Contents
Information and the Origin of Qualia
www.ncbi.nlm.nih.gov/pmc/articles/PMC5399078/
要旨
この論文では、クオリアは局所的な皮質ネットワークにおける情報の処理の結果である可能性が高いと主張している。情報ベースのアプローチを用いて、情報構造(脳内の情報の物理的具現化、主に活動電位のパターン)と情報メッセージ(脳にとってのそれらの構造の意味、クオリアの基礎)を区別している。この論文では、これら2種類の情報の間の形式的な関係を発展させ、情報構造がどのようにメッセージを表すことができるのか、情報メッセージがどのように構造から識別できるのかを示している。
この論文では、この視点を大脳皮質ネットワークやアンサンブルにおける基本的な処理に適用し、ネットワークがどのように2種類の情報の間で変換できるかを示している。この論文では、入力された発火パターンが情報メッセージとしてネットワークによって識別され、生成された発火の出力パターンがそのメッセージの表現であることを主張している。
ネットワークが注意や他の再入室プロセスによってアトラクター状態になるように促されている場合、物理的な情報がネットワークを循環するたびに識別されるメッセージは、「前回のメッセージの表現」になる。嗅覚の例を用いて、前のメッセージの上にメッセージを重ねることで、嗅覚のクオリアがどのように発生するかを示している。情報の各パスで識別されたメッセージは、内側のアイデンティティーから、内側の形、内側の類似性やイメージへと進化する可能性がある。その結果が嗅覚クオリアである。
このように、情報がネットワークの階層を通って共鳴状態で循環しても、同じ結果が得られることが示されている。クオリア生成の議論は他の感覚モダリティにも適用され、脳全体の制約の充足のプロセスを通じて、どのようにして特定の意識状態が任意の瞬間に発生しうるかを示している。
ECoGデータやガンマ振動やアトラクターの研究を用いて、理論の主要な予測を裏付ける証拠を提示し、理論を支持するためにはどのような更なる証拠が必要なのかを概説する。
キーワード:クオリア、意識、シャノン情報、意味情報、ニューラルネットワーク、アトラクター
序論
意識研究における本当に困難な問題は、主観的経験そのものの起源の問いに対する答えを見つけることである。意識というテーマで発表された膨大な数の論文の中で、ほとんどの論文は意識の組織(Baars, 1988; Dehaene er al)。 このような研究は、私たちがどのようにして経験を持つことができるのか、どのようにして脳が意識的な経験をもたらすように自分自身を組織化することができるのか、ということを明らかにするものである。しかし、これらの物理的な神経活動がどのようにして現象的な結果を生み出すことができるのかという口うるさい疑問は、ほとんど取り上げられていない。ミラーは、意識科学における主な重点は、意識の神経構成、つまり必要最小限の基質を追求することにあるべきだと主張してきた(Miller, 2014)が、それさえも(彼が認めているように)意識的経験の説明にはならないだろう。
現象的経験の起源を探る試みはいくつか行われてきた。システムがそれ自身の内部状態を反映することができる表象的再記述の考え方(Clark and Karmiloff-Smith, 1993)は、特定の種類の行動を現象的経験の特定の特性にマッピングしようとする初期の提案であり、システムがそれ自身に自身の活動を再記述することができるというラジカル可塑性のテーゼ(Cleeremans, 2008)のような考え方につながった。実際、システムが高次の表現につながる自身の内部状態を観察するメタ認知の概念全体もまた、この自己反省のテーマに従っている(Pasquali et al 2010)。自己反省はまた、主観的経験が注意のプロセスの脳内モデルであると提案する注意スキーマ理論(Graziano and Kastner, 2011)の核心にもなっており、著者らはその考えを支持するいくつかの証拠を提示している(Webb and Graziano, 2015)。
しかし、特定の神経活動が現象的な結果につながるはずの直接的なメカニズムを示す理論的な説明はない。この記事は、純粋に物理的なものを現象的なものとリンクさせるための一つの試みであり、このトピックに関する以前の記事(Orpwood, 2013)を基に構築されている。
この記事では、クオリアの生成に注目している。クオリアは多くの場合、多くの著者の定義では感覚的な経験に限定されている。しかし、この記事では、より広い視野を取り、哲学者フラナガンが好んだクオリアの定義を反映している。彼はクオリアの広い意味を主観的、一人称、現象学的な感覚を持つ経験として定義している(Flanagan, 1992)。この定義では、感覚的なクオリアは、意識を構成するすべての現象的経験のサブセットにすぎない。この定義には、これらすべての経験に共通の原因があるという前提がある。感覚的クオリアの原因は、他のすべての経験と同じで、感覚的知覚に焦点を当てているだけである。
この論文の主張は、大脳皮質ネットワーク機能の情報処理分析に強く基づいている。意識を理解するための情報アプローチは、意識状態が高度に統合され、高度に分化した情報を含むという事実によって特徴づけられると主張したTononi(2004)によって開拓されてきた。彼は、ネットワークによって生成される情報関係の定量化に有用な尺度を開発した(Balduzzi and Tononi, 2009)。しかし、この論文では、意識を情報の観点から特徴づけるのではなく、純粋に物理的な情報とクオリアの中心にある意味的な情報との関係をより直接的に探ろうとしている。ある状況下では、局所的な皮質ネットワークにおける情報の処理がクオリアにつながることを示すことを目指している。
脳の情報
二つの情報の種類
情報は神経構造の中で起こるすべてのことの核心にあるが、神経科学の研究の大部分は、情報とは何かという伝統的な工学に基づいた定義を使用している。半世紀以上前のシャノンの研究(Shannon, 1948)は、この理解の基礎を築いた。これらの手法を用いて可能となる分析の種類は、明らかに脳の情報処理能力に当てはまるものである。シャノンの分析における情報とは、物理的な情報である。通信バスのバイナリパルス、スクリーンのピクセル、ページに書かれた文字などである。脳内で行われている重要なプロセスには、もちろんこのような物理情報が関与している。膨大な量の物理情報は、細胞内の局所的な化学的・電気的変化と、細胞間でこの情報を伝達するための電気的・化学的事象を用いて生成され、伝達されている。実験的神経科学の大部分は、これらの情報生成者と伝達者のモニタリングに従事している。しかし、神経の文脈における情報の重要性には、いくつかの懸念を引き起こすと思われる別の側面があり(例えば、Pockett, 2014)それは意味情報の重要性である。意味情報とは、物理的な情報に関連付けられた意味である。それは、物理情報に具現化され、それによって伝達されるメッセージである。脳では、意味情報の重要な側面は、活動電位の発火に具現化されたメッセージである。それは、物理的情報の大規模な神経細胞間通信と処理によって伝達される意味である。神経系全体で活動電位を介して伝達されるすべての情報は、明らかに何らかの形で内面の精神世界を支えているが、これらの純粋に物理的な出来事と、それを支える豊富な内面の意味との間の関連性は、当惑させられる。しかし、この問題は神経科学の中心にある。神経細胞の発火という物理的な情報は、私たちの内なる世界の意味の意味情報とどのように関係しているのであろうか?
