
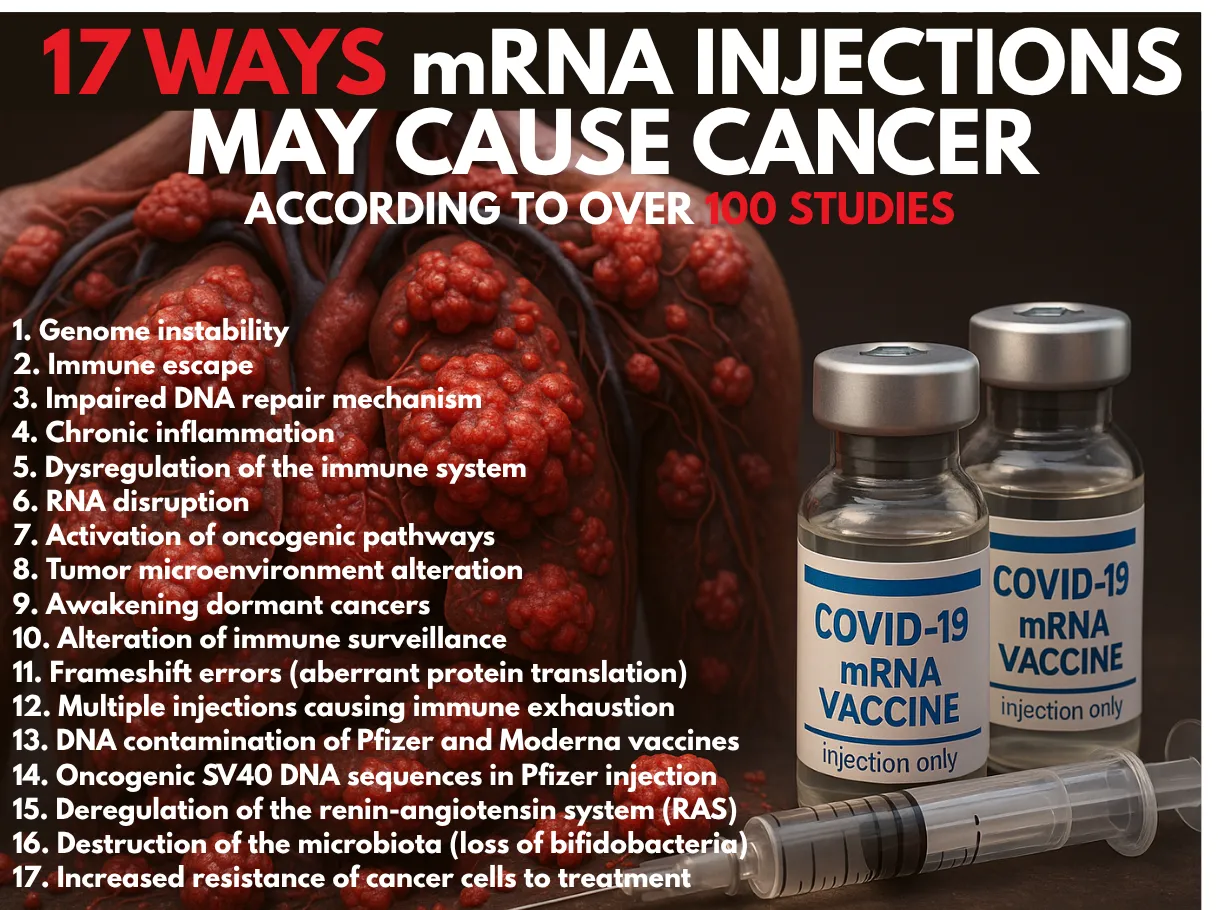
包括的な文献レビューにより、mRNAワクチンが17の異なる経路を通じてがんを引き起こす、促進する、または再活性化させる可能性が明らかになった。
https://www.thefocalpoints.com/p/17-ways-mrna-shots-may-cause-cancer
当記事の深層分析または超訳版はnote「Alzhacker」で公開中です。背景を深く理解したい方も、ポイントを気軽に確認したい方も、それぞれに適した形で情報を提供しています。
2025年6月25日
マチルド・デボードによる「COVID-19 mRNAワクチンは100件以上の研究によると17の異なる方法で癌を引き起こす可能性がある」というタイトルの包括的な文献レビューが、Le Point Critiqueに掲載された。100件以上の査読付き研究を引用し、注射が悪性プロセスを開始、加速、または再活性化させる可能性のある17の異なる生物学的メカニズムを概説している。
以下は、同論文で特定された17のメカニズムの要約である(これらの主張の根拠となる参考文献は、記事内に記載されている):
17のメカニズム(簡略版)
1. ゲノム不安定性
mRNAが逆転写されホストのDNAに組み込まれることで、がんを引き起こす突然変異を引き起こす可能性がある。
2. 免疫逃避
スパイクタンパク質は、p53やBRCA1などの腫瘍抑制遺伝子を結合し阻害し、がん細胞を免疫破壊から保護する。
3. DNA 修復メカニズムの障害
スパイクタンパク質は、必須のDNA 修復酵素の働きを妨害し、制御不能な突然変異のリスクを高める。
4. 慢性炎症
脂質ナノ粒子とスパイクタンパク質は、がんのよく知られた原因である長期にわたる炎症を引き起こす。
5. 免疫系の調節機能障害
T 細胞および I 型インターフェロンの抑制は、がんの監視機能を弱め、免疫回避を促進する。
6. RNAの破壊
コドン最適化はマイクロRNAネットワークを破壊し、細胞増殖調節とアポトーシスの安定性を損なう。
7. 発がん経路の活性化
スパイクタンパク質は間接的にMAPKとPI3K/mTORシグナル伝達経路を活性化し、腫瘍の成長と転移を促進する。
8. 腫瘍微小環境の変容
脂質ナノ粒子腫瘍に蓄積し、透過性を高め、がん転移を加速する可能性がある。
9. 休眠がん細胞の活性化
ワクチン接種後の炎症と免疫障害は、以前に寛解していた患者で再発を引き起こす可能性がある。
10. 免疫監視の異常
改変されたmRNAはトール様受容体を阻害し、腫瘍細胞を免疫系から「見えない」状態にする。
11. フレームシフトエラー
合成mRNAは時に意図しない異常なタンパク質を産生し、腫瘍形成リスクに寄与する。
12. 複数回接種
反復投与は免疫系を消耗し、IgG4へのクラススイッチを促進し、腫瘍に対する耐性を高める可能性がある。
13. DNA汚染
ワクチンバイアルに残存するプラスミドDNAは複製能を有し、ホストゲノムに組み込まれる可能性がある。
14. 発がん性SV40 DNA配列
ファイザーのバイアルに含まれるSV40プロモーター配列はゲノム挿入を促進する可能性があります—この同じ要素は実験動物で腫瘍を誘導するために使用されている。
15. レニン・アンジオテンシン系(RAS)の調節障害
スパイクによるAT1Rの活性化は、酸化ストレスと制御不能な細胞増殖を促進する。
16. マイクロバイオータの破壊
注射はビフィドバクテリウムを減少させ、免疫バランスを弱め、抗がん反応を妨げる。
17. 治療抵抗性の増加
スパイクへの曝露は化学療法中の癌細胞の生存期間を延長し、治療抵抗性を引き起こす可能性がある。
これらのデータは、COVID-19 mRNAワクチン接種キャンペーン開始以来、米国で記録された110,750件の過剰癌死亡を説明するのに役立つ。CDCの公式データセットの分析によると、過剰癌死亡率は単に持続しているだけでなく、2025年にさらに加速していることが明らかになっている:
疫学者、マッカロー財団事務局長
www.mcculloughfnd.org
文献レビュー(17のメカニズムの詳細)
マチルド・デボルド | 2025年6月19日
https://lepointcritique.fr/2025/06/19/vaccins-arnm-anti-covid-peuvent-induire-cancer-de-17-manieres-distinctes-selon-plus-de-100-etudes/#_edn69
100件を超える研究によると、COVID-19用mRNAワクチンは17の異なる方法でがんを引き起こす可能性がある
科学界の一部は、4年以上前からCOVIDに対するmRNAワクチン接種に関連するがんリスクについて警鐘を鳴らしてきた。現在の文献データは、これらのワクチンが17種類の異なる方法でがんを引き起こす、促進する、または再発させる可能性があると示唆している。
COVIDに対する大量ワクチン接種が、多くの科学者(その一部は2021年5月に予言していた)が主張するように、癌の症例急増の原因となっているのか?フランスの腫瘍学者集団は2年前に論説を発表し、この仮説を断固として否定している。「現在まで、COVID-19ワクチン接種後、あるいは他のワクチン接種後に、がんの発生率の増加や進行の急速な進行との関連について、警告する報告は発表されていない」と。彼らは現在、特に若者に、急速に進行するがんの津波に直面しており、その原因について合理的な説明は見つからないと主張している。
膵臓がんの急激な増加が見られるが、その理由はまったくわからない。何かが起こったのか?わからない。世界中の、すべての腫瘍学者がこの疑問を抱いている。[…]がんの理解を可能にするシステムが機能していない。
カヤット教授の主張が首尾一貫しているならば、彼は、ワクチン接種がこの癌の爆発的な増加の原因である可能性を理論的には排除できない。なぜなら、この癌の増加は (1) 彼のこれまでの発言から判断すると、ごく最近のことである(2) これは世界全体に影響を及ぼしている(特に、社会生活を維持するために接種を強制された人々や、ワクチン接種を積極的に推進した人々(インフルエンサーなど))、(3) これまでにない論理に基づいているように見える。これは、人間に対して初めて使用され、その組成の一部しか知られておらず、大規模な展開前に癌への影響が評価されていない物質の場合と同じだ[1]。
