Contents
The impact of frequent napping and nap practice on sleep-dependent memory in humans
要旨
昼寝は長期的な記憶形成に効果があり、多くの人が日中の機能を向上させるために使用している。このような利点があるにもかかわらず、米国では約47%の人が昼寝を避けている。本研究の目的は、少なくとも週に1回の昼寝を推奨する人(nap+)が、昼寝をほとんどしない、または全くしない人(nap-)と比較して、昼寝に依存した記憶の定着を含む昼寝の成果に違いがあるかどうかを明らかにすることであった。さらに、4週間の昼寝の練習や制限が睡眠とパフォーマンスのプロファイルを変化させるかどうかを検証した。
知覚学習タスクを用いて、昼寝は昼寝をしない人(ベースライン時)に比べて昼寝+の人の方がパフォーマンスを向上させることがわかった。さらに、パフォーマンスの変化は、各グループで異なる電気生理学的睡眠機能と関連していた。昼寝+群では、紡錘体密度はパフォーマンスの向上と正の相関があり、これは訓練された視覚野とは反対側の半球の紡錘体に特有の効果であった。nap-群では、低速振動パワー(0.5-1Hz)がパフォーマンスと相関していた。驚くべきことに、昼寝の練習や制限を4週間行っても、パフォーマンスや睡眠中の脳活動に変化は見られなかった。
これらの結果は、個人差がパフォーマンスやより良い昼寝者(better napper)になるための能力に昼寝の潜在的な利点に影響を与える可能性を示唆している。
はじめに
睡眠は、新たに学習した情報の記憶の安定化や強化(すなわち、記憶の統合)に重要な役割を果たしている1。昼間の昼寝は、これらの記憶プロセスを促進する上で、夜行性睡眠と同等の効果を発揮することがある2-4。昼寝によるパフォーマンス向上効果は、エピソード記憶5,6,感情調節7,8,手続き能力9,注意力10など、認知能力の広い範囲で認められている。また、昼寝は創造性11,12や生産性13を高め、アスリートのパフォーマンスを向上させ14, シフトワークによる疲労に対処するのに役立つとしても支持されている15-17。
昼寝の利点が実証されているにもかかわらず、誰もが昼寝をしているわけではない。米国では、全米睡眠財団の睡眠健康指数201418によると、成人の53%が定期的に昼寝をしており、過去7日間に少なくとも1回は昼寝をしたことがあると定義されている。「お昼寝をする人」または「お昼寝をしない人」の定義についてはコンセンサスがないが、若く健康な集団におけるお昼寝行動は、頻繁にお昼寝をすると報告されている他の集団(乳幼児、就学前の子ども、高齢者、睡眠障害者など)とは異なる制御パターンと環境機会に従っている可能性が高いと考えられる19-24。
若く健康な成人では、主観的な報告と客観的な測定値(例:睡眠脳波)に一貫したパターンが見られ、昼寝を推奨する人とそうでない人の間には実際に違いがある可能性があることを示唆している25。昼寝をしない人は、目が覚めたときにグロッギーで非生産的な感じがして、昼寝から何の恩恵も受けられないので、昼寝を避けていることが多いと報告している26。生理学的レベルでは、この起床後の認知障害(すなわち、睡眠慣性)は、徐波睡眠(徐波睡眠)の量と徐波睡眠からの覚醒に関連している可能性がある27。
実際に、我々は以前に、非昼寝は深い徐波睡眠でより多くの時間を費やすことを発見した一方で、頻繁な昼寝は、日中の昼寝中に光、ステージ1とステージ2の睡眠でより多くの時間と関連していた28。このように、昼寝の日常的な経験は、昼間の睡眠の質を変化させる可能性があり、その結果、昼寝中の記憶処理や覚醒時の睡眠慣性の症状に影響を与える可能性がある。
ここでは、昼寝に依存した記憶の定着と睡眠時の慣性症状に対する事前の昼寝(昼寝+ vs. 昼寝)の影響を調査した。さらに、クロスオーバーデザインを用いて、4週間の「昼寝練習」(週3回以上の昼寝)または「昼寝制限」が、昼寝+と昼寝-の人の睡眠生理学とパフォーマンスプロファイルに変化を与えるかどうかを検証した。参加者は、研究期間中の3つの異なる時点(ベースライン、中間介入、介入後)で、ラボ内での睡眠ポリグラフ記録(PSG)による昼寝を行った。それぞれのラボ内昼寝の日に、昼寝前と昼寝後の行動パフォーマンスを知覚学習タスク(テクスチャ弁別)を使用して測定した。我々はまた、降順減算テスト29,30を使用して昼寝後の認知機能をテストし、一日中主観的な眠気の評価を収集した。
我々はまず、昼寝をした人と起きたままの人を比較することで、テクスチャ識別タスクの当日のパフォーマンスのみが昼寝後の改善を示すという知見2を再現することを目的とした。重要なことは、昼寝グループは昼寝+と昼寝-の両方の人で構成されていたということである。仮説として、昼寝経験の違いを考慮に入れることで、昼寝後の昼寝+と昼寝-の間には、より大きな知覚学習の向上、より少ない睡眠慣性(降順減算テストのパフォーマンスによって指標化される)昼寝+の人のより少ない主観的な眠気など、異なるパフォーマンスの結果が見られると仮定した4。要約すると、我々は、昼寝+の参加者は昼寝+の参加者よりも昼寝後の方が良い仕事をし、良い気分になるという仮説を立てた。
第二の目的は、睡眠脳波の特徴の個人差がどのようにパフォーマンスの結果と関連しているか、すなわち、各睡眠段階の分数と割合、睡眠スピンドル、遅発性振動(0.5-1Hz)非遅発性眼球運動(NREM)睡眠時のデルタ(1-4Hz)とシグマ(12-15Hz)のパワー、および速発性眼球運動(REM)睡眠時のシータ(4-8Hz)のパワーを調べたことであった。これまでの研究では、必ずしも相互に排他的ではないが、複数の統合プロセスが知覚学習の根底にある可能性が示唆されている。これらの提案されている各プロセスは、異なる睡眠の特徴、例えば、シナプスの恒常性と遅波活動31,知覚劣化と徐波睡眠2からの回復、NREM睡眠中の再活性化32と睡眠紡錘体活動33(すなわち、アクティブシステム統合仮説)1,トレーニング中に達成されたパフォーマンスレベル以上の改善につながるシナプス強化、これはNREM睡眠とREM睡眠の組み合わせに依存している可能性がある2,34-37などと関連している傾向がある。この図の全体的な複雑さを考えると、グループレベルの違い(例えば、昼寝中の徐波睡眠の増加)は、あるグループが昼寝からより多くの知覚学習の恩恵を受けるかもしれない理由を十分に説明できるとは考えなかった。むしろ、昼寝の経験が睡眠の特徴とパフォーマンスの結果との関連を緩和するのではないかという仮説を立てた。具体的には、昼寝+の人は昼寝に関連したパフォーマンスの改善をより強く示すと考えられたため、タスクの改善に関連した睡眠脳波の特徴(スピンドルとレム睡眠)が、昼寝+の人のパフォーマンスの変化と昼寝+の人のパフォーマンスの変化とより強く関連していると予想した。
最後に、昼寝に関連した学習効果が経験に依存しているかどうかを検証することを目的とした。我々は、昼寝は訓練可能なスキルであり、昼寝を4週間練習した昼寝者は、昼寝後の知覚学習の向上、睡眠慣性の減少、主観的な眠気の減少を示すだろうという仮説を立てた。
結果
訪問1:昼寝前の昼寝は昼寝後のパフォーマンス、睡眠脳活動、睡眠慣性に影響を与えるか?