この理解は、意識の理解にとって極めて重要である。Chalmers (1996)は、彼の画期的な著書の中で、情報には物理的な側面と現象的な側面の両方があるという基本的な原理を提案している。私たちの意識的な内面世界は、意味で構成されている。それは、私たちが環境を知覚する方法であり、私たちが内なる思考を経験する方法である。私たちの意識的な内なる世界を構成するクオリアはすべて情報の側面であるが、それらは意味情報の側面である。それらは、私たちのニューロンの発火で行われている物理的な情報処理に何らかの形で関連している。このように、このように、「クオリア」の原因を理解するためには、ニューロンの発火の物理的な情報と「クオリア」の意味的な情報との間の関連性を探る必要がある。
この論文では、上で議論した脳内の2種類の情報に対して2つの新しいラベルを定義することで、以前の研究(Orpwood, 2013)を反映させている。これらのラベルは、両者をより強く分離することを目的としている。まず、脳内に存在する物理的な情報には「情報構造」というラベルが与えられている。情報構造とは、細胞の発火や活動電位の列の物理的な活動のことである。それらは、情報が伝達され、処理され、脳内に保存される形態である。第二に、意味情報には「情報メッセージ」というラベルが与えられる。情報メッセージとは、脳内での身体活動が何を意味するのか、それが私たちにとって何を意味するのかを表している。
情報構造と情報メッセージの変換
情報構造と情報メッセージの間のつながりを探るためには、それらの間で起こりうる変換を検討することが重要である。2つの明確な変換がある。その第一は、情報構造が何らかの形で情報メッセージを表すということである。脳内の情報構造は、神経細胞の発火という形で、何らかの形で私たちにとっての意味を表している。
例として、視覚野のV4領域(色処理領域)での発火活動を考えてみよう。脳が青色の物体に注目している場合、V4領域で発生している発火のパターンがある。これは神経活動であり、それは情報構造にすぎない。しかし、その脳にとっての情報の意味、何らかの形で解釈された情報メッセージは、「青という色」です(図(図1).1)。
ある意味では、V4のネットワークにおける発火活動の情報構造は、情報メッセージ「色は青」を表している。さらなる例として、我々が記憶と呼ぶ特定の情報メッセージを考えてみよう。これらの情報メッセージは、おそらく大脳皮質錐体細胞の可変入力感度の長期記憶の中に具現化されているのであろう。しかし、これらの可変的な感受性は記憶ではない。昨年の素晴らしい休日」などということはありえない。単なる情報構造なのだ。しかし、それらは記憶を表すことができる。大脳皮質錐体細胞の可変入力感度にある情報構造は、記憶の情報メッセージを表すことができる。情報構造は情報メッセージを表す。クオリアがどのように生成されるかを調べるためには、この表象プロセスの分析が一つの重要な課題である。
図1
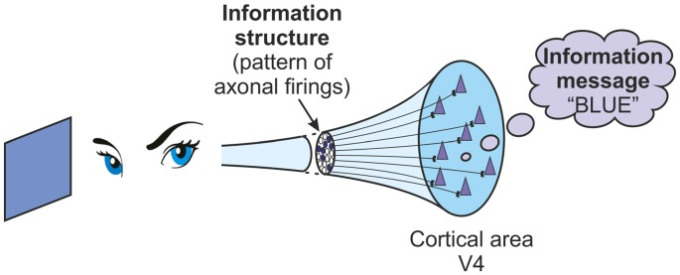
色の青に出席している脳は、その視覚野のV4領域に入力のパターンを受け取る。このパターンは情報構造である。脳にとっての「青」の意味は、情報メッセージである。
情報構造とメッセージの間の第二の関係は、メッセージが情報構造の認識と識別から生じるということである。情報メッセージは曖昧な存在であるため、この関係を想像するのは容易ではない。最初は単一のニューロンを考えてみよう。ニューロンは、入力の中の空間的に分散したパターンを認識できることが示されている(Mel, 1992; Orpwood, 1994)。ニューロンが学習した入力パターンを受信すると、ソーマの脱分極が大きくなる。認識が重要であればあるほど、ソーマの脱分極は大きくなる。最終的に脱分極は閾値に達し、細胞は発火する。発火は、この特定の入力パターンが認識されていること、認識されていないことに 対して認識されていることの二項決定を外界に伝えることを表している。しかし、認識するニューロンは入力パターンをどのような方法でも識別することはできない。認識されたパターンはすべて同じ結果、つまり発火につながる。入力パターンは、ニューロンにとって「前に見たことがある」以外の意味を持つことはできない。
ニューロンのネットワークでは、状況は大きく異なる。このような場合には、「入力パターン」の特定のセットに基づいて訓練されたネットワークを考えてみよう。 ネットワークは入力構造を認識しているが、それは構造があるときには応答を生成し、構造がないときにはほとんど生成しないからである。しかし、ネットワークはまた、その特定の入力構造が受信されるたびに典型的な出力応答を生成しているので、入力構造を識別している。ネットワークは、それが訓練された入力構造を受信したときだけ、その特徴的な応答を生成する。これは、ネットワークが、学習済みの入力構造を受け取るたびに、典型的な出力応答を生成しているからである。入力はネットワークにとって何らかのアイデンティティを持っており、そのように認識され、出力はそのアイデンティティを表している。このように、ネットワークは、入力された情報構造を認識して識別することができれば、メッセージとして解釈することができる。
この分析は、情報構造と情報メッセージの間の相互作用が双方向であることを示している。メッセージは、表現のプロセスを通じて構造体に変換されることができる。構造体は、識別のプロセスを経てメッセージに変換することができる(図22)。
図2 情報構造とメッセージの相互関係
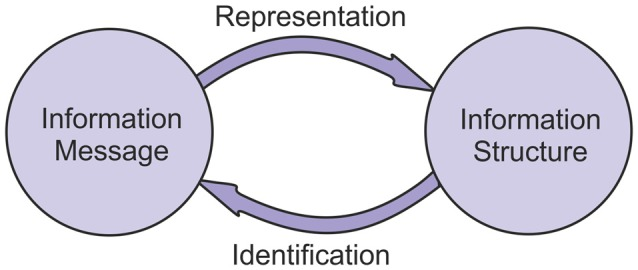
情報構造体は情報メッセージを表し、情報メッセージは情報構造体から識別できる。
情報構造体は、情報送信者から情報受信者に伝達されることができる。送信者は情報構造を生成することができ、その情報構造は送信者にとって何かを表している。例えば、送信者が音に反応していて、短いノイズにさらされていたとすると、送信者は情報構造を生成することで応答することができる。この構造は送信者にとってノイズを表すことになる。送信者はその構造体を情報受信者に送信することができるが、送信されるのは構造体だけである。それは、情報メッセージをそれと一緒に運ぶことはない。送信者が生成した構造体は、わずか数ビットである可能性がある。それらのビットが受信機によって受信されたとき、それらのビットは受信機にとって何を意味するかわからない。ノイズを意味する場合もあるが、数字の「2」や三角形の形を意味する場合もあるし、「去年の素敵な休日」を意味する場合もある。すべては、受信機がどのように構成されているかに依存する。構造は送信者にとってはノイズを表していたが、受信者にとってはどのようなメッセージを表すこともできる。したがって、情報送信者から情報受信者への通信は、メッセージではなく構造体の形でしかありえない。
したがって、これまでの情報処理の議論から、いくつかの基本的な結論を導き出すことができる。
- 情報は構造体やメッセージの形をとることができる。
- 脳の身体活動は情報の構造を扱います。
- 私たちの内なる意識世界のクオリアは情報のメッセージである。
- 構造はメッセージを表す。
- メッセージは構造から識別できる。
- 構造はメッセージではなく、送信者から受信者に伝達される。
ネットワーク情報の変換
基本的なネットワーク動作
この論文では、脳内の基本的な情報処理主体は、個々のニューロンではなく、ニューロンのアンサンブル、あるいはこの論文でラベル付けされているようなネットワークであるということが重要な前提となっている。個々の細胞ではなく、ニューロンのダイナミクスと情報処理における機能との間のつながりの鍵を握るのは、ニューロンのアンサンブルであるという確信が高まっている(Harris, 2005; Buzsáki, 2010; Bharmauria et al 2016)。このようなアンサンブル/ネットワークの存在を示す最近の証拠については、後述する。大脳皮質のこれらのネットワークは、多数の錐体細胞とそれを支える介在ニューロンで構成されており、これらの細胞は入力の弾幕に反応して協調的に行動している。このようなネットワークは、入力のパターンを認識することができ、その認識に応答して自らの発火活動の出力パターンを生成することができる。
