
doi.org/10.1093/ACPROF%3AOSO/9780195327939.001.0001
The Red Queen and Her King: Cooperation at all Levels of Life
ペンシルベニア州立大学人類学部,ユニバーシティパーク,ペンシルベニア州,16802
キーワード
協力、競争、発生遺伝学、自然選択、遺伝的ドリフト、進化、表現型の複雑性、発生コンピュータシミュレーション
概要
ダーウィンが生命を表現した、絶え間なく続く競争闘争のメタファーとして、「鏡の国のアリス」に登場する赤い女王はよく使われる。そのイメージは、競争と不公平を強調する私たちの文化にはしっくりとくるが、自然そのものにはあまり当てはまらない。生命は、競争よりも、あらゆるレベルで、どこにでもあるさまざまなメカニズムを通じて、明らかに協力し合うものである。ほとんどの種の生物は、マルサス的な生存の限界に近づいており、淘汰がその性能のわずかな違いを感知して、その遺伝的基盤を強化するような状況にはない。複数の主体の相互作用による協力は、生命の多くの基本的な側面に内在するものだが、その重要性は十分に広く認識されているとは言えない。ここでは、これが機能するための一連の原理について議論する。人類学的に興味深いテーマである「頭の発達」をコンピュータ・シミュレーションすることで、そのポイントを説明する。ある意味で、私たちの文化は比喩が逆になっている。赤い王家の方が、人間であれ何であれ、生命の本質をより正確に象徴しているのである。
あまり知られていないことだが、私たちの現代的な人生観は、鉄道王から得たものである。鉄道王の祖先は、特権階級の紳士で、私有財産による産業革命の立ち上げに貢献した。この考え方では、競争は成功のための機関車であり、自発的な平等は逆効果である。なぜなら、人間もその他の生物も、社会は自己利益によって形成され、それ故に自己利益に基づくからだ。その鉄道王の名は、チャールズ・ロバート・ダーウィン(1808-82)。
この世界観は、有名な文学作品によく表れている(図1参照)。ルイス・キャロルの『鏡の国のアリス』(1871)では、赤の女王がアリスの手を掴んで颯爽と歩き、女王の世界では、今いる場所に留まるために全力疾走しなければならない、と指摘しているのだ。女王の世界では、今いる場所に留まるために全力疾走しなければならない。1973年、著名な進化生物学者リー・ヴァン・ヴァレンは、このイメージを進化論の標準的なイメージとし、進化の競争的警戒は決して休まることがないというダーウィン的な考えを示した。もし変異が生じ続け、集団が過剰に増殖し、Thomas Malthusが言ったように資源を激しく奪い合うなら、個体は適応度忘却の縁にあり、そのため自然選択はわずかな優位性も見抜いて人生の勝者を特定することができる、この点はDarwinが繰り返し強調したところだ。「適者生存」の世界では、残りの人間はウサギの穴の中に消えていくだけなのだ。
