機能、活性化研究、分子標的を評価する「機能的ニューロイメージング研究」(n=26件、表2)に簡単に分類された。後者には、MRS、タスクベースfMRI、安静時fMRI(rsfMRI)接続性解析を伴うASL、PET、SPECTが含まれた。34本の論文のうち3本は構造画像と機能画像を併用しており、両セクションに含まれていた。6組の論文は同じ被験者で異なる解析を行っており、構造的研究と機能的研究の両方のセクションに含まれていた。
表1 ヨガに関する形態学的脳イメージング研究
表1
ヨガに関する形態学的脳画像研究。
| イメージング技術 | 効果 | 設計 | ヨガスタイル | 数(♀)、年齢(平均±SD)、年 | 対照群対照条件 | 調査 |
| MRI-VBM | WB 灰白質密度 | CSS | ハタヨガ瞑想(HYM)(MY) | HYM:7(6)、36.4±11.9 CON:7(6)、35.5±7.1 | 非瞑想者対ヨガ | Froeliger et al。、2012b |
| MRI-VBM MRI-VBCT MRI-DTI | WB 灰白質密度、灰白質T、ROI 脳白質 | CSS | 不特定のヨガ(3年) | YOG:14(9)、37.0±6.6 CON:14(9)、36.7±7.3 | 非瞑想者対ヨガ | Villemure et al。、2014 |
| MRI-VBM | WB 灰白質密度 | CSS | 不特定のヨガ(3年) | YOG:14(9)37.0±6.6 CON:14(9)36.7±7.3 | 非瞑想者対ヨガ | Villemure et al。、2015 |
| MRI-VBM | WB 灰白質密度 | CSS | サハジャヨガ瞑想(SYM)(YM) | SYM:23(17)、46.5±11.4 CON:23(17)、46.9±10.9 | 非瞑想者対ヨガ | エルナンデス他、2016年 |
| MRI-VBM | WB 灰白質密度 | LS | サハジャヨガ瞑想(SYM)(MY) | SYM:12(2):21.6±2.0 CON:30(12):22.2±1.3 | SYMトレーニングvs.コントロールグループ(待機期間) | Dodich et al。、2019 |
| MRI-VBCT | WB 灰白質T | CSS | ハタヨガ(3年) | YOG:21(21)、66.2±4.5 CON:21(21)、67.9±4.6 | 非瞑想者対ヨガ | Afonso et al。、2017 |
| MRI-VBM | ROI灰白質V | CSS LS | 瞑想とヨガの練習(MY / 3Y) | YOG:289(211)、61.9±6.8 CON:3453(1830)、64.3±7.7 | ヨガ、瞑想、呼吸法のコントロールと実践者 | Gotink et al。、2018 |
| MRI-VBM | ROI灰白質密度 | LS | ヨガ(3年) | ヨガ:7(3)、69–81 * | ヨガトレーニングの前と後 | Hariprasad et al。、2013 |
| MRI-VBM | ROI灰白質密度 | CSS | ハタ、アイアンガー、クンダリーニヨガ(3年) | YOG:13(12)、35.8±15.4 CON:13(12)、35.7±14.6 | コントロール対ヨガ施術者 | Gothe et al。、2018 |
| MRI-VBM | ROI灰白質密度 | LS | MBSRプログラム(ヨガを含む)(MY / 3Y) | ヨガ:27(16)、35.2±6.7 | MBSRトレーニングの前と後 | Hölzeletal。、2009 |
| MRI-VBM | ROI灰白質V | LS | 瞑想とクンダリーニヨガの組み合わせ(MY / 3Y) | YOG:14(6)、67.1±9.5 CON:11(6)、67.8±9.7 | MCI患者におけるMET対ヨガ | ヤン他、2016年 |
MRI-VBM ROI 灰白質V LS 瞑想とクンダリーニヨガの組み合わせ(MY/3Y) YOG:14(6)67.1±9.5 CON:11(6)67.8±9.7 MCI患者におけるMET vs.ヨガ Yang et al 2016年
イメージング技術。DTI、拡散テンソルイメージング、MRI、磁気共鳴イメージング、VBM、ボクセルベースの形態計測、VBCT、ボクセルベースの皮質厚さ 効果。灰白質D、灰白質密度、灰白質V、灰白質体積、灰白質T、灰白質厚、灰白質、灰白質、ROI、関心領域、WB、全脳、脳白質、白質。デザイン。CSS、横断的研究、LS、縦断的研究。ヨガスタイル:3Y、ヨガのトライアッド(身体的なポーズ、呼吸法、瞑想を含む)HYM、ハタヨガ瞑想、MBSR、マインドフルネスに基づくストレス軽減、MY、瞑想ヨガ、SYM、サハヤヨガ瞑想。数。SD、標準偏差。対照群 対照条件。MCI、軽度認知障害、MET、記憶力強化トレーニング。* 平均年齢、SDについては情報なし。
表2 ヨガに関する脳機能イメージング研究
表2
ヨガに関する機能的脳イメージング研究。
| イメージング技術 | 効果 | 設計 | ヨガスタイル | 数(♀)、年齢(平均±SD)、年 | 対照群対照条件 | 調査 |
| A.脳灌流と糖代謝 | ||||||
| 18 F-FDGPET | 糖代謝 | CSS | ヨガ瞑想(YM) | ヨガ:8(2)、32、21–39 * | 同じ被験者の覚醒状態 | Herzog et al。、1991 |
| H 2 15 O PET | CBF | CSS | ヨガニドラ(YM) | ヨガ:9(3)、23–41 * | 同じ被験者のコントロール状態 | Lou et al。、1999 |
| 99m Tc-ECD SPECT | CBF | LS | アイアンガーヨガ(3年) | ヨガ:4(2)、45.0 | プログラム前のベースラインスキャン | コーエン他、2009年 |
| 99m Tc-HMPAO SPECT | CBF | CSS | クンダリーニの詠唱(YM) | ヨガ:11(5)、35.4±13.5 | 同じ科目のBL | Khalsa et al。、2009 |
| fMRI ASL | CBF | CSS | クンダリーニ瞑想(YM) | ヨガ:10(4)、53.7 * | 同じ科目のBL | Wang et al。、2011 |
| B.神経の活性化 | ||||||
| fMRI | 神経の活性化 | CSS | クンダリーニ、アセムの伝統(MY) | ヨガ:8(5)、34.6±9.7 | 同じ科目のBL | Engströmetal。、2010 |
| fMRI | 神経の活性化 | CSS | OM詠唱(MY) | ヨガ:12(3)、28±6 | 「ssss…」の制作 | Kalyani et al。、2011 |
| fMRI | 神経の活性化 | CSS | サハジャヨガ瞑想(MY) | ヨガ:19(11)、46.6±9.5 | 呼吸への注意 | Hernándezetal。、2015 |
| fMRI | 神経の活性化 | CSS | パタンジャリヨガ:ヨガ瞑想(MY) | ヨガ:4(0)、60代半ば* | リラクゼーション(対照条件)対瞑想 | ミシュラ他、2017年 |
| fMRI | ニューロンの変動 | LS | ハタヨガ(3年) | CON:12(12)、16–60 * YOG:23(23)、16–60 * AE:23(23)、16–60 * | 統合失調症の患者:コントロール待機リストvs.ヨガvs.有酸素運動(AE) | Lin et al。、2017 |
| fMRI | 神経の活性化 | CSS | 未定義 | YOG:19(16)、35.9±11.5 CON:12(6)、32.9±9.1 | レクリエーションアスリートvs.ヨガプラクティショナー(YP) | Wadden et al。、2018 |