最初に、大脳皮質における基本的なニューラルネットワーク/集合体の動作を考えてみよう。このネットワークは、上で説明した情報構造とメッセージ間の変換の両方を示すことができる。このネットワークの入力では、情報構造を構成する活動電位のパターンを受信している。もし入力された情報構造が認識されると、ネットワークは反応し、その中のいくつかのニューロンが発火する。この発火がネットワークの出力を構成し、この出力は別の情報構造である。この出力構造は、ネットワークへの入力の同一性を表している。これはメッセージであり、入力の意味を表している。この同一性は、ネットワークが行った学習や、遺伝的に構成された方法に依存している。もちろん、メッセージは意識的なものではない。それは要約アイデンティティに過ぎない。しかし、入力構造が特定されれば、情報メッセージが得られる。つまり、基本的なネットワークは、情報の受信者であると同時に、情報の送信者でもあるのである。基本ネットワークは、入力された構造を識別の過程でメッセージに変換することができ、そのメッセージを表現の過程で発信構造に変換することができる。
このような全体的な基本的なメカニズムは、情報構造体を受信する大脳皮質ネットワークで行われているものである。入力情報は構造体である。ネットワークがその入力構造を認識して識別すると、出力情報構造が生成される。その出力情報構造は、ネットワークへの入力構造の情報メッセージ、アイデンティティを表している。このように、基本的な皮質ネットワーク内では、情報構造と情報メッセージの関係をより厳密に定義することができる。出力構造は、ネットワークがその入力構造を識別したときに得られるメッセージを表す。構造からメッセージへの変換から再び構造への変換がある(図(図33))。
図3 ニューロンのネットワークまたはアンサンブルにおける基本的な情報処理
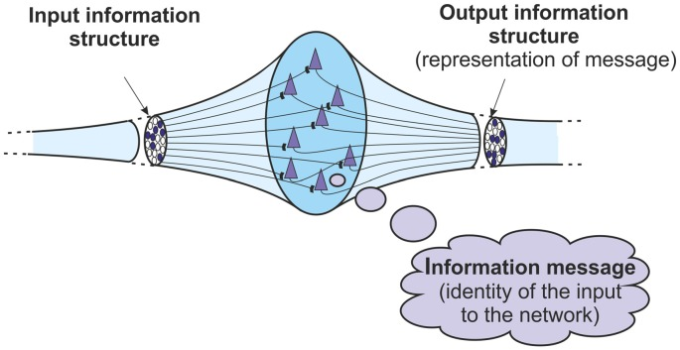
出力情報構造は、入力情報構造から識別された情報メッセージを表す。情報は、構造体からメッセージへ、そして再び構造体へ
ネットワーク間の通信
ネットワークは、階層化されたネットワークのチェーンの一部である可能性がある。鎖の中の各ネットワークについては、上で説明したのと同じプロセスが行われる。ネットワークはその入力構造を認識し、ネットワークへの入力の同一性を表す出力構造を生成することができる。その構造は、チェーン内の次のネットワークに伝達される。上述したように、通信されるのは構造だけである。チェーン内の次のネットワークは、それが受けた事前学習に応じて、その構造を認識し、それをメッセージとして識別することができる。このプロセスは、ネットワークのチェーンに沿って繰り返すことができる。
ネットワークはまた、その出力を再び入力にフィードバックすることができる。この局所的なフィードバックにより、生成されたばかりの表現をネットワークにフィードバックすることができる。ネットワークがこのフィードバックを認識すると、表面的には自分の表現を認識しているように見えるであろう。しかし、ネットワークの観点からこの状況を理解することが重要である。観察者にとっては、ネットワークは自分自身の表現を受け取り、それを識別している。しかし、ネットワークにとっては、自分自身の表現を受け取っているにもかかわらず、もちろんそれを知ることはできない。ネットワークにとっては、それはただ別の入力パターンを受信しているだけであり、それはそれまでの経験に応じて識別できるかどうかを判断しているだけである。前述したように、ネットワークは情報の送り手であり、情報の受け手でもある。情報送信側の部分が情報構造を生成し、それを情報受信側の部分にフィードバックする場合、伝達されるのは情報構造だけである。伝達されるメッセージはない。情報送信部は、この構造体が「表現」というアイデンティティを持っていることを情報受信部に伝えることはできない。
表現を表現として識別する能力は、クオリアの起源の問題の核心にある。結局のところ、我々がクオリアと呼ぶ情報メッセージは内部表現である。色のクォーレは、検出された光の色相の内部表現である。匂いのクォーレは、検出された揮発性分子の構造の内部表現である。表象は、単なる別の基本的なアイデンティティとは対照的に、どのようにして「表象」のメッセージとして識別されるのであろうか?表現がどのようにして「表現」のメッセージとして識別されうるのかについてある程度の洞察を得るためには、メッセージがどこから来るのかに焦点を当てる必要がある。ネットワークにとっては、出力が何を表していようと、それがメッセージであるということしか言えない。ネットワークにとってのメッセージは、単に出力が表現しているものである。では、出力が表現を表すことは可能なのであろうか?出力が表現を表すことができるのであれば、メッセージは「表現」でなければならない。これは、ネットワークがアトラクター状態に落ち着いたときに可能になることを以下に説明する。
アトラクター状態
これは、あるネットワークが、その出力を再び入力にフィードバックしていると考えてください。このような活動が行われると、ネットワークはアトラクタ状態になる可能性がある。このような安定したアトラクタ状態に落ち着くと、ネットワークの出力構造は入力構造と同じになる。ネットワークは、生成された各出力構造が、それを導いた入力構造と同じであるという循環的な活動状態に入る。この状態では、出力が入力と同じであるため、ネットワークは何もしていないように見える。伝達関数は一様に見える。しかし、これはそうではない。入力された情報構造を処理するために複雑な変換が行われている。各ニューロンはパターン認識を行い、ネットワーク内の他のニューロンと相互作用して、たまたま入力と同じ 発火パターンで結論を出しているのである。
では、このようなアトラクター状態に落ち着くと、どのような影響があるのであろうか?このように、このように、「ネットワーク」というものは、「ネットワークからの出力は、入力の同一性の表現である」ということだけが述べられている。しかし、アトラクタ状態では、出力は入力と同じである。それらは同じ構造である。つまり、この状態では、出力はそれ自身のアイデンティティの表現になるのである。これはなかなか難しい概念であるが、このアトラクター状態では、出力構造はその同じ出力構造の同一性の表現になるということである。アウトプットは、アウトプットの同一性の表現である。しかし、ネットワークにとっては、出力はメッセージの表現でもある。つまり、出力はメッセージの表現の同一性の表現になる。このやや複雑な文は、メッセージについて何かを教えてくれる。上で議論されたように、出力が何を表していようと、それがメッセージである。したがって、この文から、メッセージはメッセージの表現の同一性の表現でなければならない。情報がアトラクター状態でネットワークを介して循環されるように、毎回得られるメッセージは、最後のメッセージの表現の同一性である(図4参照).4)。メッセージは、前のメッセージの表現の同一性である。表現を表現として識別する能力がクオリアの起源の問題の核心にあることは上で述べたとおりである。この能力は、安定したアトラクター状態では実現されることがここで示されている。観察者にとっては、情報がネットワークを介して循環するときはいつでも、ネットワークが前のメッセージの表現を識別していることは明らかである。しかし、アトラクタ状態が落ち着くまでは、ネットワークはこのことを知ることができない。ネットワークにとっては、アトラクタ状態で毎回識別されるメッセージは「前のメッセージの表現」である。
図4
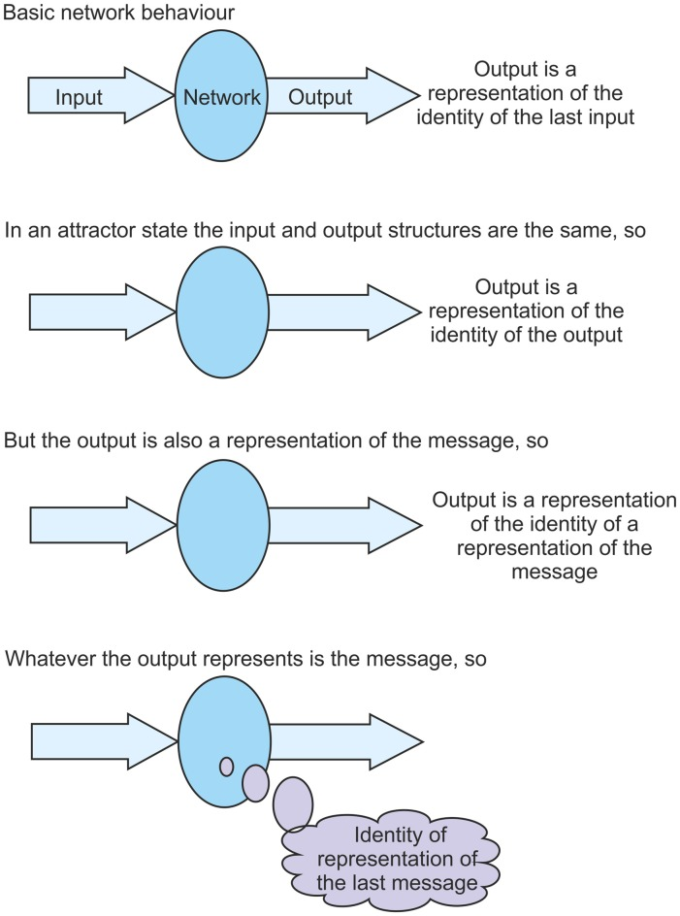