疫学者のニコラス・フッシャー氏は、昨年 3 月、COVID 対策のmRNA ワクチンが癌を引き起こす 10の方法をリストアップした。このリストは、2023 年 12 月に雑誌Cureusに掲載された研究[2]に基づいて作成されたものだが、100 件以上の研究(すべては網羅していない)を基に、現在では17 項目に拡大できると思う。
1. ゲノムの不安定化
ワクチン用RNAがワクチン接種を受けた人のゲノムに組み込まれるリスクは、2021年に一連の研究[3]、[4]、[5]によって確認された。DNAの統合によって誘発される挿入性突然変異は、フレームシフト(frameshift)と呼ばれる突然変異を引き起こし、がんを引き起こす異常なタンパク質の産生を誘発する。
欧州医薬品庁は、ワクチン用mRNAは細胞核に侵入することはできないと依然として主張している。この統合には、人間には存在しない酵素(逆転写酵素)が必要だからだとしている。しかし、何の証拠もないこの主張は、2021年6月に否定された。この現象は、2023年7月にマウスで観察され、1回のmRNA注射で遺伝子変異が誘発された[6]。さらに最近では、ワクチン接種を受けた患者の腫瘍からワクチン用スパイクタンパク質が検出され[7]、これがゲノムに組み込まれる可能性があることが示唆された。このような組み込みの最初の懸念される影響は、がんの発症だ。
この仮説は、4月中旬、生体分子研究研究所(Neo7Bioscience)の科学者と北テキサス大学の研究者たちによって再浮上した[8]。彼らが収集した分子データは、ワクチン由来のRNAが宿主のゲノムに逆転写され、遺伝子の調節を恒久的に変化させる可能性があることを示唆している。また、発がん性および免疫系の崩壊の兆候も示している。
2. 免疫回避
スパイクタンパク質(S2)は、複数の腫瘍抑制遺伝子(p53、BRCA1/2、RB1)[9],[10],[11]に結合し、がん細胞が免疫系による検出と破壊を回避することを可能にする。疫学者のニコラス・フルッシャーは、これを「発癌の逆転」と呼んでいる。
スパイクタンパク質が「ゲノムの番人」とも呼ばれるp53 タンパク質と干渉することを示した最初の研究は、2021年10月にJiang らによって発表された[12]。この研究は、2022年5月にアンソニー・ファウチ氏率いるNIHの指示により撤回された。この撤回に関するメール交換の公開を求める請求が情報公開法に基づいて提出されたが、NIH は依然として 490 ページにわたる通信文書の公開を拒否している。これらの結果は、2024年にZhangとEl Deiry[13]によってin vitroで、1 か月後にはin vivo[14]で確認された。
3. 損傷したDNAの修復メカニズム
ワクチンに含まれるスパイクタンパク質は、ゲノムに損傷を与え、DNA修復システムを阻害する(Jiang、Zhang、El Deiry)。。このメカニズムは、通常、生物が攻撃を受けた場合に、がんの発生を促進する突然変異を防ぎ、がん遺伝子や腫瘍抑制遺伝子に影響を与えるエラーを修復するために働く。この損傷は、免疫不全を引き起こし、「がんへの直接的な道[15]」となる。
2016年にModerna社のCEO、ステファン・バンセル氏によって特許取得されたスパイクタンパク質の戦略的配列は、その改変によりDNA修復の欠陥を引き起こす遺伝子(MSH3)を[16]標的とすることができる。スパイクタンパク質がこのメカニズムを阻害する経路は、昨年4月に発表されたBaşaran et al.[18]の論文に記載されている。
4. 慢性炎症
ワクチン用mRNAの輸送に使用される脂質ナノ粒子[19]、[20]は、炎症性タンパク質の大量分泌[21]、[22]、[23]、[24](サイトカインストーム)を引き起こし、がん幹細胞の出現につながる。これらの細胞は、スパイクタンパク質の広範な生体内分布[26]、[27]により、すべての臓器(血液幹細胞[25]を含む)で増殖する可能性がある。その病原性は、3つの文献[28]、[29]、[30] および 320 件以上の研究で詳しく説明されている。この炎症は、T 細胞の消耗を引き起こし、その結果、T 細胞は癌細胞を排除できなくなる。
AI Grok は、注射が急性炎症を引き起こすことを確認しているが、この炎症は数日で治まり(Bergamaschi、Ogata)、他のワクチンと類似していると述べている。また、慢性炎症には持続的な刺激が必要であり、「ワクチン由来のスパイクタンパク質の産生は時間的に限定されており (mRNAは数日で分解され、スパイクは数週間で分解される)ため、慢性炎症は起こりにくい」と述べている。この主張は、一連の研究[31]、特にスパイクタンパク質が注射後709日まで血液中に検出された4つの最近の研究によって反論されている[32](245 日[33]または 12 ヶ月[34]、他の研究では)で、日本の女性患者の組織および臓器、特に脳では 17 ヶ月[35]まで検出されている。最初の接種から 4 年以上が経過した現在、体がその産生を停止するかどうかは実際には不明だ。
5. 免疫系の調節機能障害
mRNAワクチン接種は、T細胞(リンパ球減少症)[36]およびインターフェロンI型[37]の反応を抑制し、がんの監視と増殖に重要な役割を果たしている。これらの変化は、自然免疫の障害[38]、[39]、[40]、[41]および適応免疫の再プログラム[42]、[43]を引き起こす。中枢神経系における免疫系の調節異常も報告されている[44]。
COVID-19ワクチン接種において、この抑制は適切なスパイクタンパク質の合成と免疫応答の低減を保証する。メラノーマモデルにおいて、mRNAワクチンに100%のN1-メチル-偽ウリジン(m1′)を追加すると、がん成長と転移が促進されたことが示されている。一方、mRNAを改変していないワクチンでは逆の結果は観察されなかった。これにより、COVID-19 mRNAワクチンががん発症を助長する可能性が示唆されている。
Rubio-Casillas et al. Review: N1-methyl-pseudouridine (m1Ψ): Friend or foe of cancer? https://doi.org/10.1016/j.ijbiomac.2024.131427.