ヴァン・ヴァレンの言う「進化的軍拡競争」のイメージは、現在の生物学の世界観に合致しているからこそ、深く心に響くのである。また、70年代には軍拡競争が顕著であったとすれば、子供の頃に好きだった物語にイメージを結びつけることは、進化淘汰論の大義を傷つけるものではなく、評論家、アマチュア、専門家を問わず、糧となったのである。このキャッチーな仮説は、理論的、直感的なあらゆる種類のコメントを引き出し、人気のある本も書かれた(Ridley, 2003)ほどである。この比喩は、ヴァン・ヴァレンのニュアンスに富んだ考えをはるかに超えて、独自の生命を持つに至った。長年にわたって例外はあったものの(例:Hooton, 1930; Waddington, 1959; Washburn, 1963)、人類学者は概してこの世界観に徹底的に傾倒してきた。しかし、その購入はどれほど熟慮されたもので、どれほどの代償を払ったのだろうか。赤の女王のイメージは、人間、霊長類、その他の生命を描写するものとして、どれほど正確なのだろうか。
この問いに答える一つの方法は、それが明らかに間違っていることを指摘することである。ダーウィンは生命を絶え間ない競争的闘争と見なした。しかし、生命と進化はしばしば混同されるが、同じではないし、生命がすべて競争的であるわけでも、生物が常にマルサスの奈落の底に沈んでいるわけでもない。
競争は確かにこの世に存在するが、生命を執拗な長期的進化闘争としてではなく、さまざまな時間的側面から理解する方がはるかに建設的であり、広く応用できる。生命は瞬間から瞬間に発生し、進化は想像を絶するほど多くの局所的な事例の累積効果に過ぎない。ダーウィンは「種の起源」の中で、「時間の針が長い時間の経過を刻むまでは、こうしたゆっくりとした変化の進行は何も見えない。(ダーウィン1859)このような時間を超越した生命体の変化を直感的に理解することは難しく、チェス盤の上で競い合うクイーンについてのカラフルな比喩に変えてしまうのはあまりにも簡単である。

図1. 赤の女王はアリスを人生に引きずり込むが、何の進歩もない。キャロル(Carroll, 1871)よりテニエル(Tenniel)作画
競争は確かに世の中に存在するが、生命を執拗な長期的進化闘争としてではなく、様々な時間的次元で理解する方がはるかに建設的で、広く応用できる。生命は瞬間から瞬間に発生するものであり、進化とは想像を絶するほど多くの局所的な事例の積み重ねに過ぎない。ダーウィンは「種の起源」の中で、このように述べている。
時間の針が長い時間の経過を刻むまでは、こうしたゆっくりとした変化の進行は何も見えない。(ダーウィン 1859) このような時間を超越した生命体の変化を直感的に理解することは難しく、チェス盤の上で競い合うクイーンについてのカラフルな比喩に変えてしまうのはあまりにも簡単である。
実際、ラマルクの時代から、直接観察するには遅すぎるプロセスを、遥かな時間を超えても変わらないとして、外挿することの問題は認識されていたのだ。実際、生命を進化論的に見るには、必然的に過去を遡る必要がある。化石の手足や顎、歯を現生種のものと比較することで、その形質の性質や変化を原理的に説明するために、化石の点と点を結ぶ仮説が導かれるのだ。しかし、レッド・クイーンは、”in principle”を”in fact”と仮定し、脚や顔や歯列が何らかの競争上の優位性を付与したために進化したという競争シナリオを再構築しようとし、仮説を立てるために点と点をつなぐ遊びをするのだ。決定論的、競争的な自然淘汰という想定された力のようなプロセスで点と点を結ぶことが、階層的に競争する商業社会にうまく適合し、帝国時代の鉄道王たちの関心を集めたのである。