提示された理論の重要な構成要素は、定点アトラクター状態では、ネットワークは自分の表現を表現として自分にフィードバックすることができるということである。この図は、それがなぜそうなのかを明らかにすることを目的としている。この図は、アトラクタ状態では、情報がネットワークを介して循環するにつれて、ネットワークは、各パスでフィードバックされた入力を前のメッセージの表現として識別することができることを示している。
アトラクター状態になってから、ネットワークを介して情報が循環し始めると考えてみよう。ネットワークが最初に入力構造を受け取り、それを認識した場合、その入力構造はネットワークに対して何らかのアイデンティティを持ち、それを「メッセージ」として認識する。これは、ネットワークへのメッセージを表す出力構造を生成する。その出力構造がフィードバックされると、ネットワークはフィードバックを「前のメッセージの表現」として識別する。元のメッセージは「メッセージ」であり、2番目のメッセージは「そのメッセージの表現」である。循環が続けば、3番目のメッセージも「前のメッセージの表現」、言い換えれば「元のメッセージの表現の表現」となる。メッセージは各サイクルで不変であるが、その性質は進化している。この状況は、ネットワークがアトラクター状態を達成したときにのみ発生するが、それはいくつかの非常に深遠な特性につながる。
ここには、表面的には提示された議論を却下しているように見える複雑さがある。アトラクター状態についての議論では、関係するネットワークの性質の非常に単純なイラストを使用している。このように、アトラクターにつながるフィードバックは、神経モデリングの用語では、非常に単純化されている。出力情報構造は、ネットワークの出力から再びその入力へと変化せずに転送されると考えられている(図(Figure5A).5A)。しかし、もちろん現実には、このフィードバックと関係する接続は非常に複雑である。フィードバック活動の空間パターン、発火軸とサイレント軸索のマトリックスは、出力活動が入力にフィードバックされると必然的に変化する(図5B).5B)。しかし、ネットワークの活動の境界は固定されていない。単純なモデリング用語では、出力はネットワーク・モデルから出てくるものであるが、ネットワーク・モデルの境界は拡張することができる。境界をネットワークへの入力の直前まで拡張し、その時点での構造として出力を定義すれば、その時点までのフィードバック接続が複雑であるにもかかわらず、出力構造はそのまま入力に供給されることになる(図(Figure5C).5C)。重要なのは、その時点での出力が、やはりネットワークのメッセージの表現であるということである。もちろん、その時点での出力が、それを導いた入力と同じであることは、アトラクター状態が達成されたときだけである。
図5
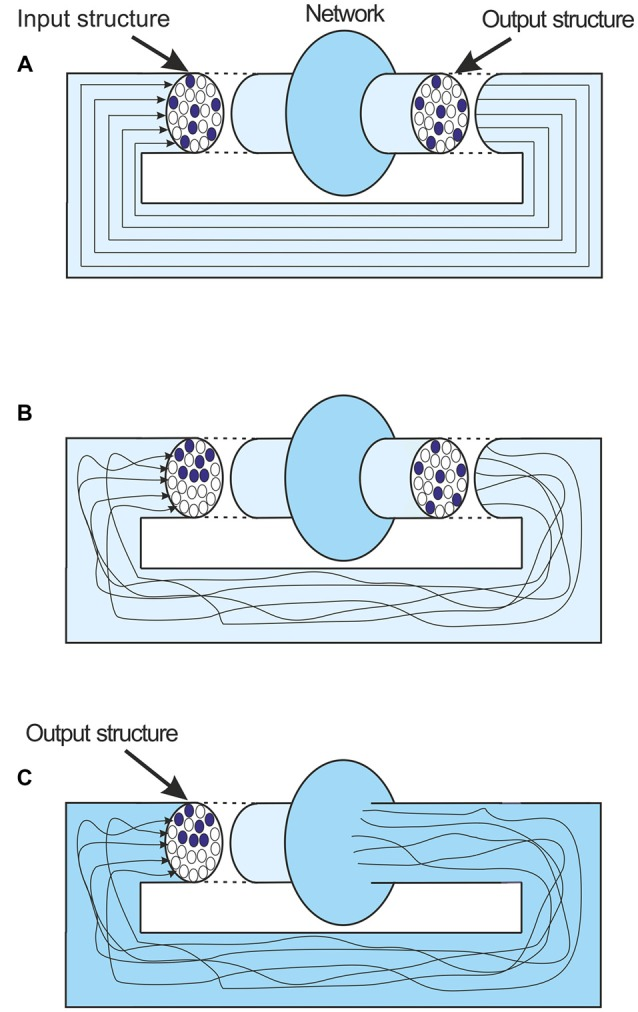
(A) ネットワークにおけるローカルフィードバックの理想化された描写。フィードバックされても出力構造は変化しない。
B)より現実的な描写。フィードバック軸索は複雑な経路をたどり、出力構造とは全く異なる入力構造につながる。
(C) ネットワークの境界がフィードバック入力の直前まで拡張されている。このように、この図では、このような情報は、ネットワークの中では、「出力」と「新しい入力」の両方に変化はない。重要なのは、出力が最後のメッセージの表現であることである。
アトラクター状態によって提供される追加の機能がなければ、ニューラル認識器は、受信した入力を常にゼロから開始する。ニューラル認識器は、その入力を事前学習に依存したものとして認識することしかできない。アトラクター状態があれば、これはすべて変わる。各フィードバックに対して、ネットワークはその入力を最後のメッセージの表現として認識する。各認識の結果として生じる新しいメッセージは、前のメッセージの上に構築され、注目すべき結果をもたらす。
共鳴ループ
もう一つの神経解剖学的実体があり、その行動はアトラクター状態と一致し、それによって処理される情報について同様の結論を導くことができる。この実体は、ネットワークの共振ループである。共鳴ループのアイデアは、グロスバーグが彼の適応共鳴理論(Carpenter and Grossberg, 1987)の中で開拓したものである。これは、感覚処理階層のような階層内の異なるレベルのネットワーク間のフィードフォワードおよびフィードバック相互作用に適用された。階層の1つのレベルのネットワークの活動は、より高いレベルにフィードフォワードされる。高次のレベルでは、事前の経験に基づいて、フィードフォワード入力の同一性について判断を行い、その判断結果を元のレベルにフィードバックして、合意があるかどうかを確認する。合意のレベルに応じて、フィードバックパターンは、フィードフォワードパターンとフィードバックパターンの両方が変化しなくなるまで修正される。その時点で、以前の経験に従った入力の同一性のより高いレベルの判断は、正しい。Grossbergは安定性のこの状態を共鳴と呼んだ。合意が達成されるまでネットワークの応答を適応させるために関与するメカニズムは、Grossbergによってfolded-feedbackと呼ばれた(Raizada and Grossberg, 2003)。感覚階層内のレベル間の相互作用は、予測符号化理論の核心にもなっている(例えば、Clark, 2013)が、その同一性に関するフィードフォワード活動とフィードバック判断の差を反映した誤差関数を減らすために適応が行われる。共鳴ループに関するこれらの考え方は、知覚のプロセスによく対応しており、感覚的クオリアはこれらの 共鳴プロセスにリンクしていると期待されている。
図66は、共鳴ループで行われる中核的なプロセスを示している。この図では、知覚プロセスはこれよりも複雑である可能性が高いが、2 つのネットワークが相互に連結しているだけである。提示される議論のためには、この複雑さは重要ではない。このように、「ネットワーク1」は、「ネットワーク1」への入力が「ネットワーク1」によって識別され、その識別の出力表現がフィードフォワードされる。このフィードフォワード活動は、再び入力を識別できるネットワーク2への入力となり、アイデンティティの出力表現を再びネットワーク1にフィードバックすることができる。この反復活動は、合意に達し、2 つのネットワークからの出力が不変の構造に安定するまで続けることができる (図 (Figure6A).6A)。元の入力の同一性について行われる知覚的判断を支えるのは、この安定した共振状態である。
図6
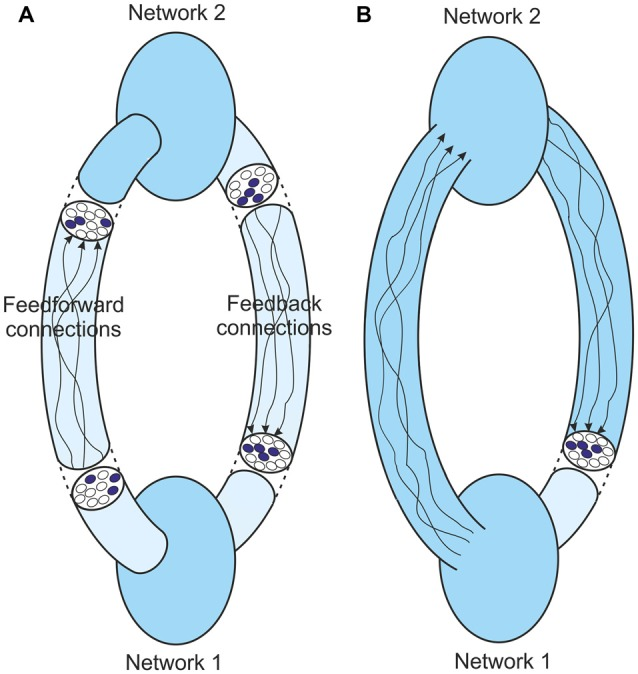
(A) 共振時の2ネットワークループのフィードバック。システムの異なる点での構造は一定のパターンに落ち着くが、フィードフォワードとフィードバックの経路は複雑で、異なる点では全く異なる安定した構造になる。
(B) ネットワーク1の境界を入力の直前まで拡張した同じシステム。共鳴時には、このネットワークへの入力はその出力と同じである。重要なのは、出力がネットワーク1で得られた最後のメッセージの表現であることである。
この安定した共鳴状態は、上述した定点アトラクターのダイナミクスと多くの類似点を持っている。単一皮質ネットワークと同様に、ネットワークの境界は、ネットワークの出力とその最終的なフィードバック入力の間に介在する複雑さを取り除くために拡張することができる(図(図6B).6B)。ネットワーク1への最終的なフィードバックは、この拡張された境界からの出力である。非安定状態では、ネットワーク1にどのような入力が提供されようとも、この境界からの出力は異なる。安定状態では、ネットワーク1にこの特定の入力が与えられるたびに、同じ出力が生成される。つまり、安定状態では、この出力はネットワーク 1 への入力の同一性を表現している。