Grok は、BioNTechのCEO であるUgur Sahinによる2020 年の研究[45]を引用し、mRNA ワクチンは、ワクチン接種後数日から検出される、持続的なCD4+ および CD8+ T 細胞の強力な反応を誘発し、全身的な持続的な免疫抑制という説と矛盾すると主張している。それどころか、ファイザー社の臨床データでは、ワクチン接種後 6~8 日間に、参加者の45%~46% で T リンパ球の減少が認められ[46]、その減少は時間とともに悪化する事が現在分かっている。
6. RNAの障害
ワクチン用 mRNA は、その寿命と産生量を増加させる目的で改変された mRNAだ。ファイザーとモデルナが採用している技術(コドン最適化)は、細胞の増殖と死、特に癌細胞の増殖と死に重要な役割を果たすマイクロ RNAを障害する[47]、[48]。2021年の研究[49]では、ワクチンに含まれるスパイクタンパク質は、マイクロRNAを含む小胞(エキソソーム)によって運ばれ、インターフェロンの働きを阻害して、細胞の増殖や腫瘍の監視などの細胞プロセスを乱し、自然免疫を阻害することが示されている。
7. 癌遺伝子経路の活性化
スパイクタンパク質は、腫瘍の成長、細胞の増殖および生存に重要な役割を果たす複数の経路(MAPK、PI3K/AKT/mTOR[50]、[51],[52])を活性化し、免疫、炎症、腫瘍の成長、転移の進行、免疫療法に対する耐性に関与する炎症マーカーであるインターロイキン 6(IL-6)のレベルを上昇させる疑いがある。IL-6の慢性的な上昇は、特定の状況において癌を促進する可能性のある炎症と関連している。
Grok は、これらの障害と癌との関連を正式に立証した研究はないと改めて指摘しているが、最近の研究[53]では、ファイザーの2 回目または 3 回目の接種後数週間で白血病を発症した患者において、PI3K/mTOR 経路などの特定の癌遺伝子経路の活性化の代謝的証拠が発見されている。
8. 腫瘍の微小環境
脂質ナノ粒子(NPL)は、Enhanced Permeability and Retention(EPR)効果により、腫瘍の組織に蓄積する。この効果は、腫瘍の血管の透過性が高まり、ナノ粒子が腫瘍組織に長期間留まるという特徴がある。これにより、NPL はがん細胞の拡散を加速[54][55] これは、病理学者や腫瘍学者によって記述され、マウスを用いた研究で観察された「ターボがん」現象を説明しているかもしれない[56]。このような病原性プロセスの加速は、スパイクタンパク質によって誘発される他の疾患にも当てはまるのだろうか?スウェーデンの研究者は2023年に、スパイクタンパク質がアルツハイマー病を引き起こすだけでなく、その潜伏期間を80%短縮[57]し、これまでに見られなかった「ターボアルツハイマー病」を引き起こすことを実証した。
9. 休眠中の癌の覚醒
COVID-19に関連する炎症やワクチン接種によって腫瘍の微小環境に生じる変化は、癌の覚醒や転移の再発に影響を与える可能性がある[58]。
長年がんが寛解していた患者が、COVID-19ワクチンブースター接種後、突然、進行が速く爆発的ながんを再発するケースが報告されている。これらの症例では、ブースター接種後に腫瘍の急速な増殖が観察されている。これらの「ターボがん」は、数年安定していた患者を含む、予想以上に早期に発症し、より悪性度が高い特徴を示している。公衆衛生当局は、この相関関係を認めることに消極的だ。この現象は、mRNAワクチンが投与された世界中のあらゆる場所で発生している。
イアン・ブライトホープ教授。大討論:ポート・ヘドランド対首相。2024年11月29日
SARS-CoV-2のスパイクタンパク質が複数の細胞を融合する能力[59],[60],[61] は、COVID-19の合併症の連鎖、その中には癌も含まれる、を説明できるかもしれない。この融合によって生じるシンシチウムの形成は、特に既存の損傷がある場合、がんの発生や進行、さらには転移や寛解後のがんの再発にも寄与する可能性がある、とイェール大学元教授のユリ・ラゼブニ氏は述べている[62]。
なお、COVID-19に対する有効性が現在 100 件以上の研究で確認されているイベルメクチンには、多くの抗腫瘍作用がある[63](腫瘍幹細胞の増殖、転移、血管新生活性の抑制、がん細胞のプログラム細胞死の促進、多剤耐性の逆転)があり、スパイクタンパク質による細胞融合で誘発される合胞体の形成を阻害する能力も持っている[64]。2006年にノーベル賞を受賞したイベルメクチンは 2006年にノーベル賞を受賞したこの物質は、子供や妊娠中の女性を含む極めて高い安全性を示しており、WHOにより必須の分子とされている。なぜ、mRNA注射の有効性、安全性、発がん性について何も知られていないのに、イベルメクチンは承認されなかったのだろうか?この質問は、いずれ早急に提起される必要がある。
10. 免疫監視の障害
改変mRNAは、免疫系の第一線受容体(Toll様受容体、TLR)の活性化を阻害することで、腫瘍細胞を「見えなく」する。。
KarikóとWeissman、ファイザーのCOVIDワクチンを開発した2人の研究者は、2005年に[65]、m1Ψの添加によるRNAの合成改変 (この発見で彼らはノーベル医学賞を受賞した)を合成的に改変することで、このシールドを部分的に除去し、天然のRNAが一次樹状細胞を活性化させる能力を阻害すると説明している。これらの細胞の機能の一つは、病原体、特にがん細胞を認識または「信号伝達」し、標的を絞った免疫応答を誘導することだ。
この結果は、2015年[66]および2016年[67]に確認された。2016年の研究では、合成mRNAの使用は天然のRNAよりも効果的ではなく、むしろ毒性(サイトカインの上昇、好中球増加)を増加させることも示されている。これは、血液と脾臓の骨髄系細胞を活性化することで、がん化プロセスを反映している可能性がある。
2021年の研究[68]では、Toll型受容体が両刃の剣として作用し、TLR反応が調節不全に陥ると、本来予防すべき「 病態の悪化」を引き起こす可能性があることが確認されている。
11. フレームシフト(frameshift)
ファイザーとモデナのワクチンに含まれる改変mRNAは、翻訳されると異常な免疫反応を引き起こす。3分の1のケースでは、ワクチンmRNAは、プログラムされたスパイクタンパク質とは異なる「意味不明」または未知のタンパク質を産生する。この研究[69] は 2024 年 1 月に発表された。著者らは、これらの翻訳エラーが次世代のCOVID-19 ワクチンで解決されない場合、重大な安全性の問題となると認めている。しかし、この発見は技術の安全性を損なうものではないと彼らは考えている。別の研究者チーム[70] は、昨年 6 月、このmRNA プラットフォームの重大な欠陥について、はるかに厳しい診断を下した。
「改変された mRNA […] は、その持続性、潜在的な永続性、および免疫刺激性により、臨床では使用できない。」 […] SARS-CoV-2のスパイクタンパク質をコードするmRNAの持続性は、この病原性タンパク質への危険なほど長期間の無制限の曝露をもたらすため、人間への継続的な使用については再評価が必要だ。
12. 複数回接種
合成 mRNA およびワクチンスパイクへの反復曝露は、免疫系の疲弊を引き起こす[71]。この免疫抑制は、おそらくコドン最適化と抗体依存性増強(ADE)メカニズム[72]、[73](臨床試験から疑われている)によって説明でき、抗体のクラス変化(IgG4)[74]、[75],[76] であり、現在では一連の研究によって、注射の有効性の欠如が証明されている。ピーター・マッカロー財団は、現在までに7 件を[77],[78],[79],[80],[81],[82],[83]。この壊滅的な免疫反応の変化は、異種ワクチンやDNA ワクチン(Irrgang)の接種後には見られないもので、免疫寛容(病原体が病原体として認識されなくなる)を引き起こし、再感染[84]、[85] および癌の発症[86]、[87]、[88]、[89]。
13. ファイザーとモデルナのワクチンにおけるDNA汚染