ダーウィンの思想は、生命科学やそれ以外の分野に大きな変革をもたらした。しかし、競争中心の説明の延長が、ダーウィンの時代以降、いかに西洋の思想に深く浸透したかは容易に理解できる。優生学や大量虐殺のような恐怖さえも、自然の選択的決定を自らの手で行うことによって社会を救うという進化論的レトリックによって実行されてきた。ビジネス界を適性や淘汰の観点から説明することは日常茶飯事であり、もちろん文化人類学のダイナミクスをダーウィン的な用語で翻訳する試みも行われている。”ミーム”が遺伝子の代わりになるのだ(ボイドとリチャーソン、1985年、ドーキンス、1989)。しかし、類似と同等は同じではなく、ダーウィン的思考の不適当な流出に関する議論はGould (2002)にある。
現代の例やアナロジーを受け入れるならば、何千世代にも及ぶ生命を単純に比較することで「ダーウィン的」な生命観が、競争力の強さや一貫性についていかに単純化した考えを生み出すかは理解できるだろう。このような連続的なプロセスを理解することは、微積分で学ぶことであり、連続的で普遍的な自然の法則に関するニュートン的な考え方が受け継がれているのだ。しかし、ダーウィン自身が主張したように、またゲノムの広範な証拠が示唆するように、選択が一般的に非常に弱いとすれば、進化で起こることの多くが偶然と人口動態(遺伝的ドリフトと集団力学)に依存し、通常信じられているよりも不規則でエピソード的である場合がある(例えば、Lynch 2007)。多くの著名な生物学者が指摘しているように、ダーウィン流の厳密な選択的ストーリーは簡単に「まあまあの空想」に崩壊してしまう(Waddington, 1959; Gould and Lewontin, 1979; Lewontin, 2000; Gould, 2002)。適応には多くの一般的で比較的に一貫した方法がある(Waddington, 1959; Gottlieb, 1992, 1997; Oyama, 2000; Oyama et al., 2001; Jablonka and Lamb, 2002; Olding-Smee et al.) 特定の胎内影響は、遺伝する「エピジェネティック」な影響(ヌクレオチド配列を変化させないDNAの化学修飾)を含み(Wongら 2005;Gilbert and Epel 2009)、出生前に発達する形質や、病気のように後年にのみ発生する形質にまで影響を及ぼす可能性がある(Gluckmanら 2008)。
人類学者もまた、進化には重要な非適応過程があることを指摘している(Weaver et al.) 霊長類学者の間では、社会生活の基本的側面として、それ自体のための協力という考え方に関心が高まっており(Sussman and Chapman, 2004 Fuentes, 2010; p15)、またこの分野の観察者の間でも(Pope, 2004など)、行動進化の原動力としての競争と比べると、この種の行動は控え目であるという考えが根底にある。
ほとんどの形質について進化的説明を思いつくことは容易だが、その妥当性を示すことは難しい。実際、なんちゃってストーリーテリングが蔓延している。文学ではこれを”神話”と呼ぶが、科学でも同様の扱いをする(あるいは否定する)べきだろう。それなのに、私たちは、ダーウィンの神話がふさわしくない状況に当てはめて、物語を語り続けているのである。このような主張が議論される場合、一般的にはどの話が「真実」だろうかという議論であって、どれが真実か、もっと重要なことは、どうやって知ることができるのか、ということではない。