したがって、ネットワーク1を分離して考えることができる。安定した共振状態では、それはアトラクターのような働きをしている。出力は入力の同一性を表現している。しかし、安定した状態では、出力はそれを導いた入力と同じである。したがって、出力は出力の同一性の表現である。そして、その出力は最後のメッセージの表現である。したがって、その出力は、最後のメッセージの表現の同一性の表現なのである。それがネットワークにとってのアイデンティティである。前に議論したように、ネットワークに対する同一性は、出力によって表現されたものであれば何でもよい。したがって、ネットワークに対するアイデンティティは、最後のメッセージの表現のアイデンティティでなければならない。安定した共振状態では、情報がネットワークを介して循環するため、ネットワークへの入力の同一性は、最後のメッセージの表現の同一性となる。この結果は、共振ループ内の各ネットワークにも適用される。
つまり、ネットワークにおける情報処理の結果を要約すると、通常、ネットワークは入力を特定の「メッセージ」としてしか識別できない。しかし、フィードバックを伴う二つの状況では、これは変化する。
- 1つ目は、単一のネットワークで安定した定点アトラクター状態を達成する場合。
- 2つ目は、いくつかのネットワークが連結された安定した共振状態を達成した場合である。このような状況では、ネットワークは入力を「前のメッセージの表現」として識別する。
再エントリー活動の重要性
クオリアの生成を検討する前に、ネットワークがアトラクター状態になるように、あるいはネットワークの階層が共鳴状態になるように、どのようにして促されるかを簡単に見ておくことが重要である。情報の一回の通過だけを受けるのではなく、関係するネットワークはローカルフィードバックを介してこの活動を繰り返すように促される必要がある。この能力の鍵を握るのがリエントラントフィードバックのプロセスであり、多くの著者がその影響を探ってきた(例えば、Edelman, 1992; Lamme and Roelfsema, 2000; Bullier, 2001; Pascual-Leone and Walsh, 2001)。基本的な原理は、大脳皮質に入ってきた情報は、最初に感覚野の領域の階層を経て、前頭前野にフィードフォワード伝達されるというものである。高次の領域のネットワーク状態が情報を生成し、それがフィードバックされて、入ってきた感覚の流れを調整し、統合する。意識的な知覚の発達は、このリエントラント活動に大きく依存しているという実験的な証拠が増えてきている(例えば、Haynes et al 2005;Silvanto et al 2005;Boehler et al 2008)。例えば、Lammeは、前頭前野から生じるリエントラントフィードバックが視覚知覚に重要であると主張している。前頭前野の活動は、視覚野からの最初のフィードフォワードの流れに続く注意の活性化の一部である(Lamme, 2010)。Reesはまた、意識的な視覚が行われるためには、このような高レベルのリエントラント活動が必要であると主張している(Rees et al 2002: Rees, 2007)。Boly er al)。 (2011)はまた、意識には前頭皮質からのトップダウンの投影が必要であることを示している。しかし、Lamme (2010)は、意識的な知覚もまた、視覚野内のリエントラント活動から生じる可能性があると主張している。(Boehler et al 2008)は、この局所的なリエントラント活動の考えを支持して、感覚情報の早期認識は、前頭前野のフィードバックを伴うにはあまりにも急速であることを示している。
Orpwood (2013)は、このリエントラントフィードバックがアトラクター状態の発達に重要であると論じている。リエントラント刺激によって提供されるファシリテーションがなければ、ネットワーク活動、特に非常に局所的なフィードバックを伴う活動はすぐに衰退し、アトラクターは発生しない。リエントラント・フィードバックの刺激があって初めて、アトラクターが落ち着くのに十分な時間、局所的な活動の循環が起こるのである。さらに、リエントラントフィードバックは共振ループの基礎となると考えられている。Lamme and Spekreijse (2000)は、それが知覚組織の重要な要素であることを示している。したがって、アトラクター状態とレゾナント状態の両方において、それらの発達を促し、形作り、振り付けをするためには、リエントラントフィードバックが必要なのかもしれない。
クオリアの生成
Orpwood (2013)は、ネットワークの入力を、それまでの入力の同一性の表現として識別する能力がクオリアの生成につながる可能性があると主張したが、その論文で提示された議論は複雑すぎて、あまり明確ではないと感じられるようになった。また、安定したアトラクター状態の場合のみを扱っていた。現在の論文では、クオリアが発生するためには、実際には2サイクルのフィードバック情報の通過が必要なだけであり、そのプロセスは安定した共振状態にある相互リンクされたネットワークをも巻き込むことができると主張している。
上で議論した情報処理がどのようにクオリアにつながるかは、例を使うと理解しやすいかもしれない。嗅覚の状況を考えてみよう。嗅覚の知覚は、その情報に基づいて、その情報がどのように反応しているのかを知ることができる。嗅覚受容体ニューロンはこの曝露に反応し、嗅球でこの情報が局所的に処理されると、嗅道に沿って発火パターンが生成される。この発火パターンが情報構造である。嗅覚野と眼窩前頭前野のどちらが臭気を意識する部位なのか(Shepherd, 2007)おそらく両方なのかについては議論があるが、この議論の目的には関係ない。嗅覚路に沿って行われている情報構造が必要な皮質領域に到達していると仮定する(図(Figure7).7)。この領域に入力構造を認識するニューロンのアンサンブルがある場合、細胞の認識ネットワークは、それ自身の発火の出力パターン、つまり出力構造を生成する。もし、生成された出力構造があるならば、その構造はネットワークへの入力の同一性を表していなければならない。このように、「ネットワーク」と「ネットワーク」の間には、「ネットワーク」と「ネットワーク」の間の関係は、「ネットワーク」と「ネットワーク」の間の関係を表している。出力構造は、それを供給するセンサーが硫化水素にさらされているときにのみ、その出力構造を生成する。出力構造はネットワークにとっては H2S を表現しているが、もちろん意識的な意味ではない。それは、検出された化学物質のネットワーク自身の描写に過ぎない。それは、このネットワークにとってのアイデンティティ「H2S」を体現した構造体である。嗅覚皮質ネットワークが構成されているため、出力構造は、検出された化学物質のネットワーク独自の表現である。このネットワークに対するアイデンティティ「H2S」の物理的な具体化である(図77)。
図7 嗅覚クオリアの生成
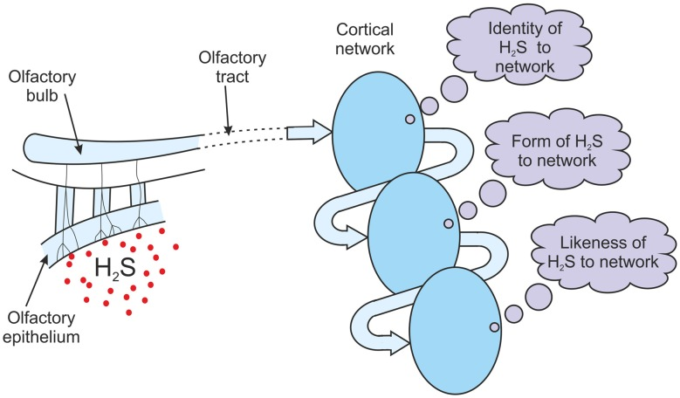
H2Sへの曝露後、嗅覚管からの情報構造は皮質の嗅覚ネットワークを介して循環する。注意がこのネットワークのアトラクター状態につながる場合、情報の各パスで得られるメッセージは、前のメッセージの上に構築される。メッセージは、H2Sの内側のアイデンティティから、H2Sの内側の形、H2Sの内側の類似性やイメージへと進化する。H2Sらしさは経験される。
リエントラントフィードバックの項で上述したように、アトラクター状態または共鳴状態のいずれかの発展を促すこの嗅覚入力に何らかの注意が払われていることが想定される。出力構造は、直接、または連鎖や階層の他のネットワークを介してフィードバックされる。フィードバックは、アイデンティティ「H2S」を表す物理構造であるが、メッセージとして識別される。それはネットワークにとって特定の意味を持っている。そのメッセージとは何か?アトラクターや共振状態が発達する前には、メッセージは単に新しいランダム入力のアイデンティティとなる。しかし、上で論じたように、アトラクタやレゾナント状態が発達すると、そのメッセージは「前のメッセージの表現」となる。前のメッセージがネットワークにとって「H2S」であると識別されたならば、フィードバックはその「H2Sの表現」として識別されることになる。フィードバックはもちろん物理的な構造であるが、そのフィードバックの正体は内部の意味である。その意味とは、ネットワークにとっての「H2Sの表現」であり、その化学的な描写であり、ネットワークにとっての「H2Sの内側の形」なのである。したがって、この第二の認識によって、フィードバックされた構造は、ネットワークによって、その内部形態であるH2Sの内部形態として識別される。したがって、硫化水素への暴露に反応する嗅覚皮質の場合、第1のメッセージは「H2Sの同一性」であり、第2のメッセージは「H2Sの内部形態」である(図(図77))。