知覚学習。昼寝 vs. 起床
参加者は、同じ日に2回、朝と夕方に1回のテクスチャ識別タスク(TDT)を行い、タスクセッションの間は覚醒したまま、または昼寝をとってた。セッション間の閾値の差を計算し、昼寝は起床時と比較してTDTのパフォーマンスを向上させるという古典的な知見2を再現した [t(67) = 1.95, p = 0.056, Cohen’s d = 0.49, 独立標本のt-検定]。昼寝をした参加者では識別閾値が有意に改善し[t(47) = 2.69, p = 0.01, 1標本t-test]、覚醒したままの参加者ではゼロと差はなかった[t(20) = -0.58, p = 0.57, 1標本t-test](図1b)。パフォーマンスの変化は、徐波睡眠分(r = 0.36,p = 0.01)と徐波睡眠%(r = 0.32,p = 0.03)および徐波睡眠とREM分の積(徐波睡眠xREM、r = 0.34,p = 0.02)と正の相関があった。後者の結果は、我々がここで再現を目指した古典的な研究2,36 と一致しているが、これは 徐波睡眠とREMのパフォーマンス向上への貢献度を統計的に検定するための最良の方法ではないかもしれないことに注意してほしい37。また、学習はステージ2%と負の相関があった(r = -0.33, p = 0.02)が、これはステージ2と徐波睡眠の間のトレードオフを反映している可能性がある(ステージ2と徐波睡眠の量は逆相関していた;分:r = -0.45, p = 0.001, パーセント:r = -0.69, p < 0.001)。
図1
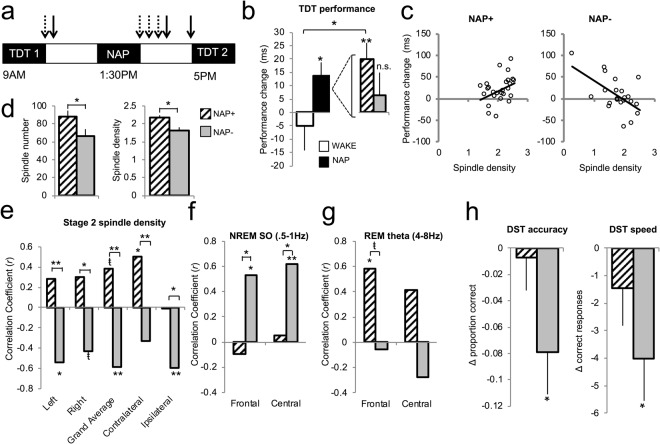
訪問1 ベースライン。(a) インラボ試験日の手順。テクスチャ弁別課題(TDT)の閾値は午前9時と午後5時に得られた。すべての参加者は1:30-3:30PMの間に昼寝した。実線矢印はカロリンスカ眠気尺度(KSS)の投与時間を示し、破線矢印は下降減算テスト(DST)の投与時間を示す。b)Wake(n = 21,白棒)およびNap(n = 48,黒棒)群のベースライン時のTDT閾値の差。Nap群内では、Nap+(n = 26,ハッチングバー)のみが学習を示した;Nap-(n = 22,グレーの実線バー)は有意な改善を示さなかった。後続のすべてのパネルは、特に断りのない限り、各グループで表される独立したデータポイントの数(n)が同じである。(c) パフォーマンスの向上は、ステージ2の紡錘密度(プロットされた大平均)と相関していたが、nap+(r = 0.38,p = 0.055)とnap-(r = -0.59,p = 0.004)では反対の方向にあった。(d) Nap+は、Nap-よりもステージ2の間、より多くのスピンドル(p = 0.04)およびより大きなスピンドル密度(p = 0.008)を有していた(示されたC3-P3avg)。(e) パフォーマンスの向上とステージ2の睡眠紡錘密度の間の相関係数(ピアソンr)(注:Nap-群では、左の相関はn = 20であり、対照側とイプシラテラルの相関は両方ともn = 21であり、不良電極からの欠落によるものである)。また、パフォーマンスと(f)NREM緩徐振動パワー(SO,0.5-1Hz)と(g)REMシータパワー(4-8Hz)の間には、nap+/nap-グループ分けに基づいて異なる関連があった。パネル(e)(f)および(g)については、バーの上または下にアスタリスクは、有意な相関を示し、括弧は、グループ間のr値の有意差を示す(注:nap-におけるNREM SO相関はn = 21を持っていた;レムシータ相関はnap+前頭n = 15,nap+中央n = 17,nap-前頭n = 16,nap-中央n = 15を持っていた;減少nは、レム睡眠および/または悪い電極を含まないnapによるもの)。h)昼寝から目覚めて5分後のDSTパフォーマンスの変化(注:昼寝+ n = 25および昼寝- n = 20; 実験者のエラーおよび包含基準を満たしていないパフォーマンスのために減少したn)。ŧ'”はp≦0.07を示し、*はp<0.05を示し、**はp<0.005を示す。エラーバーは±1 SEM。
知覚学習。昼寝+ vs. 昼寝
我々は上記のデータを再解析し、昼寝グループを昼寝前の経験に基づいて2つのサブグループに分離した(昼寝+と昼寝-)。予測通り、昼寝+群では昼寝後に有意なパフォーマンスの向上が見られたのに対し、昼寝-群ではパフォーマンスの変化はゼロから差がなかったのに対し、昼寝+群では有意なパフォーマンスの向上が見られた[t(25)=3.38,p=0.002,1サンプルt検定](図1b)両群間の差は統計的に有意ではなかったが[t(46)=1.35,p=0.18,Cohenのd=0.39,独立サンプルt検定)。起床群と比較して、昼寝+群のみが有意に良好であった[昼寝+ vs. 起床:t(45)=2.42,p = 0.02,Cohen’s d = 0.70,独立標本t-test;昼寝- vs. 起床:t(41)=0.93,p = 0.36,Cohen’s d = 0.28,独立標本t-test]。これらの効果は、覚醒、昼寝+および昼寝-グループ間の昼寝前閾値の違いによるものではなかった[F(2,66) = 0.79,p = 0.46,一方向ANOVA]。上記の結果と合わせて、この分析は、昼寝対覚醒の分析で観察された昼寝依存性の改善は、昼寝+の個人によって駆動されたことを示しており、すべての個人が昼間の睡眠から学習の利益を示すわけではないことを示唆している。
昼間の睡眠アーキテクチャ
最初のラボ内訪問時(介入群割り付け前)に、NAP+とNAP-の間で、総睡眠時間、分数、またはどの睡眠段階の割合にも有意差は認められなかった(すべてps > 0.29)(補足表2)。日中の睡眠が夜間の睡眠に直接関係している可能性があるため、アクチグラフィを用いて、前夜の睡眠をnap+とnap-の間で比較したところ、有意差は認められなかった(補足表3)。さらに、実験日前夜の総睡眠時間は、どちらの群でも昼寝睡眠段階とは相関しなかった(すべてのps > 0.17)またはサンプル全体では相関しなかった(すべてのps > 0.14)。
しかし、昼寝中の睡眠紡錘イベントには違いが認められた。Nap+は、Nap-と比較して、ステージ2の睡眠中に約31%多くの睡眠紡錘イベントを持っていた[C3-P3avg、Nap+平均87.6(SD = 34.5)対Nap-平均66.8(SD = 33.7)紡錘イベント、t(46) = 2.11,p = 0.04,コーエンのd = 0.61,独立標本のt検定]。図1d)。睡眠ステージ2と徐波睡眠の持続時間に被験者間のばらつきがあることを考慮して、残りの解析は紡錘体密度(紡錘体数/分)に焦点を当てて行った。左半球では、ステージ2の睡眠では、nap+の方がnap-よりも高い紡錘体密度を示した[C3-P3avg、nap+の平均2.2(SD = 0.4)対nap-の平均1.8(SD = 0.5)紡錘体数/分、t(46)=2.79,p = 0.008,コーエンのd = 0.80,独立標本のt検定]が、右半球では差は認められなかった(C4-P4avg、p = 0.41)。また、どちらの半球でも徐波睡眠時には睡眠紡錘の違いは確認されなかった(すべてのps>0.10)。