| fMRI | 神経の活性化 | CSS | ハタ瞑想(MY) | YOG:7(6)、36.4±11.9 CON:7(6)、35.5±7.1 | 瞑想ナイーブ対コントロール | Froeliger et al。、2012c |
| fMRI | 神経の活性化 | CSS | ハタ、アイアンガー、クンダリーニ(MY / 3Y) | YOG:13(12)、35.8±15.4 CON:13(12)、35.7±14.6 | コントロール対ヨガ施術者 | Gothe et al。、2018 |
| C.機能的な接続 | ||||||
| fMRI | 機能的な接続 | CSS | ハタ瞑想(MY) | YOG:7(6)、36.4±11.9 CON:7(6)、35.5±7.1 | 瞑想ナイーブ対コントロール | Froeliger et al。、2012a |
| fMRI | 機能的な接続 | CSS | サハジャヨガ瞑想(SYM)(YM) | SYM:23(17)、46.5±11.4 CON:23(17)、46.9±10.9 | 瞑想状態と安静状態 | Hernándezetal。、2018 |
| fMRI | 機能的な接続 | LS | MBSRプログラム(ヨガを含む)(MY / 3Y) | MBSR:18(10):37.5±9.1 RR:16(9):39.9±10.3 | MBSR対リラクゼーション応答RRトレーニング | Sevinc et al。、2018 |
| fMRI | 機能的な接続 | CSS | ハタヨガ(3Y) | CON:20(20)、68.2±4.6 YOG:20(20)、66.5±4.5 | 高齢者のヨガ実践者と健康的なヨガ未経験のコントロール | Santaella et al。、2019 |
| fMRI | 機能的な接続 | LS | 瞑想/クンダリーニヨガ(MY / 3Y) | ヨガ:14(6)、67.1±9.5 MET:11(6)、67.8±9.7 | MCI患者におけるMET対ヨガ | Eyre et al。、2016 |
| fMRI | 機能的な接続 | LS | サハジャヨガ瞑想(SYM)(MY) | SYM:12(2):21.6±2.0 CON:30(12):22.2±1.3 | SYMトレーニングvs.コントロールグループ(待機期間) | Dodich et al。、2019 |
| fMRI | 機能的な接続 | CSS | クリパルヨガ、ヴィパッサナー瞑想(MY / 3Y) | YOG:16(11)、49.4±7.8 MED:16(10)、54.1±8.1 CON:15(9)、52.9±9.8 | 瞑想vs.ヨガvs.コントロール | Gard et al。、2014 |
| D.脳の代謝物と神経伝達物質 | ||||||
| 11 C-ラクロプライドPET | ドーパミン放出 | CSS | ヨガニドラ(MY) | ヨガ:8(0)、31–50 * | 同じ被験者の覚醒状態 | Kjaer et al。、2002 |
| MRI-MRS | GABA | CSS | ヨガ(3年) | YOG:8(7)、25.8±5.2 CON:11(5)、26.6±7.6 | 読書運動 | ストリーター他、2007年 |
| MRI-MRS | GABA | LS | アイアンガーヨガ(3年) | YOG:19(11)、23.9±3.0 CON:15(11)25.6±4.9 | ウォーキンググループ | ストリーター他、2010年 |
| MRI-MRS | NAAとMI | CSS | ヨガのポーズと呼吸法(3年) | YOG:34(?)、35–65 * CON:34(?)、35–65 * | 2型糖尿病患者:ヨガ+標準治療vs.標準治療 | Nagothu et al。、2015 |
| MRI-MRS | NAAとMI | CSS | ヨガのポーズと呼吸法(3年) | YOG:5(?)、35–55 * CON:5(?)、35–55 * | 2型糖尿病患者:ヨガ+標準治療vs.標準治療 | サンタクマリ他、2016年 |
| MRI-MRS | 代謝物 | LS | 瞑想/クンダリーニヨガ(MY / 3Y) | YOG:14(6)、67.1±9.5 CON:11(6)、67.8±9.7 | MCI患者におけるMET対ヨガ | ヤン他、2016年 |
イメージング技術。ASL, arterial spin labeling; FDG, fluorodeoxyglucose; (f)MRI, (functional) magnetic resonance imaging; MRS, magnetic resonance spectroscopy; PET, positron emission tomography; SPECT, single-photon emission computed tomography. 効果。脳血流、脳血流;GABA、γ-アミノ酪酸;MI、ミオイノシトール;NAA、N-アセチルアスパラギン酸。デザイン。CSS、横断的研究; LS、縦断的研究。ヨガスタイル:3Y, triad of yoga(身体的なポーズ、呼吸法、瞑想を含む);MBSR, mindfulness-based stress reduction;MY, meditation yoga。数値。SD、標準偏差。対照群/対照条件。MET、記憶強化トレーニング。* 平均年齢およびSDに関する情報はない。
ヨガに関する構造/形態学的神経画像学的研究
2009年から 2019年の間にヨガを行った人を対象に実施されたすべての構造研究では、ボクセルベースの形態測定(VBM)解析が行われた。VBMは、事前に定義された関心領域(ROI)における灰白質(灰白質)体積、または脳全体の濃度(密度)の違いをボクセル単位またはROI単位で定量的に評価する技術である(Hutton et al 2009)。さらに、ボクセルベースの皮質厚(VBCT)分析を実行することができ、結果として得られる脳白質-灰白質表面と灰白質-脳脊髄液表面データの間の距離を計算することによって皮質厚を評価する(Lüsebrink et al 2013)。
いくつかの研究では、特定のROIに事前に制限することなく、全脳解析を行って形態学的効果を調査している。Froeliger et al 2012b)は、経験豊富なハタヨガ瞑想者の小グループ(n = 7)では、性別、年齢、学歴をマッチさせた対照群(n = 7)と比較して、灰白質密度が高いことを発見した。特に、内側前頭回、上前頭回、前頭前野、(パラ)海馬回、島、上側頭回、後頭回、小脳で高い灰白質密度が認められた(Froeliger et al 2012b)。
同様に、Villemure et al 2014)は、経験豊富なヨガ実践者(n = 14)と身体的に活動的な対照者(n = 14; 性別、年齢、体格指数、手の不自由さ、教育、ヨガ以外の運動レベルでマッチさせた)のグループを対象に行った調査でも、皮質領域の灰白質密度が高いことを確認した。これらの領域は、帯状回、上前頭前野、下頭頂葉、島皮質で構成されていた。また、VBCTでは、ヨガ参加者で灰白質厚が有意に増加した3つの脳領域、すなわち帯状回皮質、島皮質、一次体性感覚皮質がVBM解析と一致した(Villemure et al 2014)。
同じ被験者を対象とした追跡研究では、著者らはまた、ヨガの練習が神経保護効果を持つかどうかを、加齢に伴う灰白質の低下を比較することで探った(Villemure et al 2015)。その結果、対照群では年齢と全脳灰白質密度の間に有意な負の相関が見られ、健康的な加齢による萎縮効果が知られていることと一致したが、この低下はヨガ実践者には見られなかった。さらに、ヨガの経験年数と週1回のヨガ練習時間は、地域的な灰白質密度と正の相関があった(Villemure et al 2015)。