フィードバックの認識の結果として、ネットワークは再び起動する。この第2の出力構造は、第2のメッセージの表現であるので、ネットワークの「H2Sの内部形態」の表現である。この出力構造は再びフィードバックされる。フィードバックは、アイデンティティである「H2Sの内部形態」を表す物理構造であるが、メッセージとして識別される。それはネットワークにとって特定の意味を持っている。そのメッセージとは何か?アトラクターや共振状態が発達しているので、そのメッセージは「前のメッセージの表現」になる。前回のメッセージがネットワークにとって「H2Sの内側の形」であったならば、フィードバックは「H2Sの内側の形の表現」として識別される。しかし、何かの形の表現とは、それの類似性、イメージである。ネットワークはそれ自身にH2Sの内的な類似性や内的なイメージを伝えているのである。フィードバックはもちろん物理的な構造であるが、そのフィードバックの正体は内側の意味である。その意味とは、「H2Sの内側の類似性またはイメージ」である。ネットワークにとっては、それはH2Sらしさである。それは、ネットワークにとってH2Sがどのように見えるか、ネットワークにとってどのようなものであるか、ネットワークにとってどのように見えるかということである。第一のメッセージは化学物質のアイデンティティであり、第二のメッセージはネットワークにとってのH2Sの形態であり、第三のメッセージはH2Sの内面的な類似性またはイメージである(図(Figure7).7)。脳がこのプロセスの結果を報告することができた場合、それは単にH2Sのその内側のフォームを報告するだけではなく、そのH2Sがそれにどのように見えたかを報告することになるだろう。それは、H2Sらしさの内的体験を識別しているのである。それは単なる要約内的感覚であり、誰にでも説明できるものではない。しかし、言葉の事前学習から、その内的経験は「匂い」というラベルを与えられることになる。脳には硫化水素の匂いというクオリアがあった。
もし脳が鼻の上皮ではなく舌の受容体に基づいた別の化学物質の感知システムを使用した場合、認識された化学物質は同様の方法で解釈されるであろう。舌のセンサーが口の中のグルコースに反応した場合、その化学物質は最初に味覚センサーからの情報構造を受信した皮質ネットワークによって識別される。最初のフィードバックでは、新しい入力はネットワークのグルコースの内部形態として識別される。第二のフィードバックでは、新しい入力はグルコースの内側のイメージとして識別される。それは、その化学物質が脳にどのように見えるかという結果につながるだろう。しかし、言葉では言い表せない内的体験は、嗅覚体験とは異なるものであった。それは脳内の異なる構造を使用するからである(クオリアが発生部位によってどのように変化するかについての議論は、Orpwood, 2010を参照のこと)。結果は、その化学物質が脳にどのように見えたか、どのように経験したかということに変わりはない。しかし、この例では、学習によって、脳はその化学物質に「味」というレッテルを貼ることになる。脳の経験は要約内的感覚に過ぎないが、言葉の知識から、その経験を甘味として報告することができる。
先ほどの青い色の情報が大脳皮質の領域V4に伝達される例に戻ると、これはどのように解釈されるのだろうか。V4 のネットワークに対する初期のアイデンティティは「青」である。ネットワーク自身はもちろんそれを「青」と解釈することはできない。ネットワークにとっては、それは要約情報メッセージに過ぎない。しかし、ネットワークの設定と学習により、ネットワークは入力された情報構造を認識する。最初の認識の結果として生成される出力構造は、ネットワーク自身の「青」の表現であり、「青」の内的な具現化である。これがアトラクター状態でフィードバックされると、ネットワークはフィードバック入力を青という色の内側の形として認識する。ネットワークは、今度は自分の青の内側の形を表現した出力を生成する。この2番目の出力がフィードバックされて認識されると、ネットワークはそれを青の内的な類似性やイメージ、あるいは青さとして識別する。それは、ネットワークにとって青という色がどのように見えたか、どのように経験したかということである。それはただの抽象的で何とも言えない概念であるが、事前の学習から脳はそれに「青」というレッテルを貼ることができる。青色のクオリアが体験されていたのである。
同様に、音叉を叩いた結果として聴覚野に入力があった場合、脳はどのように反応するだろうか?聴覚野は、ある時点でその音を認識し、その特定の音の内部表現である出力構造を生成するネットワークを含んでいるだろう。その出力がアトラクター状態でフィードバックされると、その音の形としてネットワークに認識される。その音がミドルCの音であった場合、2回目の発射は、ネットワークの内部フォームであるミドルCの表現となる。このフォーム表現がネットワークに伝達され、内部の類似性として識別される。新しいメッセージは、ミドルCがネットワークにどのように見えたか、どのようにそれを経験したかということになる。クオリアは、説明することができないが、それはミドルCの性質、ネットワークにとってミドルCがどのようなものであったか、ということで形成されただろう。しかし、クオリアを言葉で表現することを学ぶことで、それは「音」というラベルを与えることができる。ミドルCの特定の音は言葉にするのは簡単ではないが、独特のクオリアを持っているだろう。
そこで、アトラクター状態に落ち着いたネットワークに関連した内的経験のセットが、いつでも存在し得ることが提案されている。それぞれが、内的経験、見え方、クオールにつながる可能性がある。説明されているように、感覚皮質の活動に関連したクオリアがあるだろう。感情的な経験に関連した扁桃体、場所の感覚に関連した海馬の場 所細胞などで生成されたクオリアもあるだろう。アトラクター状態を達成したネットワークでの活動のみが、これらの経験につながる。上記のように、またOrpwood(2013)で議論されたように、アトラクター状態の開発は、この行動にネットワークを刺激するリエントラントフィードバックに依存している可能性がある。だから注意と他のメカニズムは、いくつかの開発し、他の人を阻害することができるアトラクター状態の開発を羊飼いになるだろう。このような制御は、活動がアトラクター状態で維持されているすべてのネットワークを含む活動のセットに落ち着く脳全体の制約の満足のようなものにつながるだろう。その瞬間に生じたクオリアの集合が、その瞬間の脳の意識状態を構成することができる。したがって、Orpwood (2013)で下線を引かれているように、これらのアイデアは、グローバル・ワークプレイス理論のようなより一般的な意識理論とかなり互換性がある。この論文との違いは、現象的経験そのものの発生メカニズムについての理論を提供しようとしている点である。
この行動には何か証拠があるのか?
クオリアが形成されると何が測定可能になると理論は予測しているのだろうか。この理論は局所的な皮質ネットワークの活動を中心に展開し、これらのアンサンブルが個々の細胞ではなく皮質内の情報処理の主要な機能単位であると仮定している。また、クオリアが形成されるための前提条件として、これらの局所的なネットワークが周期的に活動することを必要とし、クオリアが形成される時点でアトラクターや共振状態が形成されることを必要としている。これらの予測に証拠はあるのであろうか?