弱い選択の性質について直感的に理解するために、ある遺伝子座における遺伝的選択肢の選択的差異の大きさがおよそ10-23のオーダーであるとすると、これは一般的なものよりも高いかもしれないが、ある霊長類の群れやヒト科の一団において、「有利な」対立遺伝子を持つ者は1000の子を生み、「悪い」対立遺伝子は「わずか」999しか生まないということになる。有利と不利の混沌の中で、ゲノム全体では、有利なものが固定され、不利なものが犠牲になる確率は高いかもしれないが、その運命は決して確実ではない。特に、選択的優位性が、ゲノムの他の部分、環境、競合する種、そして同じように「適合」する代替方法などの変化によって、時間の経過とともに変化する場合はそうである。勝者総取りの世界観では、勝つ、あるいは一番になる方法は一つしかなく、最終的にはそれが「総取り」になると仮定しているが、これは明らかにフィクションである。日常的な観察によれば、赤の女王が支配した40億年後でさえ、人生において同等の成功を収める方法はまだたくさんあるのだ。赤の女王や適者生存、あるいは勝者総取りの表現を持ち出す有識者の多くは、適性についてそこまで厳格に考えるつもりはなかったと反論するだろう。しかし、人類学者がよく知っているように、人間は象徴する動物であり、メタファーには私たちの行動に影響を与えたり、行動を指示したりするような意味合いがある。そして私たちは、部族の旗印に簡略化したトーテムを掲げることに弱いのだ。
数十から数百の個体からなる霊長類の部隊においても、散在する非部隊の霊長類においても、先祖代々の人間の部族においても、小さな選択的差異を意味のある統計的有意性をもって検出することは原理的にさえできないのだ。たとえ小さな優位性が何千世代にもわたって最終的に優勢となるのが普通だとしても、以下に述べるように生物形質の複雑な協同的性質は、ほとんどの場合、多くの個々の遺伝的要素が適応進化に寄与し、個々の重要性が強いものはほとんどないことを意味している(Weiss 2011; Weiss and Buchanan, 2011)。このような小さな違いに注目することは、生命そのものというよりも、私たちの文化を反映したものである。
混乱の原因の一つは、自然を容赦なく赤く、歯も爪も冷酷な存在とする一般的な特徴に見られる。宗教的な意味での親切な神様の呼びかけはともかく、植物や動物など、ほとんどの生物は悔い改めることなく厄介な死を迎えるというのが明白な現実である。外からなら捕食者に、中からなら寄生虫に、生きたまま食べられてしまうのだ。この絶え間ない生存競争は、ダーウィンの思考に深く影響し、影響を及ぼした。しかし、死が冷厳な事実だろうからといって、死が常に、被相続人の特定の遺伝子型や表現型に作用する自然淘汰によって起こることを意味するのではない。ダーウィニズムが過大評価されていることを指摘するのに、現実に目をつぶる必要はないだろう。しかし、私たちがここで言いたいのは、そのような議論に巻き込まれることではなく、生命には進化以外にも注目すべきことがあるということを指摘することである。
進化論を超えて
ドブザンスキーの有名な主張[Dobzhansky, 1973]を引用すれば、「生物学で意味をなす」唯一のものとしての進化に過度に注目すると、生命のもっと多くの部分を、もっと多くの時間にわたって特徴づけているものから注意をそらしてしまうことになる。生命のより基本的な時間スケールでは、主要なプロセスは非常に異なっている。細胞や、細胞が構成する生物の一生で何が起こっているかは、進化の時間スケールで何が起こっているかを理解するのに役立つだけでなく、進化論が扱っていない他の多くの問題にも関連している。ドブザンスキーの主張は、学校で創造論に抵抗するのには好都合だったが、生物学としては明らかに事実と異なっているのである。