サハヤヨガ瞑想経験者(n = 23)では、対照者(n = 23)と比較して、有意に大きな灰白質密度が、島皮質、前頭前野前庭、内側下側頭回に認められた(Hernández et al 2016)。
Dodich et al 2019)は、同じヨガスタイルを実践している被験者を対象に、健康な瞑想未経験の成人42人をコントロール群(ウェイティングリスト;n=30)またはヨガ群(n=12)のいずれかに無作為に割り付けて、短期のヨガ瞑想トレーニング(1週間に1時間のセッションを4週間連続で週4回)の変調を調査した。対照群と比較して、ヨガ群では下前頭回(眼窩前庭)の灰白質密度が増加しており、ヨガトレーニング後の一般的な幸福感と相関していた。
最後に、別の研究では、Afonso et al 2017)は、年齢、教育、身体活動をマッチさせた対照群21人と比較して、経験者(週2回以上、最低8年間練習)の健康な高齢女性ハタヨガ実践者21人の左前頭前野の皮質の厚さの増加を発見した。
不安の軽減や注意力の向上などのヨガの行動効果とそれらの機能的解剖学的関係について定義された先験的仮説に基づいて、他の研究では海馬、扁桃体、または背側前帯状皮質を含む特定のROI内の効果に焦点を当てて構造的MRデータを分析している。Gotink et al 2018)は、瞑想とヨガが扁桃体(感情記憶、恐怖、不安の処理における中心的な中継構造として)と海馬に与える影響を調査した。3742人の参加者を含むこの大規模な集団ベースの研究では、289人が週に1時間以上の瞑想やヨガを1年以上実践した結果、左海馬と右扁桃体で灰白質量の減少が観察された(Gotink er al)。 この研究では、ヨガと瞑想の練習を区別していないため、観察された効果の正確な起源が不明瞭であることは言及する価値がある。
同じ研究では、5年前に前回のMRスキャンを受け、5年以上ヨガや瞑想を実践している218人の被験者のサブサンプルでの追加分析も記載されている。この縦断的研究では、ヨガ/瞑想の経験年数と扁桃体の体積の減少との間の相互作用のみが観察され、海馬の違いは認められなかった(Gotink et al 2018)。
同様の結果は、健康な高齢者被験者(n = 7)を対象に、対照群を設けずに実施したVBMを用いた小規模なヨガ介入研究(身体的なポーズ、呼吸制御運動、瞑想を含む)でも見られた(Hariprasad er al)。
6ヶ月間のヨガ介入後に、プリオリROIとして海馬の灰白質密度の変化を調べた。海馬は加齢に伴う灰白質の消失の影響を受けることが知られており、以前にも血清脳由来神経栄養因子(BDNF)の上昇がこの領域で実証されていた。また、ヨガ介入後の海馬の灰白質密度の増加が、ベースラインと比較して認められた(Hariprasad et al 2013)。
前者の結果に基づいて、(Gothe et al 2018)は、経験豊富なヨガ実践者13人と年齢と性別をマッチさせた対照者13人を対象に、特に海馬の灰白質密度の違いを探った。経験豊富なヨガグループには、ハタヨガ、クンダリーニヨガ、アイアンガーヨガの実践者が含まれていた。灰白質密度は、対照群と比較してヨガ群の左海馬で高かった(Gothe et al 2018)。
他の構造イメージング研究では、ストレスが上昇した被験者(n = 26)を対象に、週1回のグループミーティングと座位瞑想やヨガなどの毎日の家庭でのマインドフルネス実践からなる8週間のマインドフルネスに基づくストレス軽減(MBSR)介入の効果を調査するために、海馬と扁桃体の両方に焦点を当てた(Hölzel et al 2009)。知覚ストレス尺度の違いは、右扁桃体内の灰白質密度の変化に関連していた(Hölzel et al 2009)。
最後に、軽度認知障害(MCI)患者における記憶トレーニングまたはヨガ介入のいずれかの後の神経解剖学的および神経化学的可塑性を調査することを目的とした特定の研究では、(Yang et al 2016)は、12週間にわたって両側海馬および前帯状体後面内の灰白質体積を測定する縦断的MR研究を実施した。合計25名の被験者が含まれ、ヨガ群(n=14)と対照群[記憶強化トレーニング群(n=11)]のいずれかに無作為に割り付けられた。グループと時間の間に交互作用が見られた:記憶力強化トレーニングセッションを行ったMCI患者のみが、トレーニング後に背側前帯状皮質の体積が増加する傾向を示した(Yang et al 2016)。
脳白質完全性に対するヨガの効果を調査した研究は1件のみである。この研究では、Villemure et al 2014)は、経験豊富なヨガコホート(n = 14)の島皮質のROIベースのDTI分析を、対照群(n = 14)と比較して行った。彼らは対照群と比較して、左島皮質に隣接する脳白質でより高いフラクショナル異方性(FA)を発見した。確率論的トラクトグラフィを用いて、この領域を通過する脳白質経路を追跡したところ、前島領域と後島領域の連結性が増加していることが示された。著者らによると、これはヨガの被験者における島内接続性の増加を表している可能性がある(Villemure et al 2014)。
形態学的研究の研究明細を表1にプールした。さらに、図1Aは、含まれている研究でヨガの形態学的効果が報告されている異なる脳領域のマップを示している。島皮質と海馬は、形態学的に肥厚していると最も頻繁に報告されている領域でした。
図1 ヨガの形態学的および機能的効果の概要を、T1強調画像に重ねて示したもの
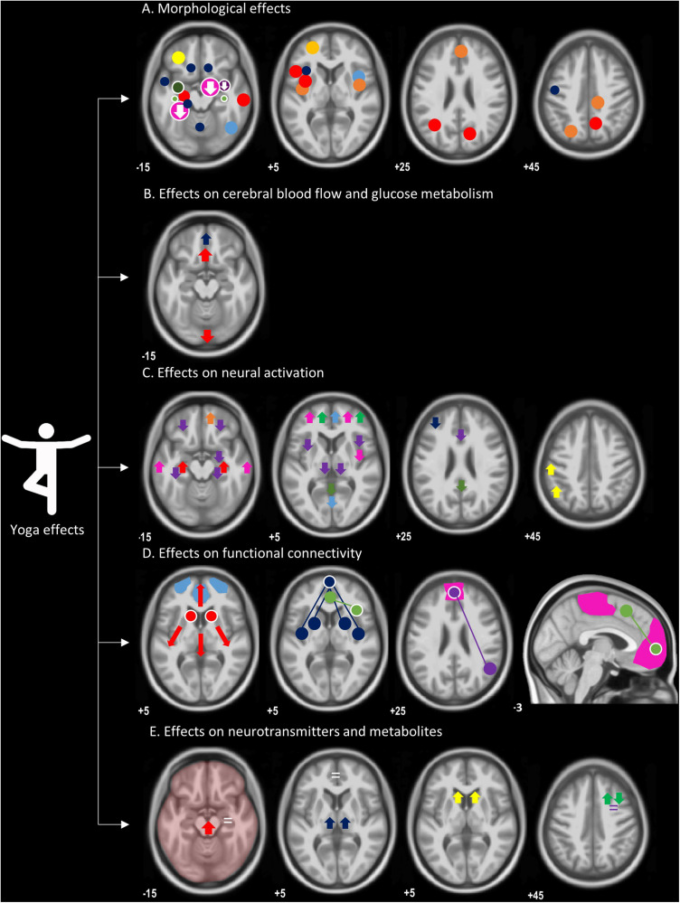
A)形態学的効果:円は灰白質の体積や密度が増加した領域を示し、減少した領域は下向きの矢印で示されている。全脳解析とROI解析の両方が含まれており、それぞれ白抜きまたは白抜きの円で表されている。円の大きさは、グループの大きさを表すものである:水色(Hernández et al 2016)紺色(Froeliger et al 2012b)オレンジ色(Villemure et al 2014)赤色(Villemure et al 2015)黄色(Dodich et al 2019)暗黄色(A. 2019)濃い黄色(Afonso et al 2017)ピンク(Gotink et al 2018)濃い緑(Gothe et al 2018)薄い緑(Hariprasad et al 2013)紫(Hölzel et al 2009)本図にはない(Yang et al 2016)。
B)神経静止状態機能:赤(Herzog et al 1991)青(Khalsa et al 2009)この図にはない(Lou et al 1999;Cohen et al 2009;Wang et al 2011)。
C)神経活性化:赤(Engström et al 2010)紫(Kalyani et al 2011)緑(Froeliger et al 2012c)ピンク(Hernández et al 2015)オレンジ(Mishra et al 2017)黄色(Wadden et al 2018)濃紺(Gothe et al 2018)水色(Lin et al 2015)。
D)機能的接続性、より高い機能的接続性を表す、脳領域(丸)と接続されたシード領域(白枠の丸)(全線):赤(Gard et al 2015);尾状核のより高い程度の中心性、青(Hernández et al 2018)緑(Hernández et al 2018)青(Hernández et al 2018)緑(Hernández et al 2018)。2018)緑(Sevinc et al 2018)ピンク(Eyre et al 2016)紫(Santaella et al 2019)水色は、パワースペクトルの変化を有するネットワークを表す(Dodich et al 2019)この図にはない(Froeliger et al 2012a)。