次に、前頭部(F3-F4avg)と中央部(C3-C4avg)の電極部位にわたって、関心のある特定の周波数帯(NREM SO、デルタ、およびシグマ、REMシータ)のスペクトルパワーを調べた。NREM睡眠中のどの周波数帯域においても、nap+とnap-の間に差はなかったが、REM睡眠では、nap+の方が中心部位よりも大きなシータパワー[t(30)=2.15,p=0.04,コーエンのd=0.77,独立標本のt検定]を有していた。これらの結果を合わせると、昼寝睡眠生理学は、左半球のスピンドルとレムのシータパワーを除いて、ほとんどの睡眠変数にわたって、昼寝+群と昼寝-群で同等であったことを示している。では、睡眠の特徴と行動の関係が、これらのグループでどのように異なるかを詳しく見ていく。
脳と行動の関係。昼寝+ vs. 昼寝
次に、昼寝+/-の参加者について、睡眠の特徴とパフォーマンスの関連性を個別に調べ、昼寝頻度群がこれらの関連性に及ぼす中和効果を検証した。事前の昼寝経験は睡眠段階とパフォーマンスの変化の関連を中和しておらず、昼寝+と昼寝-の両方で相関係数の方向性と大きさが類似していた。しかし、睡眠紡錘がパフォーマンスとどのように関連しているかについては、両群間でかなりの違いがあった(図1c,e)。nap-では、ステージ2(r = -0.59,p = 0.004)徐波睡眠(r = -0.37,p = 0.13,負の相関があるが有意ではない)およびNREMステージを合わせた(r = -0.62,p = 0.002)の間、パフォーマンスはスピンドル密度(C3-C4-P3-P4avg)と一貫して負の相関があった。nap+では、ステージ2のパフォーマンスとスピンドル密度の関係は正の傾向があり(r = 0.38,p = 0.055)徐波睡眠またはNREMを組み合わせたスピンドルでは有意な関係はなかった(徐波睡眠:r = -0.11,NREM:r = 0.16,両方のps > 0.45)。両群の相関係数の統計的な差は、ステージ2の睡眠スピンドル(z = 3.47,p = 0.0005)とNREM併用(z = 2.84,p = 0.004)で有意であり、昼寝群がスピンドルとパフォーマンス変化の関係を緩和していることが示された。
TDTでの学習が網膜特異的であることを考慮して38,我々はさらに、訓練された視野の位置に相対的に同側半球と対側半球のステージ2のスピンドルを調べた。Nap+では、網膜学習効果によって予測される結果のパターンが示され、パフォーマンスと対側の紡錘体との間に正の相関があり(r = 0.50,p = 0.009)パフォーマンスと同側の紡錘体との間には有意な関係はなかった(r = -0.008,p = 0.97)。nap-では、対側(r = -0.33,p = 0.14)と同側(r = -0.60,p = 0.004)の両方のスピンドルがパフォーマンスと負の相関を示したが、この関連は同側のスピンドルについてのみ有意であった。対側(z = 2.84,p = 0.004)と同側(z = 2.16,p = 0.03)の両方の効果について有意な中和があった。
興味深いことに、NREM SOとデルタパワーは、強く昼寝のパフォーマンスの変化と正の方向に相関していた(SO正面:r = 0.53,p = 0.013; SO中央:r = 0.62,p = 0.003; デルタ正面:r = 0.52,p = 0.015。 52,p = 0.015;デルタ中央:r = 0.60,p = 0.005)ではなく、nap+(SO前頭:r = -0.10,p = 0.63;SO中央:r = 0.06,p = 0.79;デルタ前頭:r = 0.01,p = 0.95;デルタ中央:r = 0.21,p = 0.30)であった(図1f)。我々は、nap-グループ内の強い相関が、徐波睡眠の間のSOおよびデルタパワーによって潜在的に駆動されたことに注意し、それは一貫して、ステージ2と比較してより大きなマグニチュードの相関係数を示した(SO正面:ステージ2 r = 0.02 vs. 徐波睡眠 r = 0.38; SO中央:ステージ2 r = 0.12 vs. 徐波睡眠 r = 0.38。ステージ2 r = 0.12 vs. 徐波睡眠 r = 0.45; デルタ正面:ステージ2 r = -0.05 vs. 徐波睡眠 r = 0.22; デルタ中央。ステージ2 r = 0.11 vs. 徐波睡眠 r = 0.47)。) 相関係数は、正面(z = 2.20,p = 0.03)と中央(z = 2.11,p = 0.03)の部位に対するNREM SOパワーについては群間で有意に異なった;NREMデルタパワーの相関は有意には異なっていなかった(正面:p = 0.07,中央:p = 0.10)。NREMのシグマパワーは、nap-(前頭:r = -0.45,p = 0.03;中央:r = -0.38,p = 0.08,有意ではない)でのみパフォーマンスと負の相関を示し、nap+(前頭:r = -0.01,p = 0.96;中央:r = 0.15,p = 0.46)では有意ではなかった。しかし、中庸化の検定は有意ではなかった(前頭:p = 0.10;中央:p = 0.08)。レムシータパワーは、NAP+(前頭:r = 0.58,p = 0.02;中央:r = 0.41,p = 0.10)ではパフォーマンスと正の相関があったが、NAP-(前頭:r = -0.05,p = 0.84;中央:r = -0.28,p = 0.32)では有意ではなく(図1g)中庸化の検定は有意ではなかった(いずれもp = 0.07)。紡錘体の結果とは異なり、どの周波数帯においても網膜頭特異的な違いは見られなかった。これは、睡眠中に発生する大域的な振動に比べて紡錘体がより局在的な性質を持っていることに関係しているのかもしれない39-42。以上の結果から、昼間の睡眠中の圧密化メカニズムに関連した振動回路の違いが明らかになった。具体的には、事前の昼寝頻度が、対側のステージ2スピンドルと学習、およびNREM SOパワーと学習の間の脳行動関係を有意に修飾することが明らかになった。
主観的な眠気と睡眠慣性:NAP+とNAP-の比較
非居眠りをしない人は昼寝を楽しんでいないと報告することが多く26 、昼寝後の気分の悪さや生産性の低さを訴えるという逸話があるため、我々は昼寝後の主観的な眠気と認知機能を測定した。参加者は、研究日中に3つの時点で主観的な眠気を評価した。1)昼寝前、2)昼寝後10分、3)昼寝後90分である(図1a参照)。全体的に、主観的な眠気は一日を通して減少した[F(2,82)=16.46, p < 0.001, 部分 eta2 = 0.29, 混合ANOVA 主効果]。 06,ペアードt検定)および起床後約90分後の眠気のさらなる減少は、昼寝後10分と比較して有意であった(t(44)=4.81,p < 0.001,ペアードt検定)。群による主効果はなく(p = 0.36,混合ANOVA)タイムポイント×群間の相互作用はなく(p = 0.88,混合ANOVA)両群ともに昼寝による覚醒度の向上は同様であることが示された。