科学に対する次元的な挑戦
実際、生命とは記憶を伴う細胞継承がすべてであり、それは進化的(Evo)時間次元ではなく、図2に模式的に示すように、個体の一生の間に細胞が分裂・分化する発生的(Devo)、生態的(Eco)時間次元で蓄積されるものである。
進化の単純な理論的ストーリーは、世代を経たゲノムの変化の積み重ねで成り立っている。より優れた、あるいはより幸運なDNA変異株が存続するのだ。ここで「遺伝的」ではなく「ゲノム的」と表現したのは、個々の変異株はDNAの一カ所で発生するが、その運命はその時点のゲノムの残りの部分に依存することが多いからだ。進化論は、ゲノムが何をするかということを抽象的に、通常は非特定的に説明するものである。進化論は、ゲノムが何をするかということを抽象的かつ非特異的に扱うものである。そうでなければ、赤い王冠のイメージで言えば、相続放棄されるのである。しかし、ふるい分けは非常にゆっくり行われるため、通常は直接観察することができず、わずかな化石記録を除いて、過去の生命の数十年は、現在ここにあるものとの比較分析によって間接的に推測するしかない。
生命のデボとエコの次元で起きていることは異なる。また、生物の中にも時間的なダイナミズムがあるが、それは、基本的に同じゲノムを持つ生物内の細胞間のものである。最近まで、このダイナミックは複雑すぎて、ゲノムレベルでは理解されていなかった。その代わりに、形質に対する遺伝子が存在すると仮定すれば十分であり、「遺伝子」を原理的には配列に置き換え、それらが進化してきた執拗な闘争について、検証不可能ではあるが、比較的に比喩的なシナリオを呼び出すことができる。
同じことは、生態系のスケールでも、長い間行われてきた。生物は相互作用しているが、それはゲノムのおかげであり、相互作用をうまく行っている生物はゲノムを進化時間軸に落としている。ここでも、生態学的現象は「遺伝的」であると逆変換できる。なぜなら、結局のところ、実行可能な相互作用だけが、今日、重要なスケールでその痕跡を残すからだ。
つまり、進化論に包囲されると、すべてがエボ時間軸上の遺伝子に変換されてしまうのである。そして、その時間軸上では、過去という圧縮された時間スケールのために、今日では通常ニュートン的な連続した軌道と考えられているが、すべてが、重要なものだけがそこにあり、それは当時から現在まで絶えず変化しているという一見明白な考えを支持しているように見えるのである。そして、最も重要なことは、それが生存者のラインであるため、ほとんど反論の余地なく、選択によって説明されることである。このような見方をすれば、生命は明らかに競争がすべてであり、複雑な発生や組織構造を一貫したダーウィン的枠組みに当てはめる試みが注目されている(例えば、Buss, 1987; Bonner, 1988, 2000; Gould, 2002)。仮定に反論できないから、赤の女王が支配する。
つい最近まで、遺伝子が実際の生物をどのように生み出すかについての知識は、かなり初歩的なものだった。そのため、ダーウィン以降の進化生物学は、遺伝子がどのように生物を作り出すかという表現論的な問題を基本的に無視することが可能だった。遺伝子そのものの性質に関する数十年にわたる知識が先にあって、その結果、進化が生物学の大部分を占め、その原理が普遍的かつ不可侵なものとして公式または非公式に広く採用されることになったのである。しかし、現在わかっていることは、むしろそれとは異なることを示唆している。