E)神経伝達物質および代謝物:黄色、ドーパミンシグナリングの増加(Kjaer et al 2002);赤、正確な位置を示さずにGABAの増加(Streeter et al 2007);紺色(GABAレベル)(Streeter et al 2010);緑、N-AGABAの増加(Streeter et al 2019);緑、N-AGABAレベルの増加(Streeter et al 2019)。2010);緑、N-アセチル-アスパラギン酸(NAA)の増加およびミオイノシトール(MI)の減少(Nagothu et al 2015);紫、MIまたはNAAに変化なし(Santhakumari et al 2016);白、海馬および背側前帯状皮質の代謝物レベルに変化なし(Yang et al 2016)。
ヨガに関する機能的神経イメージング研究
26の研究が神経画像法を用いてヨガの機能的効果を取り上げており、そのうち21の研究がMRベースの手法を用い、3つの研究がPETを用い(18F-フルオロデオキシグルコース(18F-FDG)によるグルコース代謝の評価、15O-H2Oによる脳血流や灌流の評価、11C-ラクロプリドによるドーパミン機能の評価を含む)残りの2つの研究がSPECT灌流を用いている。これらの研究を4つの一貫した領域にグループ化した。すなわち、安静時機能(主に)への影響(灌流とグルコース代謝)脳活性化研究、ネットワーク機能(連結性解析)分子変化(MRSと神経伝達物質)である。
ヨガの脳灌流・グルコース代謝への影響
ヨガ被験者における神経細胞の安静時機能の変化は、1つのグルコース代謝PET研究、1つの灌流PET研究、2つの灌流SPECT研究、1つの灌流ASL MR研究で調査されている(表2Aと図1B)。
Herzogら(1991)は、脳内グルコース代謝の可能性のある変化を見つけることを目的として、ヨガの被験者を対象とした歴史的に初めてのPETイメージング研究を行った。経験豊富な健康なヨガ瞑想ボランティア8名のグループを2日間のプロトコルで18F-FDG PETを用いてスキャンした。グルコースの絶対的な地域的な脳代謝率(rCMRGlc)は、ダイナミック18F-FDG PETイメージングと動脈血サンプリングで得られたが、有意差は認められなかった。前頭前野から後頭頂ROI、前頭前野から後頭頂ROI、上前頭前野から上頭頂ROIの比率でグルコース代謝が増加し、rCMRGlc比のいくつかの有意な局所差が認められた。さらに、ヨガ瞑想的リラクゼーションの状態では、変動の小さい被験者間係数が観察された(Herzog et al 1991)。
最初の脳灌流研究では、ルーら(1999)は、15O ・H 2 O PETを使用して9つの経験豊富な被験者のニードラヨガ瞑想を補助する神経領域を調べた。被験者は15O-H2Oで8回注入された:通常の安静状態の間に2つ、聴覚刺激と安静状態の間に2つ、および聴覚刺激によって誘導され、維持された瞑想中に4つの:。実験全体の脳血流(脳血流)は実験全体を通して変化しなかったが、瞑想の内容に応じて局所的に変化した脳血流パターンが観察された。さらに、瞑想時の脳血流活動は、眼窩・背外側前頭前野回、前帯状回、側頭回、ポンズ、小脳など、経営者の注意力ネットワークをサポートしていると考えられている領域で差が見られた(Lou et al 1999)。
99mTc-ECD(エチルシステイン二量体または二等分体)SPECTを使用して、参加者(n = 4)が12週間のアイアンガーヨガトレーニングプログラム(コーエン et al 2009)を受けた小規模な縦断的、介入研究では、灌流も調べられた。各参加者は、プログラム前のベースラインと瞑想のスキャン、プログラム後のベースラインと瞑想のスキャンの両方を受けた。プログラム前とプログラム後のベースラインスキャンにおける平均脳血流比の有意な減少が、扁桃体、背内側前頭前野、前頭前野および後頭前野の各領域で検出された。プログラム前後のパーセンテージ変化の有意差は、内側前頭回、前頭前野(PFC)前頭回、後頭回、下・上前頭回で観察された(Cohen et al 2009)。
99mTc-HMPAO (hexamethylpropylene amine oxime or exametazime) SPECTを実施し、経験豊富な11名の瞑想参加者を対象にクンダリーニヨガの詠唱瞑想中の灌流変化を調べた (Khalsa et al 2009)。この研究では、瞑想中のR脳血流がコントロール条件と比較して、中後頭部、上頭頂部、下側頭回、内側前頭回で減少していることが示された。一方、瞑想時には後帯状回と左上側頭回で活性化が見られた(Khalsa et al 2009)。
最後に、Wang et al 2011)は、ASLを使用して、クンダリーニ瞑想者(n = 10)のベースラインと2つのヨガ/瞑想状態の間の灌流の違いを調査した。この研究では、異なる瞑想状態(「フォーカスベース」の練習と「ブレスベース」の練習)を、瞑想前と瞑想後のベースライン状態と比較した。フォーカスベースの瞑想課題では、内側前頭回と尾状核で脳血流の増加が観察されたが、瞑想前のベースライン対照状態と比較して、下・上後頭回と下頭頂葉で脳血流の減少が検出された。呼吸に基づく瞑想タスク中には、島皮質、扁桃体、海馬、傍海馬、上側頭葉を含むいくつかの皮質および大脳辺縁系の脳構造が、瞑想前のコントロール状態と比較して脳血流の有意な増加を示し、2つの瞑想状態間で異なる脳血流パターンを示唆した。さらに、報告された瞑想の深さと、2回目の瞑想課題の間に、島皮質、下前頭前野、側頭極の脳血流反応の増加との間に強い正の相関関係が観察され、これらは接続性と瞑想の深さの両方の主観的評価と正の関係を持ち、知覚されたストレスと負の関係を持っていた(Wang et al 2011)。
神経活性化に対するヨガの効果
いくつかのfMRI研究では、ヨガをしている人の脳の活性化や変動を調べた(表2Bと図1C)。
4つの研究では、ヨガ瞑想や呼吸法の練習中にfMRIデータを取得した。まず、Engström et al 2010)は、クンダリーニヨガ瞑想の経験が2年未満の8人の被験者を対象とした横断的なfMRI研究で、中等度のヨガ経験者が海馬形成やPFCを含む特定の脳領域の活性化を示すかどうかを、オンオフのデザイン(瞑想と言葉の条件)で調べた。Engström et al 2010)は、被験者がマントラを使って瞑想しているときに、ニュートラルな言葉を発音しているとき(コントロール条件)と比較して、異なる領域で活性化が見られた。瞑想課題中に最も有意な活性化が見られたのは、両側の海馬/海馬傍形成部であった。他にも、中帯状皮質と前頭前皮質で有意なニューロン活性化が見られた(Engström et al 2010)。
第二に、海馬形成における活動の変化は、”OM “チャンティングの血行力学的相関関係を調査したやや非定型的なヨガ研究でも発見された。健康な参加者(n = 12,このうち4人だけが瞑想を含むヨガの正式なトレーニングを受けていた)が「OM」を唱える訓練を受け、この活性化条件の間にスキャンされた(Kalyani et al 2011)。ssss…」の連続生成をコントロール条件とした。プリオリROIは、”OM “詠唱の迷走効果に起因するこれらの領域の期待される不活性化に基づいて、大脳辺縁領域、眼窩前頭前野、前帯状皮質、およびタラミが含まれてた。OM詠唱中に扁桃体、前帯状回、海馬、島皮質、前頭前皮質、海馬傍回、視床で有意な不活性化が観察された(Kalyani et al 2011)。