ニューロンのネットワークまたは集合体
脳の情報処理の鍵を握っているのは、個々の細胞ではなく、ニューロンのネットワークであるという証拠が増えてきている。10年以上前にハリスは、大脳皮質での情報処理の基本となるのは、ニューロンの集合体における発火活動の空間的なパターンであると主張した(Harris, 2005)。この考え方は、他の研究者によっても増幅されている(Buzsáki, 2010; Bharmauria er al)。 ネットワークの役割の証拠を提供するためには、理想的には集団の活動を調べる研究が必要であり、必然的に2光子カルシウムイメージングは、この研究において有用なツールであることが証明されている。(Miller et al 2014)は、覚醒マウスの視覚刺激に反応するニューロンの特定のアンサンブルを示した。同じアンサンブルの活動は、はるかに低いレベルではあるが、自然発生的に発生しており、まるでアンサンブルが事前に学習された接続ネットワークであるかのようであった。任意のニューロンは、いくつかの異なるアンサンブルの一部である可能性がある。錐体細胞アンサンブルの活動の刺激依存性は、(Hofer et al 2011)によっても示されている。彼らは、この錐体細胞の活動が、局所的な抑制性ニューロンの活動と顕著に対照的であり、より単純に局所の一般的な活動を反映していることを示した。(Cossell et al 2015)は、L2/3細胞におけるシナプス駆動の大部分は、視覚入力に対して同様の反応を示す局所細胞間の少数の強い相互入力によって提供されることを示した。また、(Lee et al 2016)は、L2/3錐体細胞が同様の方向選択性を持つサブネットワークに編成されていることを示した。これらの集団研究と同様に、個々のユニットからのデータもまた、情報処理におけるアンサンブルの重要性を示している。(Yoshimura et al 2005)は、感覚入力によって活性化されたL2/3錐体細胞のサブセットが、与えられた列の中にあることを示している。(Bharmauria et al 2016)は、すべての方向でV1の異なる機能ネットワークをリクルートし、視覚入力の方向を識別する特定の細胞集合を実証した。このようなアンサンブルは、まさにこの論文で議論されている理論に従ってクオリアを生成するために必要なビルディングブロックの種類である。ゼキの微小意識に関するオリジナルの研究(Moutoussis and Zeki, 1997)では、視覚野では色のクオリアとは別に動きのクオリアが発生することが示されており、局所的なネットワークの活動がクオリア発生の核心にあることを示唆している。この研究は、他の研究者によっても繰り返され、さらに研究されている(例:Linares and López-Moliner, 2006; Self, 2014)。また、Bablioniは、ERPの研究において、特定の皮質ネットワークにおける同期化された活動がクオリアの生成の根底にあると結論づけている(Bablioni et al 2016)。
振動活動
これらのニューロンのアンサンブルが、ここで提示された理論で必要とされるような周期的な活動に従事しているという証拠はあるのだろうか?アトラクター行動は、定義上、振動性の活動につながるが、十分な数の細胞が関与していれば、これはLFPや頭皮電位で拾うことができる。局所的なネットワークフィードバックが関与していると、非常に短いフィードバック経路が生じ、非常に速い振動を引き起こすことになる。このような時間経過は、大脳皮質でのガンマ振動の発生を研究している様々な理論的研究によって示されている(例えば、Wang and Buzsaki, 1996)。局所的なアトラクター振動が発生した場合には、ガンマ周波数帯の振動する場電位や頭皮電位で示されている可能性が高い。もちろん、すべてのガンマ振動がアトラクターの行動を示すということではない。ガンマやアルファやベータなどの皮質電位の振動は、それ自体が最終的な結果として扱われることが多いので、特定の機能を持つ特異的な現象として扱われることが多いのであるが、そうであるべき理由はない。それよりも、特定の周波数は、脳内の多くの異なる活動によって生成され、その活動のダイナミクスがたまたまその振動の周波数につながっている可能性が高いの(Merker, 2013; Bosman et al 2014)。この点は、ガンマ活動をBOLD信号と比較したMerkerによってさらに増幅された(Merker, 2016)。ガンマ周波数から推論できる唯一のことは、その高速な性質が短い伝導経路で局所的な活動を示す傾向があるということである。したがって、ガンマ周波数の活動はアトラクターの行動によって生成される可能性が高いが、ガンマの活動の発生は、アトラクター以外のことが起こっていることを示すこともできる。しかし、少なくとも、意識的な知覚が報告された場合には、ガンマ周波数領域の活動が適切な脳領域で検出され、この活動はかなり局所的なものであることが、この理論から期待される。この仮説を検証するためには、アトラクター行動から生じる可能性の高いガンマ線活動と、他のメカニズムから生じるガンマ線活動を区別する技術が非常に有用であると考えられる。脳波やMEGで測定されたガンマ波形の主要な電流源はL5ニューロンの活動に由来し、L5の活動が同時に起こるとL2/3層の活動は完全にマスクされることがモデリング研究で示されている(Lee and Jones, 2013)。大脳皮質情報処理におけるこれらのラミナの異なるフィードフォワード/フィードバックの役割(例えば、Harris and Shepherd, 2015)および意識への影響の可能性(Orpwood, 2010)を考えると、ガンマ活動の解釈は一筋縄ではいかない。
脳波およびMEG記録におけるニューロンイベントとガンマ帯活動との関連は複雑であるが(Fernández-Ruiz and Herreras, 2013)しばらくの間、知覚と関連していた(レビューはMartinovic and Busch, 2011; Rieder er al)。 初期の誘発振動は特徴分析に、後期の誘発活動は知覚理解に関連しているようである(Tallon-Baudry, 2003; Herrmann et al 2004)。Tallon-Baudry (2003)は、物体認識時には、感覚野へのトップダウンフィードバックがガンマ振動の増加と関連しており、視覚イメージによっても同様の活動が誘導されることを示した。誘発されたガンマ応答と誘発されたガンマ応答の分析から、どちらも同じような神経活動の源に依存していることが示された(Porcaro et al 2011)。また、視覚刺激は、持続的な刺激ではガンマ活動が低レベルに低下するが、刺激の持続時間中は継続することが示されている(Lowet et al 2016)。報告されている痛みのレベルは、痛み刺激に対する誘導されたガンマバンド応答を反映しているようである(Schulz et al 2012)。視覚的および音響的錯覚もまた、適切な感覚皮質におけるガンマ活動の増加をもたらしている(Kaiser et al 2006; Matsuzaki et al 2012)。ガンマ同期は知覚、特に位相同期の重要な因子として強調されており(Fries et al 2007)意識におけるアンサンブル活動の重要性が提案されていることを考えると、機能的に接続されたユニットがV1で同定されたとき、接続されていないニューロンと比較して、その応答のガンマパワーが増加したことは興味深いことである(Bharmauria et al 2015)。注意は視覚領域と他の領域の間のガンマ同期を増加させ(Doesburg et al 2008)痛みに対するガンマ応答を増加させる(Hauck et al 2007)。しかし、ガンマ帯の活動と注意との間の可能性のある関連性については、いくつかの否定的な意見がある。(Aru et al 2012)は、知覚報告が同じであるにもかかわらず、感覚的な文脈と事前情報が異なるガンマ応答につながることを発見した。
ヒトの大脳皮質表面からの直接記録(ECoG)は、ガンマ線活動と意識との関連を探るために非常に有用なツールを提供している。いくつかの有用なECoGレビュー(Jacobs and Kahana, 2010; Crone et al 2011; Lachaux et al 2012)があり、(Buzsáki et al 2012)は、神経活動とECoG記録の間のリンクについての良い議論をしており、発火活動を反映したガンマ振動を示している。ECoG技術を用いて、海馬の非常に局所的な活動が意識的認識と関連していることが判明した(Rey et al 2014)。これらの著者は、特定の概念が意識に持ち込まれると、ガンマ線の活動が局所的な細胞集合体の活動を反映していると感じてた。また、(Burke et al 2014)は、被験者が単語のリストを思い出そうとしたときに起こる活動を調査した。彼らは3つの段階を示した:高レベルのシータを含む最初の探索段階、高レベルのガンマを含む第2の認識段階、運動領域のガンマを含む第3の報告段階である。シータは検索過程を反映しているのに対し、ガンマは意識的な想起を反映していると主張している。知覚的顔認識は、覚醒した人間の反応を見るために多くの研究者によって使用されていた。認識が行われると、顔領域のガンマ帯の活動が増加することが一貫して見出されている(Lachaux et al 2005; Fisch et al 2009)。サルを対象としたECoG研究でも、動物が物体を見ることに集中したときに、視覚野上のガンマ帯の活動が大きく増加することが示された(Brunet et al 2015)。この活動は、サッカードの後に低レベルに低下し、その後再び高レベルに戻る。Gaillard et al 2009)は、言葉の意識的知覚を調べた。意識的な刺激のみが高出力ガンマにつながり、非常に局所的なガンマ同期を示した。ガンマ反応は刺激提示中も持続していた。シーケンス学習もECoG技法を用いて探索されている(Madhaven et al 2015)。ガンマパワーの増加は学習過程で発生したが、記憶力が向上するとガンマパワーは減少し、新しいシーケンスでは再び増加につながった。ガンマパワーは、シーケンスが学習されると減少する意識的な内容を反映していると考えられる。ECoGデータを用いて、(Van Vugt et al 2014)は、感覚的な知覚は感覚野で同期したガンマ活動として維持されており、PFCからのトップダウン活動によって減衰するが、再活性化することができると結論づけている。注意もまた、これらのECoG反応と関連している。Lachaux et al 2008)は、注意がシフトすると、ある領域ではガンマ帯信号のパワーが減少し、別の領域では増加することを発見した。(Ray et al 2008b)は、感覚刺激に注意を払うと聴覚野と体性感覚野の両方でガンマ帯のパワーが増加し、それが起こるとPFCよりも増加することを示した。