ファイザーとモデルナのワクチンには、不正なプラスミドDNA[90]、[91]、[92]、[93]が含まれており、その形態(二重らせん構造)により「複製能を有する」、つまり理論的にはゲノムに組み込まれ、ワクチン接種者に癌を誘発する可能性がある。私たちはこの発見について数多くの記事を書いてきたが、現在までに世界 10 組の研究者チーム[94]によって確認されており、最新のものは、スロバキア政府から委託を受けた分子遺伝学者(ソニャ・ペコヴァ博士)によるものだ。[95]が、スロバキア政府から委託を受けて発表したものである。報告されている量は、欧州医薬品庁が定めた上限の500 倍にも達する驚異的なもので、ゲノムへの組み込みが自発的に起こる可能性があり、がんのリスクを最大限に高めることを意味する。
14. ファイザーのワクチンに含まれる発がん性 SV40のDNA 配列
細胞核を「ハッキング」するために遺伝子工学で使用されるSV40の戦略的な配列を追加することで、mRNAがゲノムに組み込まれる能力が10 倍に高まる[96]。
FDAによって使用が禁止されていたが、最終的にファイザーによって承認されたが、同社は健康上のリスクはないと主張している。その発がん性はすでに多くの文献で報告されているが、昨年10月、New England Journal of Medicine[97]に掲載された研究で確認された。また、ファイザーとモデルナのワクチン接種を受けた被験者を対象とした最近の研究[98]では、腫瘍抗原がファイザーのワクチン接種を受けた被験者の血液から独占的に検出されており、このワクチンががんを引き起こす可能性も示唆されている。
この発見の発端となったマッケルナン博士によると、ファイザーのワクチン(成人用、小児用、単価、二価)はすべて、パンデミックによる産業上の制約に対応するため、注射剤の認可後に実施された製造方法の変更[99]が原因とされるこの不正に関与しているとのことである。アンガス・ダルグレイシュ教授、世界でも最も著名な腫瘍学者の一人である同教授は、この文脈で、SV40 は、化学療法を試験する際に、がん研究者がマウスに注射してがんを誘発する物質であることを指摘した。現在、抗がん剤市場でほぼ独占的な地位を占めるファイザー社は、この事実を知らなかったのだろうか?いいえ、ワクチン需要の急落にもかかわらず、同社は製剤を変更しなかった。
15. レニン・アンジオテンシン系(SRA)の調節機能障害
ワクチンに含まれるスパイクタンパク質は、細胞の増殖などを制御するレニン・アンジオテンシン系の重要な受容体(AT1R)を過剰に活性化する。この過剰活性化は血管新生を促進し、その結果、腫瘍の増殖を引き起こし、細胞に有害な酸化ストレスを引き起こす。ジャン=マルク・サバティエ博士[100] は、2020年3月に、この生理的調節の乱れが、先天性免疫と獲得免疫のバランスを崩し、多くの癌を引き起こす危険性があると警告していた。
16. マイクロバイオームの破壊
mRNA「ワクチン」は、マイクロバイオーム(腸内細菌叢)に存在するビフィズス菌を破壊する。ビフィズス菌は、がんの調節や抗がん療法への反応に重要な役割を果たしている。サビーヌ・ハザン博士が発表した画期的な研究[101]は、2022年に、COVIDに対するmRNAワクチンが腸内微生物叢に存在するビフィズス菌を激減させることを示した。この減少は、侵襲性がん患者で観察された。ワクチン接種がマイクロバイオームに及ぼす有害な影響は、ファイザーワクチンを接種した患者の大腸腫瘍の生検からスパイクタンパク質が検出されたことで、現在確認されているようだ。
17. 治療に対する耐性の増加
ウイルス性およびワクチン由来の可能性のあるスパイクタンパク質は、化学療法への曝露後にがん細胞の生存を延長する。この結果は、2024年にS. ZhangとWS El-Deiryによって明らかにされた。証拠はウイルスのスパイクタンパク質に限定されているが、著者らは、p53 遺伝子の阻害やDNA損傷に対する反応の障害と密接に関連しているこの免疫応答の障害は、追加接種として投与される反復注射や、生産されるスパイクタンパク質の天文学的な量によって促進される可能性があると考えている。
この悲惨な状況[102]には、注射剤に隠れた遺伝子が存在している可能性も加わる。その遺伝子が健康に与える影響は、誰にも予測できない。残念ながら、注射の安全性を示す証拠は一切ない。なぜなら、その発がん性はどの試験でも評価されておらず、私たちの知る限り、注射ががんを引き起こしたり、再発させたり、進行を早めることはないことを示す研究はないからだ。
2021年にオーストラリアで開始された大規模な臨床試験は、この疑問に答えることを目的としていた。しかし、この試験はオーストラリア当局によって説明もなしに突然中止され、この目的のために収集された何百万もの生体組織サンプルが違法に破壊されようとしている。さらに、非常に不穏な事実として、複数の国から高毒性のファイザー製ワクチンが存在することが報告されており、この研究所が3つの異なる毒性レベルの製品を開発した可能性も指摘されている。がんの集団発生が、最近、米国とオーストラリアの複数の病院で報告されている。しかし、感染した看護師の一人に投与されたワクチンは、まさにこの高リスクのバッチから製造されたもので、最も多くのDNAが検出されたバッチと同じだった。
もはや誰もその現実を否定できない現在の世界的な流行は、その惨憺たる失敗にもかかわらず、業界が巨額の投資を投じた技術を試験するために事前に計画されたものなのか?[103]、そして、今日では後戻りが不可能なほど巨額の投資を行っているこの技術をテストするために、あらかじめ予測されていたものなのか?これは、職を失わないためにワクチン接種を受け、数ヶ月後に癌で亡くなった看護師の夫、その夫は、計画的な毒殺で告訴している。
参考文献
[1] Acevedo-Whitehouse K, Bruno R. Potential health risks of mRNA-based vaccine therapy: A hypothesis. Med Hypotheses. 2023 Feb;171:111015. https://doi.org/10.1016/j.mehy.2023.111015.
[2] Valdes Angues R, Perea Bustos Y. SARS-CoV-2 Vaccination and the multi-hit hypothesis of oncogenesis Cureus. 2023 Dec 17;15(12):e50703. https://doi.org/10.7759/cureus.50703.
[3] Aldén M, Olofsson Falla F, Yang D, Barghouth M, Luan C, Rasmussen M, et al. Intracellular reverse transcription of Pfizer BioNTech COVID-19 mRNA vaccine BNT162b2 in vitro in human liver cell line. Curr Issues Mol Biol. 2022 Feb 25;44(3):1115-26. https://doi.org/10.3390/cimb44030073.
[4] Chandramouly G, Zhao J, McDevit S, Rusanov T, Hoang T, Borisonnik N, et al. Polθ reverse transcribes RNA and promotes RNA-templated DNA repair. Sci Adv. 2021 Jun 11;7(24):eabf1771. https://doi.org/10.1126/sciadv.abf1771.
[5] Zhang L, Richards A, Barrasa MI, Hughes SH, Young RA, Jaenisch R.. Reverse-transcribed SARS-CoV-2 RNA can integrate into the genome of cultured human cells and can be expressed in patient-derived tissues. PNAS. 2021 May 25;118(21):e2105968118. https://doi.org/10.1073/pnas.2105968118.