第三に、Hernández et al 2015)は、サハヤヨガ瞑想中の脳の活性化の増加を調査した。コントロール条件では、瞑想者は呼吸、呼吸動作、心配しないこと、リラックスしていることに意識を集中させ、マインドフルネス瞑想として知られている。ヨガ瞑想中の神経活動パターンをfMRIで探ったところ(n=19)内側前頭回、前帯状回、下前頭回、島状回、上側頭回、内側側頭回において、コントロール状態と比較して神経活動が亢進していることがわかった。コントロール状態と比較した場合の活性化の程度は、瞑想の段階が遅くなり、深くなるにつれて徐々に減少していくようであった。これは、より集中的な活性化によるものではないかと考えられた。さらに、側頭葉の活性化は、コントロールのリラックス状態では音楽がないのとは対照的に、瞑想状態では音楽の存在が原因である可能性があると著者らは報告している(Hernández et al 2015)。
第四に、Hernández et al 2015)の知見と並行して、4人の経験豊富なパタンジャリヨガ実践者において、瞑想相(聴覚固定と視覚固定)中の右前頭前野領域の活性化の増加が観察された。著者らは統計、詳細な人口統計、スキャン取得パラメータについては報告していない(Mishra et al 2017)。
統合失調症スペクトラムと診断された女性集団における安静時のニューロン活動を調べた研究は1件のみで、ヨガ介入(n = 23)と有酸素運動(n = 23)対照のウェイティングリスト(n = 12)群を比較した(Lin er al)。 低周波ゆらぎの振幅(ALFF)によって測定される安静時の局所的な自発的ニューロンゆらぎは、以前に統合失調症患者における変化を示した(Hoptman et al 2010)。ベースラインでの群間の差については報告されていないが、ヨガ群は対照群と有酸素運動群に比べて、クモ膜前野と視覚野のALFFに有意な差を示した。さらに、ヨガの介入は、前帯前野のALFFの有意な減少を誘導した(Lin et al 2017)。
最後に、経験豊富なヨガ実践者における感情喚起タスクまたはワーキングメモリタスク中の神経細胞活動を3つの異なるグループで調査した。より具体的には、Wadden et al 2018)は、感情を喚起する視覚刺激を用いて、レクリエーションアスリート(n = 12,グ全身性エリテマトーデスポーツ)と比較して、ヨガ経験者(n = 19,ヨガスタイル不明)の感情調節への影響を調査した。感情を喚起する段階でのより高い活性化は、ヨガ被験者の上頭頂葉、後心回、前三叉辺上回で認められた。これらの領域は注意喚起意識と自己中心的バイアスの減少と関連している。しかし、レクリエーションアスリートでは、感情の調節時に認知的再評価と関連する領域である下前頭回と側方後頭皮質で感情喚起期に高い活性化を示した(Wadden et al 2018)。
同様に、感情の干渉が認知に及ぼす神経認知的相関関係は、(Froeliger et al 2012a)によって行われたfMRI研究で検討された。この研究では、ヨガ実践者(n = 7)と年齢、性別、教育年数をマッチさせた対照群(n = 7)は、fMRIのセッション中に、否定的または中立的な感情の気晴らしをする前に、イベントに関連した感情の視聴試行とStroop試行を行わせた。観賞試験では、背外側PFCでより大きな活動が観察された。対照的に、Stroop試験では、対照群の方がより大きな活性化反応を示す群間効果により、上前頭前野でより大きな活性化が観察された。Stroop試験の相互作用効果はベントロラテラルPFCで認められた。(Froeliger et al 2012a)は、ヨガ瞑想の実践者は、感情的に有益な情報に反応して解離性前頭幹部依存戦略を選択的に採用する可能性があることを示唆している。
長期的なヨガの練習(瞑想、身体的なポーズ、呼吸法からなる)がワーキングメモリに及ぼす影響を調査した研究は1件だけで、fMRIでシュテルンベルグワーキングメモリ課題を行った(Gothe et al 2018)。本研究には26名の被験者が含まれており、そのうち経験豊富なヨガ実践者13名と年齢と性別をマッチさせた対照者13名が含まれている。両群間で反応時間やパフォーマンスの精度に有意な差は見られなかったが、fMRIの結果では、シュテルンバーグ課題のエンコーディング時にヨガ経験者では背外側PFCの活性化が少ないことが示された。この結果は、経験豊富なヨガ実践者がタスクを実行中に効率性を高めることを示唆している可能性があり、これはワーキングメモリのパフォーマンスに対するヨガのポジティブな影響を示唆する行動学的研究と一致している(Gothe er al)。
機能的接続性に対するヨガの効果
脳の活性化の違いに加えて、fMRIはヨガが脳の機能的接続性に及ぼす潜在的な効果を調べるためにも使われている(表2Cと図1D)。脳内の機能的接続性を調べるために用いられた手法には、ROI-to-ROI分析、シード-to-ボクセル分析、独立成分分析、ネットワークベースの統計学がある。対象となった研究では、以下の安静時ネットワーク(RSN)が調査された。DMN、背側注意ネットワーク(dorsal attention network: DAN)エグゼクティブ・コントロール・ネットワーク(exececutive control network: ECN)サリエンス・ネットワーク、言語ネットワーク、上頭頂部ネットワーク。種子基底分析では、内側前頭前野(mPFC)や前帯状皮質・後帯状皮質(PCC)など、ほとんどの種がDMNに配置されていた。
Froeliger et al 2012a)は、4つのRSN(DMN、DAN、ECN、サリエンスネットワーク)に対するハタヨガ瞑想と瞑想状態の影響をROI-to-ROI分析で調査し、ヨガ瞑想は複数のRSN間のより大きな機能的接続性と関連しているだろうという仮説を立てた。
第一に、ヨガ実践者(n = 7)のDANの領域間相関値が、対照者(n = 7)と比較して高いことが判明した。
第二に、DANのノード内での領域間接続性は、ヨガグループで有意に高かった。ヨガグループ内では、瞑想状態では安静時と比較して、DANとDMNのノード間、また、サリエンスネットワークの前頭前野ノードと複数のDANのノード間で、より大きな機能的接続性が観察された。一方、安静時には、瞑想状態と比較して、DANと背側mPFC(ECNのノード)と右島皮質(サリエンスネットワークのノード)との間に、より大きな機能的接続性が認められた(Froeliger et al 2012a)。
(Hernández et al 2016)は、シード・トゥ・ボクセルfMRI研究において、彼らのVBM研究と同じ被験者を対象に機能的接続性解析を行った。彼らは、精神的静寂の深さの主観的知覚とmPFCの灰白質密度との間に正の相関があることを発見した。この領域はその後、シードからボクセルへの機能的接続性解析のためのシード領域として使用された(Hernández et al 2018)。瞑想状態では、mPFC(シード/ROI)と前部インシュラ/プータメン間の機能的接続性が両側で有意に増加した。対照的に、瞑想状態と安静状態の両方を通して、mPFCと視床/傍海馬回との間の機能的接続性は有意に低下した(Hernández et al 2018)。
同様に、Sevinc et al 2018)もmPFCをシード領域として用いて、縦断的な設定でMBSR(n = 18)介入(ヨガのポーズを含む)とリラクゼーション反応(RR; n = 16)プログラムの共通および解離可能な神経相関を評価した。この研究では、2つの先験的シーズを使用した:mPFC(集中注意に関連する)と前部島皮質(体性感覚の認識における役割のため)。どちらの介入も、安静時と比較して、瞑想中のmPFC(シード)と補助運動野の機能的な結合を明らかにした。しかし、MBSRプログラムは、安静時と比較して、ボディスキャン瞑想中に前島皮質(シード)と前帯状皮質の間の機能的結合が増加するというユニークな関連性を示した(Sevinc et al 2018)。