最近の論文の中には、より高い周波数の活動をブランケット用語で表現しないことの重要性を強調し、低周波数と高周波数のガンマが異なる反応を反映しうることを示したものもある。Crone et al 2011)は、高頻度ガンマの方が範囲が限定されており、より確実に認識反応にリンクしていること、また、発火速度の変化よりも局所的な同期にリンクしていることを論じている。上で議論した注目に連動したガンマパワーの増加も、主に高ガンマ周波数であった。Hermes et al 2015)は、高レベルの視覚知覚(顔、建物)は高い周波数のガンマとのみ相関し、低い周波数のものは相関しないことをさらに示した。低い周波数のものは、グレーティングによってのみ確実に誘発されることができた。また、高レベルの活動は、刺激の持続期間中も継続していた。興味深いことに、マカクザルでは、深さ電極を使用して、発火活動が低周波ではなく高周波のガンマにつながることがわかった(Ray et al 2008a)。一般的にECoGの文献では、低周波の脳波波形がより広く、基本的な処理に関連していることが示されているようである。例えば、Groppe et al 2013)は、アルファは感覚処理と注意に関連しており、ベータは非常に広く、ガンマは顕著に観察されるが、それは非常にまれであり、局所的であることを示唆している。
アトラクター
提示された理論の一本鎖は、皮質ニューロンのアンサンブルがアトラクター行動をとることを要求している。意識と結びついた振動波形は有用な指標であるが、大脳皮質でアトラクターが形成されるという直接的な証拠はあるのだろうか。アトラクター行動が多くの認知機能を支えているという暗黙の仮定は、20~30年前から存在していた(Hopfield, 1982; Amit, 1989; Rolls and Treves, 1998)。局所的な皮質ネットワークの構造と、その錐体細胞の反復的な側副連結性は、アトラクター行動を発達させるのに理想的に適していることを示唆している。多くの理論研究がこの枠組みを用いている。例えば、Tsodyks(1999)は、海馬の機能に関する理論を記述しており、アトラクターを用いて海馬の場所マップを定義している。Capaday et al 2013)は、運動皮質の組織についての議論の中で、大量の再帰性コラテラルがあることで、運動皮質が運動データを表現するためのアトラクターを生成することが可能になると結論づけている。また、意識に関するいくつかの理論的研究は、アトラクターの生成を中心に構築されてきた(例えば、Mozer, 2009)。意識の内容は、計算モジュールの相互接続されたネットワークの中で展開される過渡的なアトラクターに対応していると提案された(Mathis and Mozer, 1996)。グロスバーグは、意識状態はボトムアップとトップダウンの情報の間で発展する共鳴状態のサブセットであり、それが彼の適応的共鳴理論(Grossberg, 1999)の中心にあると主張した。
このように広く理論的に考えられているにもかかわらず、アトラクター行動のための証拠はまだ現れ始めたばかりである。その主な理由は、この行動がニューロンのネットワーク全体の活動に依存していることと、大規模な細胞集団をモニターする技術が利用可能になるまで証拠を待たなければならなかったことにある。これらの研究における重要な技術は、2光子カルシウムイメージングの使用であった(Wallace and Kerr, 2010)。記録される過渡的なカルシウム動態が細胞の発火を密接に反映しているという証拠がある(Kerr and Denk, 2008)。しかし、この手法でさえ、細胞の発火の詳細な時間経過を追跡することは困難であり、より遅い時間スケールでの活動を総合的に見ることしかできない間接的な測定法である。Miller et al 2014)は、覚醒マウスを用いて、視覚刺激の後に繰り返し活動するニューロンのアンサンブルを非常に明確に示したが、カルシウムイメージングの時間分解能は、アンサンブル内でアトラクターが発達しているかどうかなど、ニューロンの詳細なダイナミクスを決定するのに十分な速さではないと主張した。もちろん、これらの手法は、意識的な経験を探求する上で重要な人間を対象とした研究には適していない。それにもかかわらず、アトラクター行動が発生しているという証拠はますます広まってきている。将来の大規模なコネクノミクスプロジェクトの主な目的は、大脳皮質ネットワークのアトラクター行動と、それによって生じる創発的な特性を探求することであろう(Alivisatos et al 2012)。
海馬の行動に関するアトラクターに基づく理論は以前から議論されており(Rolls, 1996; Tsodyks, 1999)、これらの理論を支持するいくつかの証拠が出てきている(Poucet and Save, 2005; Knieren and Zhang, 2012)。特にCA3は、異なる環境にさらされると、異なるパターンの活性を生成することができるようである(Guzowski et al 2004)。定点アトラクターの予測としては、入力がゆっくりと異なる種類のものから別の種類のものへと変化していくときに、ネットワークの応答が変化するのは、変化の途中で突然、ある活動パターンから別の活動パターンへと変化するようになるまでは、ネットワークの応答は変化しないというものがある。Wills et al 2005)は、環境刺激が段階的に円形から正方形のデザインに変更されたように、期待されるように、約半分のポイントでCA1場所の細胞の応答に突然の変化があったことを示した。また、モーフィングされた刺激を用いて覚醒サルのIT皮質の視覚処理を調べたところ、サルが行った分類決定を反映したニューロンが発見された(Akrami et al 2009)。マウスの聴覚野では、似たような音に反応するL2/3細胞のグループが発見され、音が変わるとこれらのグループ間の反応の遷移が急激になり、アトラクター的なダイナミクスを示唆している(Bathellier et al 2012)。覚醒したマウスの異なる音に対する行動は、グループの活動から予測することができ、その行動の関連性を強調している。Harris (2012)はこの研究についてのコメントで、細胞集合体の発火がいわゆる「バンプ」アトラクターに似ていると感じている。しかし、他の人は、環境刺激がこのように(Leutgeb et al 2005)で形態化されているように、より進歩的な変化を示している。Mattia et al 2013)は、別の手法(生体内試験での運動前皮質での多電極記録)を用いて、ステレオタイプのネットワーク活動を記録している。
このように、アトラクター状態の発達についての証拠は、一般的な発生であるという仮定が広まっているにもかかわらず、地に足がついていないのである。現在のところ、アンサンブル活動の中に協調的な発火サイクルがあるかどうかを判断するための測定技術は十分ではない。本当に必要なのは、意識的な報告の間にアトラクターが発生し、アトラクターが発生しないときには意識的な経験がないことを示す証拠である。これは覚醒している人間でのみ可能であるが、ECoG研究やLFPを用いた研究では、意識的な報告に関連したアトラクター活動の証拠は今のところ見当たらない。脳波やMEG活動を用いたアトラクター行動の間接的なシグネチャがあれば、もちろんそのような研究はより簡単になるだろう。
さらなるエビデンスを提供するためには何が必要か?
すべての高等動物がある程度の意識を経験している可能性が圧倒的に高いにもかかわらず、100%の確信が持てるのは人間だけである。したがって、最終的には、意識的経験を支えるヒトの活動を測定する必要がある。ここで提示された理論では、個々の細胞のネットワークの活動をサブミリ秒単位の分解能でモニターし、意識的な行為の際に細胞がどのように振る舞うか、また、それが無意識的な行為とどのように異なるかを見ることで、その証拠を得る必要がある。このような作業は、注意の配分、報告プロセスに関連する活動、先読みなどに関連する変動する活動を除去しなければならない。もちろん、集団モニタリングのための技術は急速に発展しており、2光子カルシウムイメージングが先駆的に使われている。現在のところ、この技術はネットワーク内の細胞の詳細な発火活動を調べるのに十分な速度ではないが、そう遠くない将来になることは間違いない。最初の例では、意識的な経験を持っていると強く疑われている高等哺乳類では、このような技術が有用に利用できる。このような動物が知覚を示さないのとは対照的に知覚を示したように、この記事で説明されているような局所的な活動をモニタリングすることから、強いポインタが得られるであろう。このような測定と並行して、脳波、MEG、またはECoGを用いてその活動のシグネチャを定義することができれば、それらのシグネチャを探すための人間の実験が可能になる。最終的には、それが記述されている局所的な活動を検出することができ、それが意識的な意識でのみ発生することを示すために、おそらく光イメージングの倫理的に許容可能な形で、人間で使用することができる技術を見つけることが必要になるが。