[6] Breda L, Papp TE, Triebwasser MP, Yadegari A, Fedorky MT, Tanaka N, et al. In vivo hematopoietic stem cell modification by mRNA delivery. Science. 2023 Jul 28;381(6656):436-443. https://www.doi.org/10.1126/science.ade6967.
[7] Le Dr McKernan a fait cette découverte en analysant la biopsie du cancer du côlon d’une personne ayant reçu quatre injections d’ARNm de Pfizer. Le député et ancien ministre japonais Kazuhiro Haraguchi a déclaré publiquement fin mai que de la protéine Spike vaccinale avait également été retrouvée dans les cellules cancéreuses de son lymphome malin.
[8] https://twitter.com/tatiann69922625/status/1931708697379480010. Cette découverte a fait l’objet d’une discussion avec l’épidémiologiste Nicolas Hulscher : https://www.thefocalpoints.com/p/breaking-reverse-transcription-cancer.
[9] Singh N, Singh AB. S2 Subunit of SARS-nCoV-2 interacts with tumor suppressor protein p53 and BRCA: an in silico study. Transl Oncol. 2020 Oct;13(10):100814. https://doi.org/10.1016/j.tranon.2020.100814. Les résultats de cette étude ont été publiés le 30 juin 2020.
[10] Chen X, Zhang T, Su W, Dou Z, Zhao D, Jin X, et al. Mutant p53 in cancer: from molecular mechanism to therapeutic modulation. Cell Death Dis. 2022 Nov 18;13(11):974. https://doi.org/10.1038/s41419-022-05408-1.
[11] Selon Valdes Angues et al., « Les cancers associés aux mutations TP53 incluent le cancer du sein, les sarcomes des tissus osseux et mous, les tumeurs cérébrales et les carcinomes corticosurrénaliens. D’autres cancers moins fréquents incluent la leucémie, le cancer de l’estomac et le cancer colorectal. Les cancers associés à une altération de l’activité de BRCA1 comprennent le cancer du sein, de l’utérus et de l’ovaire chez les femmes, le cancer de la prostate et du sein chez les hommes, et une augmentation modeste du cancer du pancréas chez les hommes et les femmes. Les cancers les plus fréquemment rapportés avec des mutations BRCA2 incluent le pancréas, la prostate chez les hommes et le mélanome ».
[12] Jiang H, Mei YF. SARS-CoV-2 spike impairs DNA damage repair and inhibits v(d)j recombination in vitro. Viruses. 2021;13(10):2056. https://doi.org/10.3390/v13102056. Lien vers l’étude en français : https://www-mdpi-com.translate.goog/1999-4915/13/10/2056?_x_tr_sl=auto&_x_tr_tl=fr&_x_tr_hl=fr&_x_tr_pto=wapp.
[13]Zhang S, El-Deiry WS. SARS-CoV-2 Spike protein inhibits tumor suppressor p53 and creates a vulnerability to cancer. bioRxiv (preprint). 2024 Apr 15. https://doi.org/10.1101/2024.04.12.589252.
[14] Zhang S, El-Deiry WS. Transfected SARS-CoV-2 spike DNA for mammalian cell expression inhibits p53 activation of p21(WAF1), TRAIL Death Receptor DR5 and MDM2 proteins in cancer cells and increases cancer cell viability after chemotherapy exposure. Oncotarget. 2024 May 3;15:275-284. https://doi.org/10.18632/oncotarget.28582.
[15] Seneff S, Nigh G, Kyriakopoulos AM, McCullough PA. Innate immune suppression by SARS-CoV-2 mRNA vaccinations: The role of G-quadruplexes, exosomes, and MicroRNAs. Food Chem Toxicol. 2022 Apr 15;164:113008. https://doi.org/10.1016/j.fct.2022.113008.
[16] Ambati BK, Varshney A, Lundstrom K, Palú G, Uhal BD, Uversky VN, et al. MSH3 homology and potential recombination link to SARS-CoV-2 furin cleavage site. Front Virol. 2022 Feb;2:834808. https://doi.org/10.3389/fviro.2022.834808.
[17] http://genatlas.medecine.univ-paris5.fr/fiche.php?symbol=MSH3.
[18] Başaran N, Szewczyk-Roszczenko O, Roszczenko P, Vassetzky Y, Sjakste N. Genotoxic risks in patients with COVID-19. Infect Genet Evol. 2025 Apr;129:105728. https://doi.org/10.1016/j.meegid.2025.105728.
[19] Kedmi R, Ben-Arie N, Peer D. The systemic toxicity of positively charged lipid nanoparticles and the role of Toll-like receptor 4 in immune activation. Biomaterials. 2010 Sep;31(26):6867-75. https://doi.org/10.1016/j.biomaterials.2010.05.027.
[20] Turni C, Lefringhausen A. Covid-19 vaccines: An Australian review. J. Clin. Exp. Immunol. 2022 Sep 21;7(3):491-508. https://blog.fdik.org/2023-03/covid19-vaccinesan-australian-review.pdf.
[21] Ndeupen S, Qin Z, Jacobsen S, Bouteau A, Estanbouli H, Igyártó BZ.The mRNA-LNP platform’s lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. iScience. 2021 Dec 17;24(12):103479. https://doi.org/10.1016/j.isci.2021.103479.
[22] Alameh MG, Tombácz I, Bettini E, Lederer K, Sittplangkoon C, Wilmore JR, et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity. 2021 Dec 14;54(12):2877-2892.e7. https://doi.org/10.1016/j.immuni.2021.11.001.
[23] Theoharides TC. Potential association of mast cell activation by the SARS-CoV-2 spike protein with cancer progression. Med Hypoth. 2022;160:110774. https://doi.org/10.1016/j.mehy.2022.110774.
[24] Sagala G. Apparent cytotoxicity and intrinsic cytotoxicity of lipid nanomaterials contained in a COVID-19 mRNA Vaccine. IJVTPR. 2023 Oct 16;3(1):957-72. https://doi.org/10.56098/ijvtpr.v3i1.84.
[25] Estep BK, Kuhlmann CJ, Osuka S, Suryavanshi GW, Nagaoka-Kamata Y, Samuel CN, et al. Skewed fate and hematopoiesis of CD34+ HSPCs in umbilical cord blood amid the COVID-19 pandemic. iScience. 2022 Dec 22;25(12):105544. https://doi.org/10.1016/j.isci.2022.105544.
[26] Jahankhani K, Ahangari F, Adcock IM, Mortaz E. Possible cancer-causing capacity of COVID-19: Is SARS-CoV-2 an oncogenic agent? Biochimie. 2023 May 23;S0300-9084(23)00136-0. https://doi.org/10.1016/j.biochi.2023.05.014.
[27] Australian Therapeutic Goods Administration (TGA). Nonclinical evaluation report: BNT162b2 [mRNA] COVID-19 vaccine (COMIRNATYTM). 2021 Jan. p. 45. https://www.tga.gov.au/sites/default/files/foi-2389-06.pdf#page=45.
[28] Trougakos IP, Terpos E, Alexopoulos H, Politou M, Paraskevis D, et al. COVID-19 mRNA vaccine-induced adverse effects: unwinding the unknowns. Trends Mol Med. 2022 Oct;28(10):800-802. https://doi.org/10.1016/j.molmed.2022.07.008.
[29] Parry PI, Lefringhausen A, Turni C, Neil CJ, Cosford R, Hudson NJ, Gillespie J. ‘Spikeopathy’: COVID-19 spike protein is pathogenic, from both virus and vaccine mRNA. Biomedicines. 2023 Aug 17;11(8):2287. https://doi.org/10.3390/biomedicines11082287.
[30] Posa A. Spike protein-related proteinopathies: A focus on the neurological side of spikeopathies. Ann Anat. 2025 Apr 18;260:152662. https://doi.org/10.1016/j.aanat.2025.152662.
[31] Petrosin S, Matende N. Elimination/Neutralization of COVID-19 Vaccine-Produced Spike Protein: Scoping Review. Mathews J Nutr Diet. 2024;7(2):1-23. https://doi.org/10.30654/MJND.10034.