我々は、参加者が経験した睡眠の慣性の程度を降順減算テスト(DST)29,30を用いて、午前11時と昼寝から目覚めてから5,20,35分後の認知機能を測定した(図1a参照)。午前11時(昼寝前)には、昼寝+/-の間に速度(正解数)や正確さ(正解数/正解数)に差は見られなかった(いずれもps > 0.55,独立標本のt検定)。このタスクは覚醒時の認知機能の障害に敏感であることが証明されており、一日中の速度の差がある[F(3,120)=14.08, p < 0.001, 部分Eta2 = 0.26, 混合ANOVA 主効果]では、昼寝前と比較して昼寝後5分後にパフォーマンスが有意に低下し(ポストホックp = 0.005)昼寝後20分までにパフォーマンスが回復したことを含む。精度は同じ一般的な傾向を示したが、統計的有意性には達しなかった [F(3,120) = 2.45, p = 0.07, 部分Eta2 = 0.06, 混合ANOVA 主効果]。速度、精度ともに、グループ間相互作用(ps > 0.66)、タイムポイントとグループ間相互作用(ps > 0.44)の主効果は認められなかった。数値的には、予測通り、昼寝+群よりも昼寝+群の方が大きな減少を示した。各タイムポイントにおいて、各グループ内でさらに探索的な分析を行った。昼寝から覚醒した後、昼寝前と比較して、昼寝-群では速度[t(19) = 2.62, p = 0.02, ペアードt検定]と精度[t(19) = 2.44, p = 0.02, ペアードt検定]が有意に低下していた(図1h)。一方、昼寝+では、速度[t(24)=1.06,p=0.30,ペアードt検定]や精度[t(24)=0.28,p=0.78,ペアードt検定]に有意な障害は見られなかった(図1h)。昼寝後20分までに、両群とも5分評価と比較してDST速度が増加した [昼寝-: t(20)=3.43,p = 0.003; 昼寝+: t(24)=5.07,p < 0.001; 対のt検定]。昼寝前の速度と比較して、昼寝+は有意に速かった(t(24)=2.68,p=0.01,対のt検定);一方、昼寝-は回復したが、その速度は改善しなかった(t(19)=1.06,p=0.30)。昼寝後35分までに、昼寝+群は、昼寝前に比べて改善した速度を維持した[t(22)=2.04,p=0.05,ペアードt検定];昼寝-群は、速度に有意でない改善を示した(t(18)=1.91,p=0.07,ペアードt検定)。精度は、どちらのグループでも、昼寝前と比較して、昼寝後20分または35分の時点では有意差を示さなかった(すべてのps > 0.11)。全体的にこれらの結果は、主観的な報告(KSS評価)と客観的なパフォーマンス(DST)測定の両方に基づいて、昼寝+参加者では有意な睡眠慣性の兆候はないことを示唆している。一方、昼寝+の参加者は、昼寝+と同様の眠気の知覚レベルにもかかわらず、DST課題で認知機能の低下を示したことから、昼寝+の参加者は、昼寝から覚醒したときにパフォーマンスに関連した睡眠慣性をより多く経験していることが示唆される。しかし、これらの結果は有意でないオムニバス試験後のポストホック探索的分析であり、これらの結果は注意して解釈されるべきであることに注意が必要である。
昼寝の練習/制限は昼寝に関連したパフォーマンスプロファイルを変化させるか?
次に、4週間の昼寝練習または制限の効果を調査した。昼寝実践条件の参加者は、平均して週に3.13回(SD = 0.90)の昼寝を行った。週間の夜間睡眠記述統計(アクチグラフィによる)を補足表4に示す。以下に報告された統計量は、3(訪問1/2/3)×2(昼寝+/昼寝-)×2(練習/制限)の混合モデルANOVAからのものである。
行動パフォーマンス
知覚学習については、我々の予測とは対照的に、訪問または介入(練習/制限)の主効果はなく、有意な相互作用もなかったため、昼寝の練習も制限も時間の経過とともにパフォーマンスを変化させなかったことを示している。しかし、昼寝頻度群の主効果[F(1,36) = 5.61,p = 0.02,部分Eta2 = 0.14,混合ANOVA]はあったが、これは、訪問全体で昼寝崩壊よりも昼寝+の方が常に昼寝でより多くのパフォーマンス向上を示したことを明らかにした[ただし、訪問1と同様に、各訪問での群間の差は伝統的な統計的有意水準には達しなかったが、中程度の強度の効果の大きさを示したことに注意してほしい[訪問2: t(38) = 1.83,p = 0.074,1.83。 83, p = 0.074, Cohenのd = 0.58, 独立標本のt-検定; 訪問3: t(38) = 1.83, p = 0.075, Cohenのd = 0.58, 独立標本のt-検定] 図2a)。言い換えれば、第1訪問時(ベースライン)に観察された差のあるパフォーマンスアウトカムは、研究期間中ずっと維持されていた(図2b)。各訪問では、昼寝前の閾値は、昼寝+と昼寝-の間で同等であった(すべてのps > 0.20)。
図2 昼寝練習/制限介入。(a) 訪問2と訪問3でのTDT閾値の差
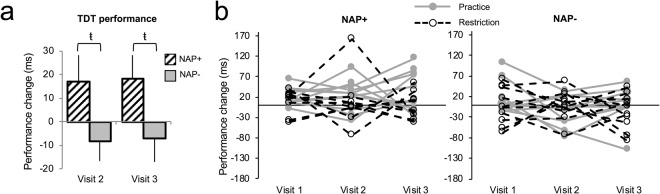
昼寝+(n = 20)は常に改善を示し、昼寝-(n = 20)は改善しなかった。b)TDT上の個々の参加者のパフォーマンスは、昼寝頻度グループ内の訪問をまたいでプロットされている。それは、昼寝依存性の記憶改善の大きさは、4週間の昼寝練習(灰色の線)または昼寝制限(黒の点線)介入にわたって安定していることが視覚的に明らかである(昼寝+練習n = 11,昼寝+制限n = 9,昼寝-練習n = 10,昼寝-制限n = 10)の両方のグループ(昼寝+練習n = 11,昼寝+制限n = 9,昼寝-練習n = 10,昼寝-制限n = 10)。ŧ'”はp≦0.07を示す。エラーバーは±1 SEM。
日中の睡眠アーキテクチャ
昼寝総睡眠時間(p = 0.88)ステージ1(p = 0.49)ステージ2(p = 0.88)徐波睡眠(p = 0.57)またはREM(p = 0.67)(報告されたp値は、3×2×2の相互作用からのものである)分について、介入をまたいで差がないことがわかった(補足表5)。同様に、昼寝の練習/制限の関数としての紡錘体密度(ステージ2 C3-C4-P3-P4avg、p = 0.30)またはパワースペクトル(NREM SO:p = 0.64,NREM delta:p = 0.76,REM theta:p = 0.89)の変化は認められなかった。これらの結果は、昼寝頻度の短期的な変化が日中の睡眠のアーキテクチャに有意な影響を及ぼさなかったことを示している。
主観的な眠気と睡眠慣性
4週間にわたって、昼寝の習慣や制限の関数として、昼寝から目覚めた後の主観的な眠気に変化は見られなかった(p = 0.56)。DSTによって測定された昼寝後の睡眠慣性の程度は、昼寝の習慣または制限による変化はなく、訪問期間中も安定していた(Δ速度:p = 0.57;Δ精度:p = 0.19)。
議論
この若くて健康なサンプルである自己申告の昼寝(昼寝+)は昼寝で知覚学習を実証したが、自己申告の非昼寝(昼寝-)では昼寝でのパフォーマンス向上は見られなかった。重要なことに、これら2つのサブグループを同じ分析(昼寝 vs. 覚醒)で組み合わせた場合、昼寝は覚醒対照群と比較して、このタスクの知覚学習に有益であるように見え、先行研究を再現している2,43。しかし、さらに分析を進めると、この結果は昼寝+の人によってもたらされたものであり、すべての昼寝が同じではないという事実を覆い隠していることが明らかになった。さらに、2つのグループのパフォーマンスは、睡眠中の異なる振動特性と関連しており、行動パフォーマンスと紡錘体密度、および行動パフォーマンスとSOパワーとの間の対照的な関連性を示していた。最後に、4週間の実験的な昼寝制限や練習は、どちらのグループのパフォーマンスプロファイルを変化させるのに十分ではなく、昼寝+の人の昼寝制限は昼寝から得られる認知的利益を減少させず、昼寝参加者の昼寝量を増加させても昼寝に関連した知覚学習を増加させることはなかった。これらの知見は、昼寝嗜好が測定可能な向上をもたらすためには、より長い訓練期間が必要であるか、あるいは昼寝嗜好は経験に依存しないことを示唆している。これらの結果は、睡眠研究者だけでなく、管理者、教育者、政策立案者、職業的な昼寝を推奨する臨床家など、幅広い人々に影響を与える可能性がある。