表1に示すように、生命の基本的な属性は、エコ/デボのタイムスケールでは、エボのタイムスケールと同じではない。この違いは些細なものではなく、生物学的複雑さを進化論と同様に普遍的かつ基本的な原理で理解することが可能になってきているため、無視することはできなくなりつつある。そして、最も深い違いのひとつが、協力関係である。競争的)自然淘汰による進化を考えるのではなく、協力による進化という観点から考えるべきなのだ。もし、人生を支配する側で考えるなら、赤い王様と赤い女王様が王位を分け合っている、つまり人生は家族のようなものだ。しかし、その前に意味論的な余談が必要であろう。
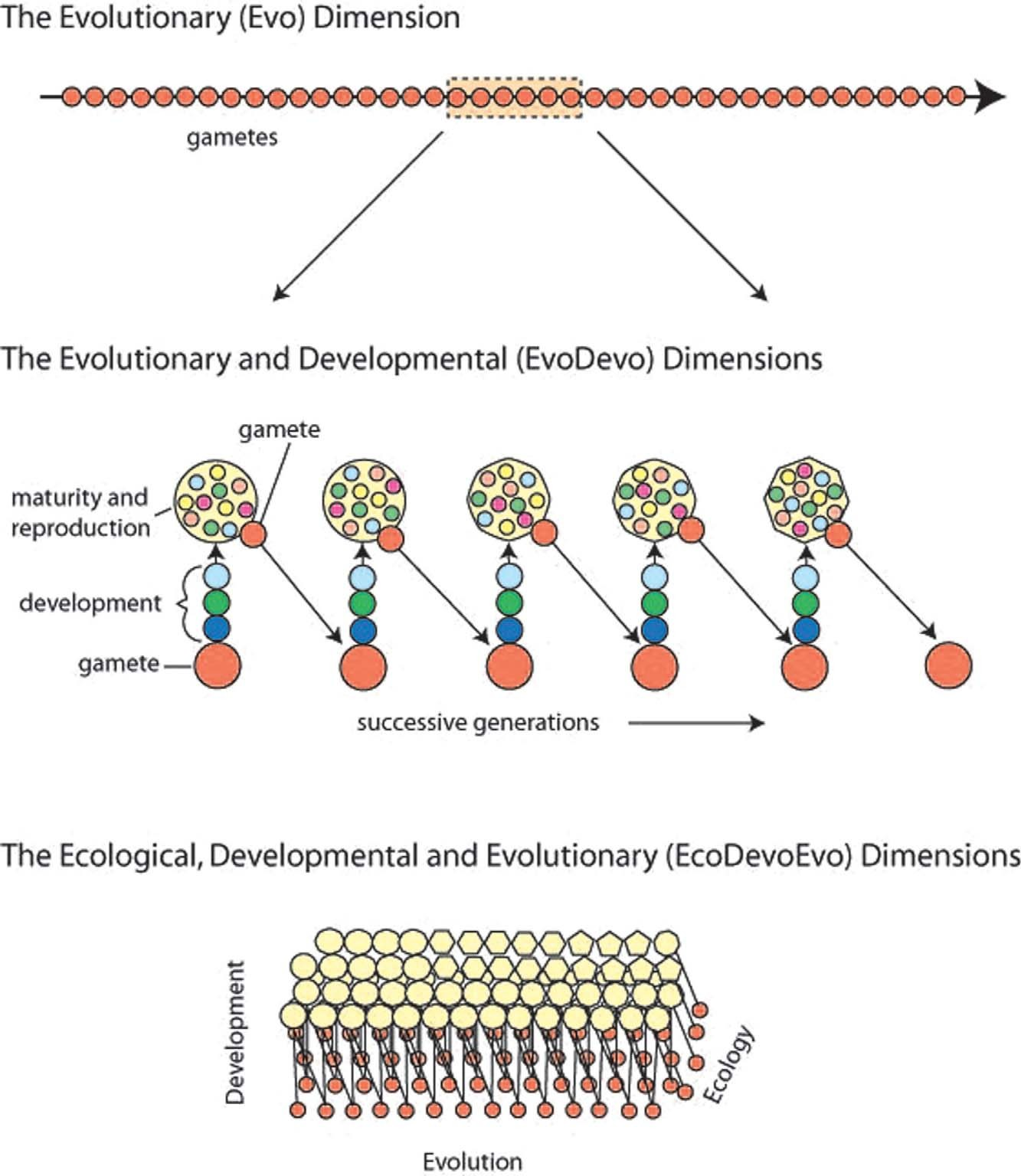
図2. 生命の時間軸 [カラー図は、wileyonlinelibrary.comで公開されているオンライン版で見ることができる]
表2. 生命の共有原理
- 記憶を伴う継承 すべての生命は連続した一つの歴史であり、DNAのコード化された性質に情報が埋め込まれている
- 隔離 生物内および生物間の生命の構成要素は、部分的に互いに隔離されているため、それらは分化し専門化することができる 分化・特化することができる。
- モジュール化 生命は、相互作用するシステムに基づく情報、構造、生理の単位で組織化されている コード化 生命の多くは、コードとして機能する構成要素の相互作用に基づいている:それらは、時間的、空間的な組み合わせで情報を運ぶ。しかし、構成要素の物理的性質は、それらがコード化する物理現象に対して恣意的である。
- Contingency(偶発性) 生命は将来の方向性よりも現在の状況に対応し、変化するものであり、現在の特性や能力によって制限される。
- 適応性 様々なレベルで、生命は状況に応じて変化する。
- 協調性 分子から生態系に至るまで、生命の機能は多くの異なる構成要素間の相互作用の成功に依存しているため、協調することは先に進むことと同様に重要であり、また必要である。
- 偶然性 生命に起こることの多くは確率的であり、方向性はない
用語と意味合いについての簡単な余談
生物学者が「競争」や「協力」といった言葉を使うのは、それぞれ野球や鍋料理のようなものを指しているのでない限り、おそらく不適切であろう。そうでなければ、私たち、特に人類学者は、文化的な用語を、あたかも事前の意味づけの負担がないかのように借用していることを認識しなければならない。
例えば、競争は自然淘汰の観点から数学的に定義することができるが、淘汰を「競争」(ましてや「利己的」等)と表現するのは誤解を招きかねない。葉の上にカモフラージュしているのに食べられない緑色の虫は、その意味では、葉に着地してすぐに鳥に見つかって飲み込まれてしまう赤い虫と競争していると言えるかもしれない。数学的には、「緑」の遺伝子の方が「赤」の遺伝子より優れているかもしれない。
数学的には、”緑”の遺伝子の方が”赤”の遺伝子より優れているかもしれない。しかし、この虫たちは、サッカーチームや企業のように互いに競争しているわけではない。しかし、この意図的な、あるいは認知的な競争という微妙なニュアンスが、しばしば原理主義的な形で、生物学の多くを支配してきた。
ダーウィンの言葉もまた、微妙な勝者総取りのイメージを呼び起こす。「適者生存」は最も一般的なもので、賞はただ一つ(進化上の成功)、それを得る方法はただ一つ(特定の有利な遺伝子型)である。それ以外の人たちにとっては、絶滅という悲惨な結果になる。数学的な利便性から、集団遺伝学では通常、遺伝子座の対立遺伝子の運命を、最も適合する単一の遺伝子型に対する各遺伝子型の適合度を割り当てることによってモデル化している。