[32] Bhattacharjee B, Lu P, Monteiro VS, Tabachnikova A, Wang K, Hooper WB, et al. Immunological and antigenic signatures associated with chronic illnesses after1 COVID-19 vaccination. MedRxiv (preprint). 2025 Feb 25. https://doi.org/10.1101/2025.02.18.25322379.
[33] Patterson BK, Yogendra R, Francisco EB, Guevara-Coto J, Long E, Pise A, et al. Detection of S1 spike protein in CD16+ monocytes up to 245 days in SARS-CoV-2-negative post-COVID-19 vaccine syndrome (PCVS) individuals. Hum Vaccin Immunother. 2025 Dec;21(1):2494934. https://doi.org/10.1080/21645515.2025.2494934.
[34] Alghamdi A, Hussain SD, Wani K, Sabico S, Alnaami AM, Amer OE, et al. Altered circulating cytokine profile among mRNA-vaccinated young adults: A year-long follow-up study. Immun Inflamm Dis. 2025 Apr;13(4):e70194. https://doi.org/10.1002/iid3.70194.
[35] Ota N, Itani M, Aoki T, Sakurai A, Fujisawa T, Okada Y, et al. Expression of SARS-CoV-2 spike protein in cerebral Arteries: Implications for hemorrhagic stroke post-mRNA vaccination. J Clin Neurosci. 2025 Jun;136:111223. https://doi.org/10.1016/j.jocn.2025.111223.
[36] Valdes Angues R, Perea Bustos Y. SARS-CoV-2 Vaccination and the multi-hit hypothesis of oncogenesis Cureus. 2023 Dec 17;15(12):e50703. https://doi.org/10.7759/cureus.50703.
[37] Rubio-Casillas A, Cowley D, Raszek M, Uversky VN, Redwan EM. Review: N1-methyl-pseudouridine (m1Ψ): Friend or foe of cancer? Int J Biol Macromol. 2024 May;267(Pt 1):131427. https://doi.org/10.1016/j.ijbiomac.2024.131427.
[38] Ivanova EN, Shwetar J, Devlin JC, Buus TB, Gray-Gaillard S, Koide A, et al. mRNA COVID-19 vaccine elicits potent adaptive immune response without the acute inflammation of SARS-CoV-2 infection. iScience. 2023 Nov 24;26(12):108572. https://doi.org/10.1016/j.isci.2023.108572.
[39] Seneff S, Nigh G, Kyriakopoulos AM, McCullough PA. Innate immune suppression by SARS-CoV-2 mRNA vaccinations: The role of G-quadruplexes, exosomes, and MicroRNAs. Food Chem Toxicol. 2022 Apr 15;164:113008. https://doi.org/10.1016/j.fct.2022.113008.
[40] Liu J, Wang J, Xu J, Xia H, Wang Y, Zhang C, et al. Comprehensive investigations revealed consistent pathophysiological alterations after vaccination with COVID-19 vaccines. Cell Discov. 2021 Oct 26;7(1):99. https://doi.org/10.1038/s41421-021-00329-3.
[41] Ning W, Xu W, Cong X, Fan H, Gilkeson G, Wu X, et al. COVID-19 mRNA vaccine BNT162b2 induces autoantibodies against type I interferons in a healthy woman. J Autoimmun. 2022 Oct;132:102896. https://doi.org/10.1016/j.jaut.2022.102896.
[42] Qin Z, Bouteau A, Herbst C, Igyártó BZ. Pre-exposure to mRNA-LNP inhibits adaptive immune responses and alters innate immune fitness in an inheritable fashion. PLoS Pathog. 2022 Sep 2;18(9):e1010830. https://doi.org/10.1371/journal.ppat.1010830.
[43] Föhse FK, Geckin B, Zoodsma M, Kilic G, Liu Z, Röring RJ, et al. The BNT162b2 mRNA vaccine against SARS-CoV-2 reprograms both adaptive and innate immune responses. Clin Immunol. 2023 Oct;255:109762. https://doi.org/10.1016/j.clim.2023.109762.
[44] Abramczyk H, Brozek-Pluska B, Beton K. Decoding COVID-19 mRNA vaccine immunometabolism in central nervous system: human brain normal glial and glioma cells by Raman imaging. Biovrix. 2022 Mar 2. https://doi.org/10.1101/2022.03.02.482639.
[45] Sahin U, Muik A, Derhovanessian E, Vogler I, Kranz LM, Vormehr M, et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature. 2020 Oct;586(7830):594-599. https://doi.org/10.1038/s41586-020-2814-7.
[46] Mulligan MJ, Lyke KE, Kitchin N, Absalon J, Gurtman A, Lockhart S, et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature. 2020 Oct;586(7830):589-593. https://doi.org/10.1038/s41586-020-2639-4.
[47] Raisch J, Darfeuille-Michaud A, Nguyen HT. Role of microRNAs in the immune system, inflammation and cancer. World J Gastroenterol. 2013 May 28;19(20):2985-96. https://doi.org/10.3748/wjg.v19.i20.2985.
[48] Han Z, Estephan RJ, Wu X. MicroRNA regulation of T-cell exhaustion in cutaneous T cell lymphoma. J Invest Dermatol. 2022 Mar;142(3 Pt A):603-612.e7. https://doi.org/10.1016/j.jid.2021.08.447.
[49] Mishra R, Banerjea AC. SARS-CoV-2 spike targets USP33-IRF9 axis via exosomal miR-148a to activate human microglia. Front Immunol. 2021 Apr 14;12:656700. https://doi.org/10.3389/fimmu.2021.656700.
[50] Kyriakopoulos AM, Nigh G, McCullough PA, Seneff S. Mitogen activated protein Kinase (MAPK) activation, p53, and autophagy inhibition characterize the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spike protein induced neurotoxicity. Cureus. 2022 Dec 9;14(12):e32361. https://doi.org/10.7759/cureus.32361.
[51] Barreda D, Santiago C, Rodríguez JR, Rodríguez JF, Casasnovas JM, Mérida I, et al. SARS-CoV-2 spike protein and its receptor binding domain promote a proinflammatory activation profile on human dendritic cells. Cells. 2021 Nov 23;10(12):3279. https://doi.org/10.3390/cells10123279.
[52] Chaudhary JK, Yadav R, Chaudhary PK, Maurya A, Roshan R, Azam F, et al. Host cell and SARS-CoV-2-associated molecular structures and factors as potential therapeutic targets. Cells. 2021 Sep 15;10(9):2427. https://doi.org/10.3390/cells10092427.
[53] Erdoğdu B, Kaplan O, Fidan BB, Çelebier M, Malkan ÜY, Haznedaroglu IC. Metabolomic profiling of leukemic hematopoiesis: effects of BNT162b2 mRNA COVID-19 vaccine administration. Curr Mol Med. 2025 Jun 3. http://dx.doi.org/10.2174/0115665240361878250601074746.
[54] Omo-Lamai S, Wang Y, Patel MN, Essien EO, Shen M, Majumdar A, et al. Lipid nanoparticle-associated inflammation is triggered by sensing of endosomal damage: Engineering endosomal escape without side effects. BioRxiv (preprint). 2024 Apr 18. https://doi.org/10.1101/2024.04.16.589801.
[55] Ko M, Quiñones-Hinojosa A, Rao R. Emerging links between endosomal pH and cancer. Cancer Metastasis Rev. 2020 Jun;39(2):519-534. https://doi.org/10.1007/s10555-020-09870-1.
[56] Eens S, Van Hecke M, Favere K, Tousseyn T, Guns PJ, Roskams T, Heidbuchel H. B-cell lymphoblastic lymphoma following intravenous BNT162b2 mRNA booster in a BALB/c mouse: A case report. Front Oncol. 2023;13:1158124. https://doi.org/10.3389/fonc.2023.1158124. Voir notre article : https://lepointcritique.fr/2023/07/19/une-souris-decede-dun-turbo-cancer-deux-jours-apres-avoir-recu-une-deuxieme-dose-de-vaccin-pfizer/.