習慣的な昼寝の定義については合意されていないが、先行研究では、昼寝を頻繁に行う人の睡眠構造は軽度の睡眠段階が優勢であるのに対し、昼寝頻度の低い昼寝はより多くの徐波睡眠を含む昼寝と関連していることが明らかになっている25,44。我々は以前に、昼寝の睡眠アーキテクチャに対する昼寝の用量依存効果を報告している。今回の研究では、McDevittら28と比較して、このサンプルでは事前の昼寝頻度の範囲が小さかったためか、マクロアーキテクチャー睡眠ステージの違いは見られなかった。しかし、昼寝+の参加者では睡眠ステージ2のスピンドルが増加しており、スピンドル密度は昼寝+のグループのみでより良いパフォーマンスと関連していることがわかった。この相関は、訓練された標的部位とは反対側の大脳半球で発生したスピンドルに対して特に強かった。この結果は、潜在的に学習に関与する脳領域に局在する紡錘体の活動を反映している可能性がある33,45が、頭皮脳波の限られた空間分解能とこの研究で使用された電極の数を考慮することが重要である。それにもかかわらず、これは有望な結果であり、今後の研究では、視覚知覚学習における睡眠紡錘体の役割について研究を進めるべきであることを示唆している。
この種の学習におけるスピンドルの役割は何であろうか?スピンドルの活動は、事前の昼寝の好みが特定されていない研究4,48-51や、主に定期的な昼寝者24,44をテストした研究を含め、記憶課題の広い範囲でより良いパフォーマンスと相関している46,47。スピンドルは、新たに学習した情報の再生や再活性化を反映して、脳ネットワーク間の情報伝達やシナプス結合の強化や修正につながると考えられている(アクティブ・システム・コンソリデーション・モデル)46。知覚学習の領域では、シグマ帯(すなわちスピンドル周波数)のパワーの増加は、以前にヒトではパフォーマンスの向上と関連していた33。視床と一次視覚野との間で情報を中継するためにはスピンドルの振動が重要であり、最終的にはV152の睡眠依存性の応答変化につながっていた。このように、紡錘体は、情報を再活性化し、知覚学習ネットワークのシナプス結合を強化する統合プロセスの電気生理学的マーカーである可能性があり、行動性能の向上につながる。この重み付けの変更は、低レベルの視覚領域、高レベルの意思決定ユニット、またはその両方で起こっている可能性がある53-55。
昼寝+の人における正の紡錘体相関と有意な中和は、第2期睡眠に特有のものであった。研究では、ステージ2のスピンドル48,49,56,57,徐波睡眠のスピンドル51,またはNREM睡眠中のスピンドルを合わせたもの4,50,58,59のいずれとの記憶関連性を報告しているかで異なる傾向がある。知覚学習の領域における先行研究では、第2期睡眠中の紡錘体の活動が重要な役割を果たしているが、徐波睡眠ではないことが明らかになった33。なぜ、徐波睡眠のスピンドルではなく、ステージ2のスピンドルが、本研究で検討された睡眠に基づく統合とより強く関係しているのだろうか?Genzelら39のモデルによると、軽度のNREM睡眠(すなわちステージ2)は大域的な情報交換と能動的な増強を促進するが、徐波睡眠はシナプス単位の局所的な弱体化(シナプス恒常性、後述)を促進するという。彼らのモデルでは、スピンドルはグローバルなリプレイイベントを駆動しない(cf.60)。むしろ、スピンドルはこれらのリプレイイベントの直後に、先行するリプレイイベントに関与する大脳皮質ネットワークの局所的なシナプス可塑性を引き起こすのである。このモデルを今回の結果に拡張すると、第2段階では、視床皮質と視床皮質のグローバルなコミュニケーション(例えば、視床と視覚野の間のリプレイ)52 と皮質シナプスの可塑性(例えば、視覚野の長期増強)61 を促進することで、視覚知覚学習の統合が促進されるのではないかと推測される。
また、睡眠中には他の統合メカニズムが働いている可能性もある39。例えば、先行研究では、睡眠なしでテクスチャ弁別課題の反復訓練を行うと、一般的に知覚が低下することが示されており、NREM睡眠と一緒に昼寝をとることで低下を逆転させることができるが、ベースライン以上のパフォーマンス向上にはレム睡眠が必要であることが示されている43,62-65。注目すべきは、今回の研究では、昼寝参加者の昼寝後のパフォーマンスはセッション間で有意に悪化しておらず、昼寝パフォーマンスは知覚学習に重要であることが実証されている低速周波数帯のパワーと相関していたことである31。このことは、昼寝参加者が紡錘体に関連したパフォーマンスの向上がなかったとしても、低速波に関連した知覚維持の恩恵を受けた可能性があることを示唆している。これは、睡眠中の低周波徐波活動が、覚醒中に増強されたシナプス結合をダウンスケールさせるという仮説である66,67。学習の面では、SHYは、覚醒中の符号化が関連する皮質ネットワークのシナプスの増強を増加させ、その後の睡眠中に、低速波がシナプスの重みを「ダウンスケール」し、弱いシナプスが強いシナプスよりも相対的に多くのダウンスケールを受けていると仮定している。このプロセスは、強いシナプスと弱いシナプスの間のS/N比を増加させ、その結果、記憶処理を改善することが提案されている68。同様のダウンスケーリングのメカニズム31を介して知覚の劣化を軽減するために、遅波が重要である可能性がある。
先行研究では、レム睡眠がテクスチャ識別タスクのパフォーマンス向上に重要であることが示されている2,34。今回の研究では、昼寝後に学習を示したのはnap+のみであった。nap+ (n = 20) と nap- (n = 18) の両方の参加者の大多数はレム睡眠を持っており、グループレベルでは同等の量のレム睡眠を持ってたが、レムシータ活動は、おそらくグループ間のレム睡眠に質的な違いがあることを示していた。具体的には、REMシータパワーはNAP+で増加し、NAP+のみではパフォーマンスの向上と相関していた。シータ振動は覚醒時の記憶処理に関与していることが知られている69が、現在では睡眠中の記憶の再処理に関与していることを示唆する証拠が増えている。脳波のシータ活動は、単語のペア70と感情的な絵を学習した後に増加した71。他の研究では、睡眠中に記憶の再活性化の手がかりを提示した後にシータが増加したことが示されている72,73。また、睡眠前にどのような情報を学習したかを解読するために、レム前頭葉シータと時間シータは高い識別力を持っていた74。これらの結果から、昼寝+の参加者における紡錘体とシータの結果は、昼寝がこれらの人の記憶の再処理に関連する神経機構に関与する可能性を高めている可能性を示唆している。さらに、これらのデータは、NREM睡眠中の統合プロセス(スピンドルなど)とレム睡眠中の統合プロセス(シータなど)の両方が知覚学習に寄与し、Stickgoldらによって提案された2つのプロセスモデルを拡張していることを示唆している36。