主観性が忍び寄る例として、「正」「適応」「方向性」といった古典的な用語に代わって、「選択的掃引」という用語が広く使われていることが挙げられる。選択的掃引は、ある遺伝子の近傍で遺伝的変異が減少している状況を説明するためによく使われ、その遺伝子に対する強い正の自然選択の結果であると推定される。例えば、FOXP2は、言語プロセスに障害がある家系でこの遺伝子の変異が見つかっていることから、ヒトの言語の起源を担う遺伝子であると提唱されている。ヒトではこの遺伝子付近の変異が減少していることから、この遺伝子は言語との関連から強い選択の対象となり、その結果、選択的掃引によるものだという説がある(例えば、Enardら 2002)。しかし、このようなヒッチハイクは古典的な用語からも予想されるが、チンギス・ハーンが無抵抗のまま草原を咆哮し、行く手にいる無力なキリスト教徒の乙女をすべて略奪するというようなイメージを思い起こさせる「スイープ」ほどテレジェニックなものでもない。
何世代にもわたる進化のスケールだけでなく、細胞の世代(あるいは細胞内)のスケールや、個体、種、細胞、細胞の構成要素など、同時代の人々の間で起こる生命の構成要素の相互作用についても、理性的で一貫した理解を可能にする、通常のダーウィン的視点よりも広い枠組みを取ることが可能であり、実際に簡単かつ自然なことなのである。
以下に述べるのは、”協力”を強調する生命観である。それは、”競争”と同じく文化的な意味合いが強い。代わりに”機能的相互作用“と言うこともできる。しかし、”差動増殖”のような中立的なものではなく、競争的なレトリックが、強いダーウィンの視点から生命を表現するために広く使われている限り、私たちは、競争的なレトリックを使うことはできないと思う。
ダーウィンの視点から生命を説明するために、「差動増殖」のような中立的なものではなく、競争的なレトリックが広く使われている限り、「協力」は正当な修正であると感じる。
また、より広い視野を持つことで、説明が困難なヒトの形態的形質の本質(時に風変わりなもの)に関するような進化的な疑問も解決できる(例えば、Held 2009; Held 2011)。また、この枠組みは、人類学者が関心を寄せる人類の進化や行動に関するより良い説明を提供するために、研究デザインを策定し、結果を評価する際にも重要である。
進化の時間軸を越えてダーウィンの原理よりも一般的な原理発達遺伝学の最近の進歩により、モノとしての遺伝子では発達過程を説明できないことが分かっていた。受胎以降、生物の身体的・行動的形質は、ゲノムの機能的側面によってもたらされる協調的要因の論理的あるいは「ブール」的関係の結果である。これらの関係は、体内の適切な四次元(空間と時間)の場所に、組み合わせ的に存在、不在(ある/ない、および/またはない)、数量があることだ。古典的なタンパク質コード遺伝子そのものは、絵の一部に過ぎない。発生過程で遺伝子発現を制御している制御配列や多数の非翻訳RNAは、タンパク質をコードしていないが、遺伝子相互作用の制御において基本的に重要である。現在では、生物が相互作用し、その生涯を終えるまでの生態学的、発生学的時間スケールと、新しい機能が生まれるまでのより長い進化的時間スケールに一般化できるいくつかの基本的一般原則を見出すことが可能である。