[57] Larsson J, Hellstrand E, Hammarström P, Nyström S. SARS-CoV-2 Spike amyloid fibrils specifically and selectively accelerates amyloid fibril formation of human prion protein and the amyloid β peptide. 2023 Sept 1. bioRxiv. https://doi.org/10.1101/2023.09.01.555834.
[58] Francescangeli F, De Angelis ML, Baiocchi M, Rossi R, Biffoni M, Zeuner A. COVID-19–induced modifications in the tumor microenvironment: Do they affect cancer reawakening and metastatic relapse ? Front. Onco l. 2020 Oct 26. https://doi.org/10.3389/fonc.2020.592891.
[59] Theuerkauf SA, Michels A, Riechert V, Maier TJ, Flory E, Cichutek K, et al. Quantitative assays reveal cell fusion at minimal levels of SARS-CoV-2 spike protein and fusion from without. iScience. 2021 Feb 9;24(3):102170. https://doi.org/10.1016/j.isci.2021.102170.
[60] Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat Commun. 2020 Mar 27;11(1):1620. https://doi.org/10.1038/s41467-020-15562-9.
[61] Nguyen HT, Zhang S, Wang Q, Anang S, Wang J, Ding H, et al. Spike glycoprotein and host cell determinants of SARS-CoV-2 entry and cytopathic effects. J Virol. 2021 Feb 10;95(5): e02304-20https://doi.org/10.1128/JVI.02304-20.
[62] Lazebnik Y. Cell fusion as a link between the SARS-CoV-2 spike protein, COVID-19 complications, and vaccine side effects. Oncotarget. 2021 Dec 7;12 (25):2476-2488. https://doi.org/10.18632/oncotarget.28088.
[63] Tang M, Hu X, Wang Y. Ivermectin, a potential anticancer drug derived from an antiparasitic drug. Pharmacol Res. 2021 Jan;163:105207. https://doi.org/10.1016/j.phrs.2020.105207.
[64] Braga L, Ali H, Secco I, Chiavacci E, Neves G, Goldhill D, et al. Drugs that inhibit TMEM16 proteins block SARS-CoV-2 spike-induced syncytia. Nature. 2021; 594:88–93. https://doi.org/10.1038/s41586-021-03491-6.
[65] Karikó K, Buckstein M, Ni H, Weissman D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity. 2005 Aug;23(2):165-75. https://doi.org/10.1016/j.immuni.2005.06.008.
[66] Andries O, Mc Cafferty S, De Smedt SC, Weiss R, Sanders NN, Kitada T. N(1)-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. J Control Release. 2015 Nov 10;217:337-44. https://doi.org/10.1016/j.jconrel.2015.08.051.
[67] Kauffman KJ, Mir FF, Jhunjhunwala S, et al. Efficacy and immunogenicity of unmodified and pseudouridine-modified mRNA delivered systemically with lipid nanoparticles in vivo. Biomaterials. 2016 Dec;109:78-87. https://doi.org/10.1016/j.biomaterials.2016.09.006.
[68] Kayesh MEH, Kohara M, Tsukiyama-Kohara K. An overview of recent insights into the response of TLR to SARS-CoV-2 infection and the potential of tlr agonists as SARS-CoV-2 vaccine adjuvants. Viruses. 2021 Nov 18;13(11):2302. https://doi.org/10.3390/v13112302.
[69] Mulroney TE, Pöyry T, Yam-Puc JC, Rust M, Harvey RF, Kalmar L, et al. N1-methylpseudouridylation of mRNA causes +1 ribosomal frameshifting. Nature. 2024 Jan;625(7993):189-94. https://doi.org/10.1038/s41586-023-06800-3
[70] Boros LG, Kyriakopoulos AM, Brogna C, Piscopo M, McCullough PA, Seneff S. Long-lasting, biochemically modified mRNA, and its frameshifted recombinant spike proteins in human tissues and circulation after COVID-19 vaccination. Pharmacol Res Perspect. 2024 Jun;12(3):e1218. https://doi.org/10.1002/prp2.1218.
[71] Benitez Fuentes JD, Mohamed Mohamed K, de Luna Aguilar A, Jiménez García C, Guevara-Hoyer K, Fernandez-Arquero M, et al. Evidence of exhausted lymphocytes after the third anti-SARS-CoV-2 vaccine dose in cancer patients. Front Oncol. 2022 Dec 20;12:975980. https://doi.org/10.3389/fonc.2022.975980.
[72] Lacout A, Lesgards JF, Lounnas V, Azalbert X, Perronne C, Zizi M. Dealing with COVID-19 vaccine related antibody-dependent enhancement: A mini review. Arch Microbiol Immunol. 2024 Jun;8(2): 233-8. https://www.doi.org/10.26502/ami.936500170.
[73] Igyártó BZ, Qin Z. The mRNA-LNP vaccines – the good, the bad and the ugly? Front Immunol. 2024 Feb 8;15:1336906. https://www.doi.org/10.3389/fimmu.2024.1336906.
[74] Uversky VN, Redwan EM, Makis W, et al. IgG4 Antibodies induced by repeated vaccination may generate immune tolerance to the SARS-CoV-2 Spike protein. Vaccines. 2023 May 17;11(5):991. https://doi.org/10.3390/vaccines11050991.
[75] Irrgang P, Gerling J, Kocher K, et al. Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination. Sci Immunol. 2023 Jan 27;8(79):eade2798. https://doi.org/10.1126/sciimmunol.ade2798.
[76] Selva KJ, Ramanathan P, Haycroft ER, Reynaldi A, Cromer D, Tan CW,Preexisting immunity restricts mucosal antibody recognition of SARS-CoV-2 and Fc profiles during breakthrough infections. JCI Insight. 2023 Sep 22;8(18):e172470. https://doi.org/10.1172/jci.insight.172470.
[77] Eythorsson E, Runolfsdottir HL, Ingvarsson RF, Sigurdsson MI, Palsson R. Rate of SARS-CoV-2 reinfection during an omicron wave in Iceland. JAMA Netw Open. 2022 Aug 1;5(8):e2225320. https://doi.org/10.1001/jamanetworkopen.2022.25320.
[78] Shrestha NK, Shrestha P, Burke PC, Nowacki AS, Terpeluk P, Gordon SM. Coronavirus disease 2019 vaccine boosting in previously infected or vaccinated individuals. Clin Infect Dis. 2022 Dec 19;75(12):2169-77. https://doi.org/10.1093/cid/ciac327.
[79] Chemaitelly H, Ayoub HH, Tang P, Coyle P, Yassine HM, Al Thani AA, et al. Long-term COVID-19 booster effectiveness by infection history and clinical vulnerability and immune imprinting: a retrospective population-based cohort study. Lancet Infect Dis. 2023 Jul;23(7):816-27. https://doi.org/10.1016/S1473-3099(23)00058-0.
[80] Nakatani E, Morioka H, Kikuchi T, Fukushima M. Behavioral and health outcomes of mRNA COVID-19 vaccination: A case-control study in Japanese small and medium-sized enterprises. Cureus. 2024 Dec 13;16(12):e75652. https://doi.org/10.7759/cureus.75652.
[81] Feldstein LR, Ruffin J, Wiegand R, Grant L, Babu TM, Briggs-Hagen M, et al. Protection From COVID-19 vaccination and prior SARS-CoV-2 infection among children aged 6 months-4 years, United States, September 2022-April 2023. J Pediatric Infect Dis Soc. 2025 Jan 20;14(1):piae121. https://doi.org/10.1093/jpids/piae121.