(Santaella et al 2019)は、健康な高齢女性集団(n = 40;Afonso et al 2017)と同じ被験者)の神経細胞の接続性に対処するために、同じシード(DMNの主な前方シードとしてmPFC)を使用した。彼らは、ヨガグループ(n = 20)のmPFCと角回の間の安静時の接続性が、ペアのヨガ未経験のコントロール(n = 20)と比較して、より大きいことを発見した(Santaella et al 2019)。
合計で2つの研究では、同定されたネットワーク内およびネットワーク間の機能的接続性を調査するために独立成分分析(ICA)アプローチが用いられた。介入的縦断的fMRI研究では、Eyre et al 2016)は、MCI患者におけるヨガトレーニング(n = 14)またはMET(n = 14)の前後で、機能的接続性と記憶テストのパフォーマンスとの相関を調査した。被験者は両群に無作為に割り付けられた。分析は、長期記憶に関する研究で関連性があるとICAによって同定された、DMN、後DMN、言語ネットワーク、上頭頂ネットワークを含む4つのネットワークについて行われた。その結果、言語記憶能力の向上は、DMN内の接続性が高いことと有意に正の相関があることが示された。さらに、機能的接続性の変化と長期的な視覚空間記憶能力の変化との間には、両群とも上頭頂ネットワーク内のクラスターに有意な負の相関が認められた(Eyer et al 2016)。ICAも同様に適用し、VBM解析に加えて、(Dodich et al 2019)は同じ被験者を対象にrsfMRIを行った。その結果、前頭-頭頂部ネットワークの前頭部セクターで有意な相互作用効果(群×時間)が観察された。このネットワーク内では、サハヤヨガのトレーニング後、超低周波ではパワーの減少が、低中周波ではパワーの増加がみられた。前頭-頭頂部ネットワークのパワースペクトルのこの変化は、ウェルビーイングスコアと相関していた(Dodich et al 2019)。
最後に、ヨガと瞑想の違いを切り離す試みとして、(Gard et al 2014)は、ヨガ実践者(n = 16)瞑想実践者(n = 16)対照群(n = 15)の3つの異なるグループにおける機能的接続性を調査した。グループ間で有意に異なる接続のクラスターを検出するNetwork-Based Statistics(NBS)を用いて、馬尾核を中心とした3つのノードから構成される比較ヨガ実践者>対照群の有意差成分が発見された。瞑想者対対照者、ヨガ対瞑想者の比較では有意な結果は得られなかったが、対照者と比較して瞑想者、ヨガ実践者ともに有意な高度の中心性が観察された。さらに、対照群と比較して、瞑想者とヨガ実践者は、多くの脳領域への接続性が等しく強いことが明らかになった(Gard et al 2014)。
脳内代謝物と神経伝達物質に対するヨガの効果
これまでのところ、ヨガ瞑想中の内因性線条体ドーパミン放出を 11C-ラクロプリド PET を用いて調査した PET 研究は 1 件のみであり、経験豊富なヨガニードラ実践者 (n = 8) を対象に、2 日間の休息瞑想とリラクセーション瞑想の間、ランダムな順序で実施した (Kjaer et al 2002)。瞑想中に腹側線条体で有意な8%の結合電位の減少が観察され、行動への準備態勢の低下と関連しており、報酬や快楽効果と最も密接に関連する線条体領域でのドーパミン作動性シグナル伝達の増加を示唆している(Kjaer et al 2002)。
最初のMRS研究では、Streeter et al 2007)は、経験豊富な8人のヨガ参加者からなる1つのグループに、特にヨガの身体的なポーズに焦点を当てたヨガセッションの練習を指示し、対照群(n = 11)はMRSスキャンの前に60分間読書をさせた。MRSボクセルは大脳皮質と深部灰白質構造(正確な位置は特定されていない)に配置された。ヨガ実践者では、ヨガ実践後にGABAレベルの上昇が認められたが、対照群では読書運動後に変化は観察されなかった(Streeter et al 2007)。その後の介入研究では、同じグループが、60分間のヨガ(n=19)またはウォーキング(n=15)介入に無作為に割り付けられた健康な被験者の左視床におけるMRS GABAレベルを測定した(Streeter et al 2010)。ヨガの被験者は、ウォーキング群と比較して、気分と不安の改善が大きかったと報告された。グループ間での視床GABAレベルの有意な変化は見られなかったが、気分尺度(活性化、静けさ、状態-trait不安形質)の変化と視床GABAレベルの変化との間に有意な正の相関がヨガグループで見られた(Streeter et al 2010)。
ある研究では、2型糖尿病患者におけるヨガ介入が右背側前頭葉のN-アセチルアスパラギン酸(NAA)とミオイノシトール(MI)の脳代謝物に及ぼす影響をMRSを用いて調査した(Nagothu et al 2015)。この症例対照研究では、合計68名の患者を、ヨガ群(n=34名)と対照群(n=34名)のいずれかに割り付けた。ヨガグループは、資格を持ったヨガ教師の毎日の監督の下で、週6日、45~60分間、6ヶ月間ヨガ(身体的なポーズと呼吸法を含む)を行った。対照群は特定の運動レジメンを行っていなかった。(Nagothu et al 2015)は、正確な脱落者数も、各グループの女性対男性の正確な年齢と数も報告していない。ヨガ群では対照群と比較して右背側前頭葉のNAA値が高く、MI値が低かったことから、それぞれ神経細胞の完全性が高く、神経グリア機能が低下していることが示唆された。ベースライン値は報告されていない(Nagothu et al 2015)。前述の研究と同じ研究デザインと介入を用いた別のMRS研究では、対照群(n=5)とヨガ群(n=5)の間でNAA値とMI値に差は認められなかった(Santhakumari er al)。
(Yang et al 2016)は、VBM効果に加えて、ヨガ群(n = 14)または記憶力強化トレーニング群(n = 11)のいずれかに無作為に割り付けられたMCI患者25人の被験者を対象にMRSを取得した。MRSの研究は、海馬と背側前帯状皮質にROIを配置して行われた。コリンについては、時間と群間の有意な相互作用効果が検出され、ポストホック分析では、記憶力トレーニング後の海馬でコリンが減少したが、ヨガ群では変化しなかったことが示された。特筆すべきは、ベースライン時のコリンレベルは、記憶トレーニング群の方がヨガ群よりも高かったことである(Yang er al)。
ヨガにおけるこれらの機能的神経イメージング研究に関連する重要な結果の要約は、表2A-Dで提供され、図1Bに示されている(脳血流とグルコース代謝への影響、主に前頭部領域での増加と後部領域での減少を示している。ただし、ヨガの瞑想的内容ごとに特徴的なパターンを示している)図1C(神経細胞の活性化、主にヨガ中の前頭前野での活性化の増加を示し、後野での活性化の減少を示している)図1D(機能的接続性、主にDMN内の前頭前野での機能的接続性の増加を示している)図1E(神経伝達物質と代謝物、ヨガの介入と測定された代謝物に応じて広範な効果を示している)が提供されている。
考察
ヨガの神経画像効果に関する発表された研究は、ヨガの様々な形態、ほとんどが少人数のグループと異質なターゲットの評価によって特徴づけられているが、比較的一貫した脳の構造的・機能的変化が見出されている。これらは、臨床研究研究の増加するボディで報告されているヨガの有益な身体的・心理的効果と推定されるものと関連している可能性がある(Field, 2011, 2016; Vancampfort et al 2012; Villemure et al 2015)。