我々の仮説に反して、4週間の昼寝練習介入は、興味のある4つのアウトカム(昼寝のアーキテクチャ、行動パフォーマンス、睡眠慣性、主観的な眠気)のいずれにも変化をもたらさなかった。我々の介入が十分な時間ではなかったか、あるいは十分な練習を必要としなかった可能性がある(最低20分、週3回、4週間)。実際、人が習慣を形成するのにかかる時間にはかなりのばらつきがある75 。ある研究では、摂食、飲酒、活動のいずれかの行動を形成するのに平均66日(範囲18~254日)という結果が出ているが、我々の介入の期間はこれよりも有意に短かった。しかし、習慣形成の研究では、環境中での手がかりを与えられた反応の自動性を測定しているため、頻繁な昼寝による認知的利益の測定とは異なる可能性があることに注意が必要である。今回の研究では、習慣形成の枠組みで行動の自動性(例えば、昼寝を「楽しみにしている」「昼寝を「頼りにしている」と感じるようになったかどうか)を調べることができるような自己報告を収集しなかった。しかし、4つのアウトカムに関しては、実践群に割り当てられた昼寝をした人のパフォーマンスに変化の傾向は見られず、介入が延長されたとしても変化の軌道に乗っていなかったことを示唆している。1つの潜在的な注意点として、介入全体の夜間睡眠の信頼できる推定値を得ることができなかったということがある。これは2つの要因によるものと考えられる。第一に、通常の睡眠スケジュール(2時間の就寝時間と2時間の起床時間の範囲内)に従うように求めた。このように、参加者は夜間の睡眠スケジュールをより意識しており、完全に「自由に生きる」状態にはなっていなかった。このため、昼寝の習慣や制限による夜間の睡眠特性の変化を検出する能力が制限されていたのかもしれない。第二に、アクチグラフ装置は頻繁に誤作動を起こし、その結果、かなりの欠落が生じ、推論統計を実行するのに十分なデータが各セル内に存在しなかった(補足表4参照)。したがって、昼寝の習慣や制限が夜間の睡眠にどのような影響を与えたかについての我々の解釈は、記述統計に見られるパターンに限定されており、注意を払ってアプローチする必要がある。それにもかかわらず、これらのデータから浮かび上がってくる興味深いパターンは、既存の昼寝嗜好とは異なる介入群の参加者(例えば、昼寝制限条件の昼寝+の人)が、研究期間中に夜間睡眠時間に変化を示したことである。昼寝を制限された昼寝+の人は、一晩の平均総睡眠時間が~17分増加したのに対し、昼寝を実践した昼寝+の人は~16分減少した。しかし、昼寝を制限されていた昼寝+の人では同様の減少が見られたが、昼寝を制限されていた人、すなわち、既存の昼寝嗜好と一致していた人では、昼寝の練習を続けていた昼寝+の人では、~2分の増加にとどまった。このことは、夜間の睡眠が日中の睡眠を補うために変化している可能性を示唆している。しかし、この質問に答えるために特別に設計された研究は、今回の研究よりも有益であろう。
昼寝の好みの個人差については、どのような説明が考えられるのであろうか?遺伝学がその役割を果たしている可能性が高く、一つの候補は時計遺伝子PERIOD3であり、可変数タンデムリピート多型を含む。この多型は、朝/夜型嗜好、遅発性睡眠相症候群、遅発性波動、睡眠喪失に伴う覚醒能力と関連している76。昼寝もこれらの要因に関連しており、PERIOD3は人々の昼寝表現型の1つのマーカーである可能性があると推測している。もう一つの可能性としては、発達初期に発生する昼寝習慣が成人の習慣に影響を与えている可能性がある23。最近の研究では、昼寝が就学前の子どもの学習に重要であり、昼寝から制限された子どもには学習が発生しなかったことが報告されている24。データを詳しく調べてみると、パフォーマンスの低下は、昼寝を制限された習慣的な昼寝者にのみ明らかであり、昼寝を制限されていない者にはパフォーマンスの低下は見られなかったことが明らかになった。この研究から浮かび上がった作業仮説は、習慣的な昼寝をする人は頻繁な統合の必要性が高く、これは発達段階での脳の成熟に関係している可能性があることを示唆している。就学前の昼寝習慣が成人の昼寝習慣とどのように関連しているかは不明であるが、この仮説を拡張して、成人の学習戦略には機能的な違いがあり、それによって認知負荷に差が生じ、一日中の睡眠の必要性が増加する可能性があることを示唆している。生涯にわたる昼寝パターンを追跡する縦断的研究は、昼寝習慣がどのように発達し、時間の経過とともに変化するか(あるいは変化しないか)を理解する上で有益であろう。もう一つの関連する疑問は、睡眠依存性の統合の違いが夜間の睡眠にまで及ぶかどうかということである。1つの可能性としては、(上述したように)睡眠を調節するさまざまな要因のために、昼寝をしている人の昼間の睡眠中は圧密化メカニズム(視床皮質同期46や紡錘体屈折77など)が最適化されず、圧密化は夜間に最もよく行われるということが考えられる。最後に、今回の結果が高齢者や臨床サンプルなどの他の集団にどのように一般化するか、また、昼寝が健康や幸福に関連する他のアウトカムにどのように影響するかを明らかにするためには、さらなる研究が必要である78。
研究方法
参加者
睡眠障害、神経学的、心理学的、またはその他の慢性疾患の既往歴のない18歳から35歳までの健康で非喫煙の成人83名(51F)がインフォームドコンセントを得て本研究に参加した。すべての実験手順は、カリフォルニア大学リバーサイド校の人間研究審査委員会によって承認された。方法は、すべてのガイドラインおよび規則に従って実施された。
以下に説明する実験的昼寝プロトコルには58人が参加し、残りの25人は1日のWake対照群の一部であった。過度の昼間の眠気(ESSスコア>10)または極端な慢性型(rMEQ<8または>21)を持つ潜在的な参加者を除外するために、Epworth Sleepiness Scale(ESS)79およびreduced Morningness-Eveningness Questionnaire(rMEQ)80を使用した。すべての参加者は、定期的に午前2時までに就寝し、午前10時までに起床し、1晩の合計睡眠時間は平均して7時間以上であると報告した。カフェインのヘビーユーザー(1日3食以上)は、実験中に重大な離脱症状が起こる可能性を除外するために登録されなかった。それにもかかわらず、1人の参加者がカフェインの離脱症状を経験したと報告し、分析から除外された。
一般的な手順
この試験は5週間のプロトコルで、1週間の自宅でのベースラインモニタリングと4週間の実験が含まれていた。参加者は、実験期間の最初(第1訪問日)途中(第2訪問日)終了(第3訪問日)にそれぞれ1日ずつ、2週間(14+/-2日)の間隔をあけて3日間の実験室内試験を行った。試験は以下のように進められた。(i)ベースライン週、(ii)ベースライン週の終わりにin-lab Visit 1,(iii)実験群の割り当て(昼寝練習または制限)はVisit 1の終わりに行われた(下記の昼寝練習・制限の項参照)(iv)昼寝練習・制限の後の2週間の実験、(v)in-lab Visit 2,(vi)昼寝練習・制限の後の2週間の実験、(vii)in-lab Visit 3であった。Wake群の参加者はVisit1のみを完了した。
試験中、参加者は習慣的な睡眠-覚醒スケジュールを維持することに同意した(上記参照)。睡眠スケジュールの遵守は、ベースライン週を含む研究期間中、毎日のオンライン睡眠日記とアクチグラフリストモニター(Actiwatch Spectrum、Respironics)で追跡された。参加者には、試験日の24時間前から試験日を含む24時間、カフェイン、アルコール、およびすべての刺激物の摂取を控えるように求めた。
試験当日の手順
各研究日、参加者は午前9時にカリフォルニア大学リバーサイド校のSleep and Cognition Labに集合した。アクチグラフィデータを確認することで睡眠スケジュールが守られていることを確認した後、参加者はテクスチャー弁別課題(TDT)のセッション1を完了した。