[82] Ioannou GN, Berry K, Rajeevan N, Li Y, Yan L, Huang Y, et al. Effectiveness of the 2023-to-2024 XBB.1.5 COVID-19 vaccines over long-term follow-up : A target trial emulation. Ann Intern Med. 2025 Mar;178(3):348-359. https://doi.org/10.7326/ANNALS-24-01015.
[83] Martín Pérez C, Ruiz-Rius S, Ramírez-Morros A, Vidal M, Opi DH, Santamaria P, et al. Post-vaccination IgG4 and IgG2 class switch associates with increased risk of SARS-CoV-2 infections. J Infect. 2025 Apr;90(4):106473. https://doi.org/10.1016/j.jinf.2025.106473.
[84] Shrestha NK, Burke PC, Nowacki AS, Simon JF, Hagen A, Gordon SM. Effectiveness of the coronavirus disease 2019 bivalent vaccine. Open Forum Infect Dis. 2023 Apr 19;10(6):ofad209. https://doi.org/10.1001/10.1093/ofid/ofad209.
[85] Yamamoto K. Adverse effects of COVID-19 vaccines and measures to prevent them. Virol J. 2022 Jun 5;19(1):100. https://doi.org/10.1186/s12985-022-01831-0.
[86] Raszek M, Cowley D , Redwan EM, Uversky VN, Rubio-Casillas A. Exploring the possible link between the spike protein immunoglobulin G4 antibodies and cancer progression. Explor Immunol. 2024;4:267-84. https://doi.org/10.37349/ei.2024.00140.
[87] Cavanna L, Grassi SO, Ruffini L, Michieletti E, Carella E, Palli D, et al. Non-Hodgkin lymphoma developed shortly after mRNA COVID-19 vaccination: report of a case and review of the literature. Medicina (Kaunas). 2023 Jan 12;59(1):157. https://doi.org/10.3390/medicina59010157.
[88] Goldman S, Bron D, Tousseyn T, Vierasu I, Dewispelaere L, Heimann P, et al. Rapid progression of angioimmunoblastic T cell lymphoma following BNT162b2 mRNA vaccine booster shot: a case report. Front Med (Lausanne). 2021 Nov 25;8:798095. https://doi.org/10.3389/fmed.2021.798095.
[89] Sekizawa A, Hashimoto K, Kobayashi S, Kozono S, Kobayashi T, Kawamura Y, et al. Rapid progression of marginal zone B-cell lymphoma after COVID-19 vaccination (BNT162b2): a case report. Front Med (Lausanne). 2022 Aug 1;9:963393. https://doi.org/10.3389/fmed.2022.963393.
[90] McKernan K. Pfizer and Moderna bivalent vaccines contain 20-35% expression vector and are transformation competent in E. coli. 2023. https://anandamide.substack.com/p/pfizer-and-moderna-bivalent-vaccines.
[91] McKernan K (Medicinal Genomics, US), Nitta T (Tokyo Univ, Japan), Buckhaults PJ (University of South Carolina, US), König B et Kirchner JO (MMD Indep Lab, Magdeburg, Germany), Speicher DJ (University of Guelph, Canada) et McKernan K (Medicinal Genomics, US), Speicher DJ (University of Guelph, Canada), Raoult D (IHU Marseille, France), Kämmerer U (Univ. Hospital of Würzburg, Germany), Wang TJ, Kim A et Kim K, (FDA scientists, Centreville High School, US, executed at FDA White Oak Campus). Résultats dans : https://docs.google.com/spreadsheets/d/1gJj3GSrM-UJR9c6Lrcn1k8_buQkQznuUVSKuMR8_2lU/edit?gid=0#gid=0.
[92] Speicher DJ. Full Detailed Report. 2024 Sept 9. https://russellbroadbent.com.au/wp-content/uploads/David-Speicher-Report-2.pdf.
[93] Kämmerer U, Schulz V, Steger K. BioNTech RNA-Based COVID-19 Injections Contain Large Amounts of Residual DNA including an SV40 Promoter/Enhancer Sequence. Public Health Policy Law. 2024 Mar 12. https://publichealthpolicyjournal.com/biontech-rna-based-covid-19-injections-contain-large-amounts-of-residual-dna-including-an-sv40-promoter-enhancer-sequence/.
[94] Wang TJ, Kim A, Kim K. A rapid detection method of replication-competent plasmid DNA from COVID-19 mRNA vaccines for quality control. J High School Sc. 2024;8(4):427-39. https://jhss.scholasticahq.com/article/127890-a-rapid-detection-method-of-replication-competent-plasmid-dna-from-covid-19-mrna-vaccines-for-quality-control.
[95] Peková S. Quantitative Multiplex Real-Time PCR analysis of Moderna (Spikevax) and Pfizer (BNT162b2) vaccines. 2025 Mar 8. https://www.10letters.org/CzechResearch.pdf.
[96] Dean DA, Dean BS, Muller S, Smith LC. Sequence requirements for plasmid nuclear import. Exp Cell Res. 1999 Dec 15;253(2):713-22. https://doi.org/10.1006/excr.1999.4716.
[97] Duncan CN, Bledsoe JR, Grzywacz B, Beckman A, Bonner M, Eichler FS, et al. Hematologic cancer after gene therapy for cerebral adrenoleukodystrophy. N Engl J Med. 2024 Oct 10;391(14):1287-1301. https://doi.org/10.1056/nejmoa2405541. Voir notre article : https://lepointcritique.fr/2024/10/14/nouvelle-preuve-inedite-cancerogenicite-injections-anti-covid-pfizer/.
[98] Hickey TE, Mudunuri U, Hempel HA, Kemp TJ, Roche NV, Talsania K, et al. Proteomic and serologic assessments of responses to mRNA-1273 and BNT162b2 vaccines in human recipient sera. Front Immunol. 2025 Jan 27;15:1502458. https://doi.org/10.3389/fimmu.2024.1502458.
[99] OpenVaet, Syed KA, Guetzkow J, Cobalt J, Kunadhasan J, Jackson B. Pfizer/BioNTech C4591001 Trial – Audit Report – v1 (2024-05-31) Reanalysis of the data and anomalies inventoried. 2024 May 31. https://openvaet.substack.com/p/pfizerbiontech-c4591001-trial-audit?open=false#%C2%A7evidence-the-planned-comparison-of-processes-and-was-never-conducted.
[100] Fajloun Z, Sabatier JM. The unsuspected role of the renin-angiotensin system (RAS): Could its dysregulation be at the root of all non-genetic human diseases? Infect Disord Drug Targets. 2024;24(1):e140923221085. https://doi.org/10.2174/1871526524666230914114524.
[101] Hazan S, Dave S, Barrows B, Borody T. Messenger RNA SARS-CoV-2 vaccines affect the gut microbiome. Am J Gastroenterol. 2022 Oct;117(10S):e162. https://doi.org/10.14309/01.ajg.0000857548.07509.09.
[102] Beaudoin CA, Bartas M, Volná A, Pečinka P, Blundell TL. Are There Hidden Genes in DNA/RNA Vaccines? Front Immunol. 2022 Feb 8;13:801915. https://doi.org/10.3389/fimmu.2022.801915.
[103] Au delà de l’échec des injections anti-COVID, qui ont été homologuées sur la conviction des laboratoires qu’elles pourraient « probablement prévenir de nouvelles vagues pandémiques et ainsi réduire considérablement la mortalité due à la maladie » (p. 14 du rapport d’évaluation du vaccin Comirnaty de Pfizer/BioNTech), la généticienne Alexandra Henrion Caude rapporte qu’en l’espace de vingt ans, aucun des 70 essais cliniques où cette technologie a été testée, sur 17 maladies différentes, n’a franchi le stade de la phase 1-2 (Les Apprentis Sorciers, p. 84). Voire la présentation de son livre au Parlement européen, le 18 avril 2023 : https://www.youtube.com/watch?v=6HH5IyccJNk.