全体的に、形態学的な神経画像所見は、対照群と比較して、ヨガ実践者の地域灰白質密度またはボリュームの増加と一致している。これらの研究では島皮質はROIとして用いられていなかったが、島皮質の灰白質密度の増加は、全脳解析で最も一貫して報告されている(Froeliger et al 2012b; Villemure et al 2015; Hernández et al 2016; 図1A)。これらの結果は、瞑想実践者の形態学的神経イメージングと一致しており、島皮質の増加が瞑想スタイルの違いによって最も一貫した構造的変化であることがわかった(Fox et al 2014)。
島皮質における形態学的効果に加えて、本レビューに含まれる複数の機能研究(Kalyani et al 2011;Wang et al 2011;Froeliger et al 2012a;Hernández et al 2015,2018)では、ヨガ実践者における島皮質の活性化または接続性の違いが示された(図1C,D)。重要なことに、これらの多くはヨガの完全な三位一体を調査しておらず、身体的なポーズ(アーサナ)をとらずに瞑想に焦点を当てたヨガスタイルのみが含まれてた。並行して、マインドフルネスに基づく介入を含む瞑想の機能的効果に関するメタアナリシスと系統的レビューでは、複数の瞑想スタイルにわたって一貫して島皮質が採用されていることが報告されており(Fox et al 2016;Young et al 2018)瞑想プロセス全体にわたって島皮質の重要な役割が示唆されている。
島は、身体の相互受容的な意識、自分自身の身体の心理的状態に対する感受性(例:心拍数の増加、汗をかいた皮膚、胃の緊張などを感じる)共感、そしておそらくはメタ認知において中心的な役割を果たしている(Gu et al 2012; Simmons et al 2013)。島皮質は瞑想中だけでなく、姿勢の変化やゆっくりとした呼吸時にも関与している(Critchley et al 2015)。様々な神経イメージング研究では、間受的意識が高いほど効果的なストレス対処能力の向上と関連しているのに対し、間受的意識が低いことがうつ病患者で観察されていることが示されている(Avery et al 2014)。ヨガは、島皮質と感情処理や感覚処理の調節に関与する脳領域(PFCなど)との間の機能的接続性の増加を誘発する可能性があり、島皮質の灰白質密度が高くなる可能性がある。
さらに、海馬における灰白質体積または密度の増加は、フルトライアドを含むヨガ研究を含む、さまざまな形態学的研究(Froeliger et al 2012b; Hariprasad et al 2013; Villemure et al 2015; Gothe et al 2018)において一貫して観察された。これらの結果に沿って、瞑想のいくつかの機能的・構造的神経イメージング研究でも、海馬の形成に違いがあることが報告されている(Fox et al 2014,2016)。これらの効果は、ヨガ介入後に観察されたように、神経可塑性のマーカーであるBDNFレベルの上昇と関連している可能性がある(Naveen et al 2016)。BDNFは海馬領域内で高濃度に存在することから、海馬で観察される形態学的効果に役割を果たしている可能性がある(Murer et al 2001)。対照的に、ヨガや瞑想の実践者を対象とした大規模な集団研究(両者を区別せずに)では、対照群と比較して海馬体積の減少が横断的に観察されており、これは初期の生活のストレス要因に起因している可能性がある。瞑想やヨガの練習は海馬体積と経時的に有意な関係はなかった(Gotink er al)。
扁桃体は2つの形態学的構造研究で明示的に調査され、密度と体積の減少が観察された(Hölzel et al 2009; Gotink et al 2018)。知覚ストレススコアの変化と扁桃体の体積の間には有意な相関が見られた(Hölzel et al 2009)。実際、広範な証拠は、扁桃体と海馬の両方の構造と機能のストレス誘発性変調の神経的相関を示した。したがって、扁桃体が小さくなり、海馬の体積が増加したのは、ヨガの練習を通じて経験されたストレスが少ないためであると考えるのが妥当であろう(Roozendaal et al 2009; McEwen et al 2016; McEwen, 2017; Gotink et al 2018)。
感情的なタスクや認知機能と比較して、ヨガ瞑想中の神経活性化の違いを調査した機能的神経イメージング研究では、様々な皮質および皮質下の脳領域でヨガによって誘発される効果が発見されている。しかし、すべての研究で不均一な結果が得られているが、これは研究デザインの不均一な性質や、機能活動画像中に複数のヨガのスタイルやタスクがあったことに起因していると考えられる。
ヨガ瞑想の練習中の脳血流/灌流およびグルコース代謝の変化に焦点を当てた5つの研究から、脳血流はヨガ瞑想によって影響を受けることが示された(Herzog et al 1991; Lou et al 1999; Cohen et al 2009; Khalsa et al 2009; Wang et al 2011)が、脳血流への影響は瞑想の種類および焦点に大きく依存する可能性がある(Fox et al 2016; 図1B)。8つの研究がヨガ瞑想中の神経活性化の変化を調査しており、大多数の研究ではヨガ瞑想中に前頭前野の活性化が増加していることが示されている(Froeliger et al 2012c; Hernández et al 2015; Lin et al 2017; Mishra et al 2017)。
機能的接続性に関する研究(Froeliger et al 2012a; Gard et al 2015; Eyre et al 2016; Hernández et al 2018; Sevinc et al 2018; Dodich et al 2019; Santaella et al 2019)では、複数のネットワークおよび領域において、対照と比較して、ヨガの被験者において改善された接続性が観察された。機能的接続性に関する研究の半数はDMNに焦点を当てており、PFCのシードを用いたシードベースの分析を用いていた(Hernández et al 2018;Sevinc et al 2018;Santaella et al 2019)。
異なるヨガスタイルを調査した研究では、異質な結果が見られたが、PFCに向かって一貫した接続性の増加が見られた。一般的に、DMN内の神経活動はマインドワンダリングと相関することが知られており、それと引き換えに幸福度の低レベルと関連している。マインドワンダリングを減らす方法としては、ヨガの瞑想が考えられる。瞑想は以前、DMNと自己モニタリングや認知制御に関与する領域との間のより強い結合に関連していることが知られている(Brewer et al 2011)。
ドーパミンD2/3受容体放射性物質11C-ラクロプリドを用いて、ドーパミン機能に対するヨガ(ヨガニドラ瞑想)の効果を調査した研究は1件のみで、ヨガニードラ瞑想の練習中に腹側線条体でドーパミン放出が有意に増加したことを示している(Kjaer et al 2002)。
腹側線条体は、目標指向行動、行動感応、情動状態の変化の基礎となる回路に重要な役割を果たしている (Ito et al 2004) が、報酬/モチベーション回路にも重要な役割を果たしている (Volkow et al 2012)。また、大脳基底核を介してドーパミン系が皮質下前頭前野相互作用の調節に関与していることにも注目すべきである。合計5件の研究では、脳内代謝物濃度の変化を調べるためにMR分光法を使用した。ほぼすべての研究で異なるROIが用いられた。
ストレスの軽減、MIの減少、NAAレベルの上昇と関連したGABAレベルの上昇がヨガ後に報告されている(Streeter et al 2007,2010;Nagothu et al 2015)。GABAレベルの増加は、複数のヨガ研究で観察されるように、ヨガ中またはヨガ後のPFCの活性化の結果である可能性がある(Froeliger et al 2012c; Hernández et al 2015; Lin et al 2017; Mishra et al 2017)これはグルタミン酸を増加させ、視床の網状核を刺激してGABAの分泌を増加させると考えられている(Guglietti et al 2013)。