午後12時30分に、睡眠の標準的な睡眠ポリグラフ(PSG)記録のための電極が取り付けられた。すべての参加者は午後1時30分から3時30分までの間に2時間の昼寝の機会を与えられ、合計睡眠時間を90分まで取得した。総睡眠時間が90分に達した後、または2時間の昼寝の機会の時間が経過した後、現在の睡眠段階に関係なく、実験者がドアをノックして寝室に入ることによって昼寝を終了させた(「昼寝終了」)。寝室の照明は消灯したままで、参加者は、昼寝後の脳波および心電図の測定が行われている間、眠りに落ちることなく5分間仰臥位で横になり続けるように求められた。参加者が昼寝窓の間に30分以上連続して目を覚ましていた場合、参加者は寝室から削除され、昼寝が終了した。
降下減算テスト(DST)29,30は、睡眠の慣性による認知機能を調べるために使用された。タスクは、昼寝前に1回(〜午前11時)昼寝後に3回、具体的には5分、20分、35分後の “昼寝終了 “の4つのタイムポイントで管理された。5分間のDSTタイムポイントは、参加者が仰臥位で横になっている間に、暗い寝室で実施された;この評価の後、照明が点灯され、参加者は自由に座って寝室の周りを移動することができた。この手順の目的は、昼寝後の睡眠慣性の初期評価(5分間のタイムポイント)に続いて、参加者が通常の昼間の状態に戻ったときの睡眠慣性の消失を測定するための2つの後続の評価(20分および35分間のタイムポイント)を得ることであった。
午後5時(セッション2)に、参加者はTDTで再テストを受けた。参加者はまた、研究日中に3回、(i)セッション1の終了時(〜午前11時)(ii)昼寝後10分(〜午後3時40分)および(iii)セッション2の開始時(〜午後5時)にカロリンスカ眠気尺度(KSS)81を完了した。セッションの間(~午前11時~午後12時30分、昼寝参加者は午後4時10分~午後5時、Wake参加者は午前11時~午後5時)参加者は研究室を出て通常通りに一日を過ごしたが、昼寝(アクチグラフィで確認)運動、カフェインやアルコールの摂取をしないように指示された。Wake群の参加者は、昼寝をしなかったほか、DSTタスクや10分後のKSSを完了しなかった。
昼寝頻度群
昼寝をしている人としていない人をどのように分類するかについて、研究間の合意が得られていなかったため、我々は、自己評価(アンケートの回答に基づく)と昼寝の直前の証拠(1週間の睡眠日誌とアクチグラフィ)に基づいて区別することを選択した。昼寝習慣に関する情報は、複数の方法で入手した。まず、研究登録前の電話またはオンライン調査のスクリーニング質問票で、参加者に「日中に昼寝をする?もしそうならば、週に何回昼寝をする?また、どのくらいの時間昼寝をする?次に、研究開始前のベースラインの週に参加者の睡眠日誌に報告された昼寝の回数をカウントし、アクチグラフィデータを確認することで、これらの昼寝が行われたことを確認した。我々は、昼寝+を週に少なくとも1回の昼寝を報告した人々として定義した[平均1.54(SD = 1.03)週あたりの昼寝]、および昼寝-を週に1回未満の昼寝として定義した[平均0.18(SD = 0.40)週あたりの昼寝;すなわち、昼寝をしたことがないか、または月に1~2回しか昼寝をしない28,44。2つの情報源が一致しない場合(例えば、参加者がスクリーニング調査で昼寝をしないと報告したが、研究の前の週に昼寝をしたと報告した場合など)実験者が昼寝の習慣について聞き取り調査を行い、決定した。例えば、参加者が昼寝をしないと報告していたが、その週に病気で昼寝をしていた場合、病気は通常の出来事ではなかったため、参加者は昼寝の状態を維持していた。
昼寝の習慣と制限
これらの昼寝頻度群のそれぞれの中で、昼寝+と昼寝-の参加者は、昼寝実施群と昼寝制限群のいずれかに無作為に割り付けられた。昼寝実践群の参加者は、研究の残りの4週間は少なくとも週3回、20分以上の昼寝をするように指示された(研究訪問2と3の研究室での昼寝は、1週間の昼寝の合計にカウントされた)一方、昼寝制限群の参加者は、研究訪問中に研究室での昼寝を要求されない限り、昼寝をしないように指示された。これらの条件の遵守は、睡眠日誌とアクチグラフィのチェックによって確認された。昼寝+制限群の1人の参加者が、病気のために2週目に1回昼寝をした;この参加者のデータは解析に保持された。
睡眠ポリグラフ(PSG)
PSGデータはAstro-Med Grass Heritage Model 15アンプとGrass Gammaソフトウェアを用いて収集した。8個の頭皮脳波(EEG)と2個の電気頭文字(EOG)電極を、リンクされていない対側乳腺(F3/A2,F4/A1,C3/A2,C4/A1,P3/A2,P4/A1,O1/A2,O2/A1,LOC/A2,ROC/A1)に参照し、筋緊張を測定するために2個の筋電図電極をあごの下に装着した。PSGデータは256Hzでデジタル化され、RechtschaffenとKales82の睡眠ステージング基準に従って30秒エポックで視覚的にスコア化された。睡眠構造変数には、ステージ1,ステージ2,徐波睡眠(徐波睡眠)および急速眼球運動(REM)の分数および割合、ならびに総睡眠時間(TST)睡眠潜時(SL)および睡眠効率(SE)が含まれた。参加者は、最初の昼寝中に入眠しなかった場合(2名)または3回の昼寝のうち2回のTSTが20分未満、SEが35%未満の場合(1名)に除外された。
脳波データは、BrainVision Analyzer 2.0(BrainProducts、ミュンヘン、ドイツ)およびMatlab 2011b(MathWorks、ナティック、マサチューセッツ州)を用いて前処理および解析を行った。脳波データは0.3~35Hzの間でバンドパスフィルタリングされ、潜在的なバックグラウンドノイズを除去するために60Hzのノッチフィルタも使用された。アーティファクトと覚醒を伴うすべてのエポックは、目視検査によって識別され、拒否された。睡眠紡錘は、Wamsleyら57によって開発されたウェーブレットベースのアルゴリズムを用いて、ステージ2と徐波睡眠の間に自動的に検出された。要するに、脳波信号は8パラメータの複素モーレットウェーブレットを用いて時間周波数変換を受けた。約10~16Hzの周波数範囲に対応するウェーブレットスケールを抽出し、Wamsleyらの57のしきい値化アルゴリズムを用いて紡錘体の検出に用いた。紡錘体検出後、紡錘体密度は、離散的な紡錘体イベントの数を、各頭皮脳波電極部位における各睡眠段階での滞在時間で割ることによって計算された。個々のチャネルのデータは、チャネルが目視検査に基づいて信頼性がない(例えば、記録中に剥離した)と判断された場合に除外された。
パワースペクトル密度(μV2/Hz)は、50%のオーバーラップを持つ睡眠の連続した3秒エポックにハニングウィンドウを適用して、高速フーリエ変換(FFT)によって計算した。スペクトルパワーは以下の周波数帯について求めた。ステージ2,徐波睡眠、NREM(S2+徐波睡眠の組み合わせ)レム睡眠中の0.5-1Hz(低速振動;SO)1-4Hz(デルタ)4-8Hz(シータ)8-12Hz(アルファ)12-15Hz(シグマ)およびベータ(15-30Hz)。
テクスチャ判別タスク(TDT)
参加者はKarniとSagi38によって開発されたものと同様のテクスチャ識別タスク(TDT)を行った。TDTの視覚刺激はPsychophysics Toolbox83,84を用いて作成した。各刺激は2つのターゲットを含んでいた:中央の文字(’T’または’L’)と、画面中央から2.5°~5.9°の偏心で4つの象限(左下、右下、左上、右上)のうちの1つにある周辺線(垂直または水平方向)の配列である。象限は、参加者と訪問者の間でカウンターバランスされた。周辺配列は、水平方向に配向された背景ディストラクターの背景に対して水平または垂直に配列された3本の対角線バーで構成され、ターゲットと背景の間にテクスチャーの違いを生じさせた。