ヨガの中心的な効果の基礎となるメカニズムを構成する神経生物学的モデルは、ヨガは脳血流、大脳活動、グルコース代謝の増加を誘導し、皮質基質の追加的な高需要の募集を反映しているが、減少は、学習が進むにつれて、より少ないニューロン資源を使ってヨガの練習を行うことができることを示唆している(Dayan and Cohen, 2011)。機能的な活性化を繰り返すことで、樹状突起形成/シナプス形成、神経新生(海馬に限られる)ミエリンのリモデリング、および繊維の再編成を介して、白色および灰白質に影響を与える形態学的変化を誘発する可能性がある(Dayan and Cohen, 2011; Zatorre et al 2012; Lövdén et al 2013)。
その代償として、機能的連結性の変化が起こり得る。さらに、複数のボトムアップ(例えば、制御された呼吸や物理的なポーズ)とトップダウン(例えば、集中した注意やインターセプト)のプロセスも、ヨガのようなマインドボディ介入において重要な役割を果たしている(Muehsam et al 2017)。ヨガの練習の有益な効果は、その3つの主要な構成要素である深呼吸、瞑想、身体的なポーズの組み合わせが副交感神経系を活性化し、それによって脳内の主要な抑制性神経伝達物質であるGABA濃度を増加させることであり、(Streeter et al 2007, 2010)の知見と一致している。
副交感神経の緊張の増強効果は、主に深くゆっくりとした呼吸によって誘発される圧反射反応によって引き起こされる迷走神経求心性を介して、また、ヨガセッション中の様々なヨガのポーズ(チンロック、逆転、開胸のポーズ)によって活性化された圧受容体によって起こる可能性がある(Streeter er al)。 これはまた、ヨガの不安やストレス緩和効果の基礎となるかもしれない。ストレスに関連したコルチコステロイドの放出を打ち消すことはGABAを減少させ、長期的な結果をもたらす可能性のある神経学的形態学的変化を引き起こす可能性がある(Roozendaal et al 2009; McEwen et al 2016)。
このレビューでは、ヨガの神経生物学的効果を研究するための神経画像法(MRI、PET、SPECTを含む)の応用をまとめようと試みた。それにもかかわらず、一貫してデザインされた研究の数が比較的少なく、不均質であったため、結果の正式なメタアナリシスは適切ではないと考えられた。これらの比較的小さなサンプルサイズは、効果を特定する感度を制限している可能性がある。さらに、38%の研究(13/34件)では対照群が含まれていなかった。対照群と対照条件の選択は非常に異質であり、認知タスクから運動タスクまで様々であるため、研究の比較が困難である。
対照群や対照条件としては、それぞれ、身体運動、または身体運動レベルに基づくマッチングが一般的に最も一貫して用いられていた。これは、ヨガ研究における比較群に関するシステマティックレビューの結果と一致している(Park er al)。 身体運動の研究は、ヨガの効果が主に運動によって決定されるのか、ヨガが付加的な効果を提供するのかを確立することを可能にする。集団参加、注意、活動の非特異的効果をコントロールするために重要であることを考えると(Park et al 2014)、しかし、活動的な対照群を用いた研究の希少性を考えると、私たちは将来の研究デザインに組み込むことを強く提唱する。
身体活動が脳の構造と機能に及ぼす影響に関する研究では、いくつかの類似点が見出されている。一般的に、身体活動(または有酸素トレーニング)もまた、灰白質密度の増加と関連している。最も一貫して観察された領域には、海馬、大脳皮質、小脳、運動野などの運動関連領域が含まれていた(レビューは、Thomas et al 2012年およびErickson et al 2014年を参照のこと)。また、Sextonらのレビューでは、身体活動と脳白質構造との関連が複数の研究で示唆されていることが示されている(Sexton et al 2016)。
機能的活動の変化については、Herold et al 2020)は、運動に反応して特に前頭葉、小脳、海馬に深遠な変化があることを報告している。したがって、特にヨガの実践に関連しうる認知的、実行的、運動的効果については、今後の研究デザインでは、構造的、機能的効果が純粋な身体運動の要素とは異なるかどうかを明確に実証しようとすべきである。なぜなら、運動は運動強度、運動時間、有酸素運動と無酸素運動の違い、局所的または包括的な運動レベルの違いなど、多くの変数によっても変化するからである。
同様に、ヨガの介入のスタイルや強度にもばらつきがあり、これらの知見に明確な共通点を見つけることは困難である。さらに、本レビューに含まれるヨガ関連の神経画像学的研究のうち、半数は瞑想を中心としたヨガスタイルの効果を調査しているのに対し、残りの半数は身体的なポーズ(アーサナ)を含むヨガを調査している。後者は、現在の欧米では身体的なポーズを中心としたヨガが主流となっているが、後者は身体的なポーズを中心としたヨガである。ヨガのスタイルによって、それぞれのヨガのサブコンポーネントが異なるため、脳への影響が異なる可能性がある。
Villemure et al 2015 の研究では、一連のヨガの実践後に観察される構造的な変化には、ポーズ、呼吸法、瞑想が異なる形で貢献していることが示されている(Villemure et al 2015)。研究対象者の約4分の3は女性であったが、これは生涯ヨガを実践している人は女性である可能性が高いという観察結果と一致している(Cramer et al 2016)。ほとんどの研究では無作為化は行われなかった。4つの研究は単なる探索的デザインであり、多重比較をコントロールしていなかった(Lou et al 1999; Cohen et al 2009; Hariprasad et al 2013; Hernández et al 2016)ため、偽陽性の知見が得られる可能性が高くなっていた。
今後の研究は、より大規模でより定義された集団において、ヨガの各サブコンポーネントが特定のニューロンパターンに及ぼす影響と、これらのサブコンポーネント間の相互作用を分離することを目的としているはずである。最後に、否定的な所見が報告されないままになっている可能性があり、出版物の偏りという一般的な問題も存在すると思われる(Egger et al 1997)。
結論
比較的少ないものの、主に健康な被験者を対象としたヨガの実践に関する神経画像の証拠が拡大していることに基づいて、ヨガがインターセプト、姿勢、モチベーション、高次の執行機能に関与する脳領域に構造的、機能的な効果をもたらすことが示されている。全体的に見て、最も一貫した構造的効果は海馬と島皮質で観察され、機能的研究では主に前頭幹部と注意領域の増加が示されている。しかし、研究の数はまだ限られており、不均質であり、含まれている異なるヨガスタイルの間での不均質性と、応用研究プロトコルの大きなばらつきのために、いくつかの矛盾が存在する。
ヨガの潜在的な有益な効果をさらに調査するために、健康な脳だけでなく、例えば大うつ病、PTSD、不安状態などの気分障害や不安障害のような疾患状態においても、ヨガの潜在的な有益な効果を調査するために、より広範で、よくデザインされた、マルチモーダル/マルチパラメトリックな研究が行われるべきであると考えられる。神経イメージングと神経生理学的技術(脳波、筋電図など)を統合することで、イメージング結果を神経生理学的、行動学的な評価や幸福度の向上と結びつけることが可能になる。