実験的試行は、以下の4つの画面のシーケンスで構成されていた:中央固定十字、33ミリ秒のターゲット画面,0~600ミリ秒の間の空白画面(刺激間インターバル、またはISI)17ミリ秒のマスク、次の試行の前に応答時間間隔(2,000ミリ秒)とフィードバック(250ミリ秒、間違った試行には聴覚ビープ音付きの赤色固定十字、正しい試行には緑色固定十字)が続いた。参加者は、中央固定時の文字(’T’または’L’)と3本の対角線(水平または垂直)の周辺配列の向きを2回キーを押すことで、1回の試行で2つのターゲットを識別した。中央課題では眼球運動をコントロールした。
各ブロックは25回の試行で構成され、それぞれが同じISIであった。閾値は、ISIが600ミリ秒から始まり0ミリ秒で終わるように、徐々に短いISIを用いて、13のブロックにわたる性能から決定された。セッション全体のISIの特定のシーケンスは[600, 500, 400, 300, 250, 200, 167, 150, 133, 100, 67, 33, 0]であった。各ブロックの正答率のサイコメトリー関数をワイブル関数でフィットさせ、パフォーマンスが80%の精度をもたらすISIを決定した。TDTパフォーマンスはセッション1とセッション2の間の閾値の差として計算され、正のスコアはパフォーマンスの改善(すなわち、セッション2での閾値の低下)を示し、負のスコアは悪化を示す43,62。
参加者にはタスクの指示が与えられ、研究開始前のオリエンテーションでタスクの練習を行った。この練習では、末梢ターゲットは研究中には使用されない象限に配置された。この練習は、参加者がタスクを理解していることを確認し、参加者が初めてタスクを実行するときに一般的に発生する一般的なタスク学習に起因する訪問順の影響を減らすことを目的とした。さらに、各研究日には、実際のタスクを開始する前に、参加者に簡単なバージョンのタスク(ISIは1,000-600ms)を練習させ、周辺のターゲットを90%から100%の間で正しく識別できることを確認させた。
降順減算課題(DST)
このタスクでは、精神的な計算スキル29,30を探りながら、ワーキングメモリにかなりの負荷をかけることで、短時間(3分間)の認知機能を測定する。最初に、実験者は参加者に3桁の数字、例えば「865」を与え、これを参加者が声に出して繰り返した。次に、参加者には、865から9を暗算し、その答え(856)を声に出して言うように指示された。この数字が、サブトラエンドを引く新しい最小値となった。サブトラエンドは、それが2の値に達するまで1ずつ減少し、その後9に戻りました。したがって、次の試行では、参加者は856から8を引いたはずである。参加者には、できるだけ多くの試行を完了するために3分が与えられた。参加者には、できるだけ早く正確に作業するように指示が出された。課題は、実験者がクリップボードに取り付けられた紙に参加者の回答を書き、書かれた回答が参加者に見えないようにして、声に出して実施された。参加者は、任意の回答を修正することができ、実験者に助けを求めたかどうかを推測するように指示された。正解した回答の総数と、回答総数に占める正解数の割合を、それぞれ速度と正確さの指標として計算した。差のスコア(例えば、昼寝後5分-昼寝前のパフォーマンス)を計算し、ΔaccuracyおよびΔspeedとして報告した。
データの削減と統計解析
最初のTDT閾値(すなわち、Visit 1, Session1)が平均値から2.5標準偏差以上離れていた参加者は、外れ者としてフラグが立てられた(本実験ではn=2,Wake群ではn=2,全員が平均値から2.5標準偏差以下のパフォーマンス不良者でした)。主実験では、睡眠不足とカフェイン離脱のために削除された2人の参加者と一緒に、2つのパフォーマンスの外れ値は、サンプルのTDTのパフォーマンスの下位4人でした。したがって、我々は均等なトリム手順を採用し、また、上位4人の訪問1,セッション1の実行者(天井にあった)を削除した。これにより、Visit 1のデータが分析された48人の参加者が残った(nap+ n = 26;nap- n = 22)。また、Wakeグループに均等なトリミングを適用し、パフォーマンスの悪い2人の外れ者と上位2人のパフォーマーを削除して、Wakeグループに21人の参加者を残した。昼寝群の参加者のうち1人は、初めてタスクを実行したとき(Visit 1 training)にタスクを理解していなかったため、DST分析から除外され、精度差スコアが平均値より3標準偏差以上高くなった。
昼寝介入実験に参加した48人のうち8人は、研究終了前に脱落または脱退した[病気(n = 1)研究と相反するスケジュールの変更(n = 3)睡眠覚醒スケジュールの不遵守(n = 2)および不明な理由(n = 2)]ため、3回の訪問すべてを完了した40人を残した。実験を完了した参加者のうち、21人が実践群(11回昼寝+、10回昼寝-)19人が制限群(9回昼寝+、10回昼寝-)に割り振られた。最終的なサンプルの人口統計学およびその他の特徴は、補足表1に報告されている。
スピンドルの結果については、我々は、スピンドル(特に高速スピンドル)が最大であることが知られている中心-頭頂部領域に焦点を当て85-87と各半球内の中心および頭頂部電極(すなわち、C3-P3avgとC4-P4avg)上で検出されたスピンドルの数を平均化した。大脳半球の違いが明らかでなかった場合、我々は、グランド平均(C3-C4-P3-P4avg)を使用していた。パワースペクトル分析のために、前頭部と中央部の間のパワーの有意な地形の違いのために(しかし、半球の違いはない)我々は、前頭部(F3-F4avg)と中央部(C3-C4avg)の電極を平均化した。
第1訪問時のnap+とnap-の間の差は、独立標本のt検定を用いて検定した。訪問先間の変化は、訪問先を反復測定とし、2つの参加者間因子を用いた混合モデルANOVAを用いて検定した。昼寝頻度群(昼寝+/-)と介入群(練習/制限)の2つの因子を用いて混合ANOVAで検証した。TDTにおけるパフォーマンスの変化の大きさを、差スコアを用いた1標本のt検定を用いて、ゼロ(すなわち、変化なし)と比較した。二変量ピアソン相関では、TDTパフォーマンスと訪問1日目の昼寝の特徴との関連を調べた。相関の数を減らすために、NREM(ステージ2+徐波睡眠の組み合わせ)とREM睡眠段階のパワースペクトルに焦点を当て、各睡眠段階(NREM:SO、デルタ、シグマ、REM:シータ)に関心のある特定の周波数帯を選択した。節度を検定するために、我々は、NAP+群とNAP-群の相関を別々に実行し、その後、Fisher r-to-z変換とz-検定88を使用して相関係数の有意差について検定した。3回の訪問でのスピンドルとパワースペクトルの分析については、混合モデルANOVA(上記参照)を使用し、特に訪問1の間に有意差および/または中和を示した変数のみをテストした。
この研究の縦断的な性質のため、昼寝記録中の不良電極、実験者のミスによるKSSスコアとDSTデータの欠落、時計の誤作動によるアクチグラフィデータの欠落など、セッションと訪問のデータが欠落している。さらに、すべての参加者が昼寝中の睡眠のすべての段階を持っていたわけではなかった(徐波睡眠はなかった。訪問 1 n = 3,訪問 2 n = 4,訪問 3 n = 4; レムなし:訪問 1 n = 10,訪問 2 n = 4,訪問 3 n = 5)。) このように、自由度は分析によって異なる。