表1では、さまざまな時間スケールにおける生命の基本的な特徴を対比させた。違いがある一方で、生命が一元的な現象であるならばそうでなければならないように、いくつかの共通項がある。これらの共通点のいくつかを表2に展開した。私たちは、他の場所でこれらについて詳しく論じているが(Weiss 2010a; Weiss and Buchanan, 2004, 2008, 2009)、ここでは簡単に説明することができる。これらの原則は互いに大きく関連しており、もちろん、これらの原則(あるいは、生活のどこにでもある他の同様の特徴)を表現するもっと良い方法があるかもしれない.ここでは生命の”高次”レベルの特性について述べているが、生命の化学の基本である中核的な代謝反応など、さらに基本的な原則もある(Morowitz et al. 2000)が、ここでは論じない。
生命の本質を知るための全体的な鍵は、記憶を伴う遺伝である。ここでいう継承とは、親から子への遺伝情報の受け渡しを意味するが、他の時間スケールでは、エピジェネティックに獲得された状態の継承や分化した細胞の特性の継承を意味することもある。進化の時間スケールで見ても、すべての細胞は、生命の始まりに遡って他の細胞から生まれたものであり、最初の細胞の記憶を宿している。なぜなら、個々の細胞が分裂するとき、その分裂産物は親細胞のDNAとその他の構成要素、そして機能状態を受け継ぐからだ。
マウスはマウスを、ライオンはライオンを生むが、胃や筋肉の細胞は胃や筋肉の細胞を生むというように、記憶を伴う遺伝が生命に予測可能性を与えている。なぜなら、糖や脂質などの他の遺伝性物質には、DNA(およびRNA)のような情報源としての「記憶」がないからだ。しかし、細胞の状態や受精卵の状態は、ゲノムと同様にRNAやその他の内容物に基づいている。
記憶を伴う遺伝という考え方は、何らかの隔離、つまりある種のユニットの分離を意味する。なぜなら、完全に均一であれば、化学反応や多様化、ひいては細胞内や生物・種間の分化が起きないからだ。これはある意味で、ダーウィンの「混合」遺伝説の根本的な問題であり、メンデルの研究によってDNAの中に遺伝子のコード化要素が隔離されていることが発見されるまでは、そうであった。分化は、それが意味するところでは、完全な混合が起こらないように隔離することが必要である。しかし、生命が時間的連続性を持つためには、与えられた状態が伝達可能でなければならない。
隔離された単位への分割は、今度は構造と機能のモジュール性を意味する、あるいは別の言い方である。モジュール化とは、毛、腸絨毛、脊椎骨、手足、指、目、色彩縞、歯、頭蓋板などの構造の繰り返しを含む、様々なものを意味する。近代生物学の歴史において多くの著者が理解してきた生物の様々な側面のモジュール性は、おそらくダーウィンのフジツボ分節に関する著書、ベイトソンのホメオティック構造の研究(ベイトソン、1894)、グレゴリーの動物構造における「ポリイソメリズム」の考え(グレゴリーら、1951)など、多くの著者に象徴されている(例えば、Webster and Goodwin, 1996など)。しかし、これらの著者は一般に特定の構造にのみ注意を向けているのに対し、モジュール性はそれよりもはるかにユビキタスである。細胞そのものは物理的に独立したモジュール単位であり、複雑に隔離された内部モジュール(ミトコンドリア、核、リボソーム、ゴルジ装置、その他多数、細胞構成要素の極性差も含む)を含んでいる。タンパク質は、通常、機能ドメインが分かれており、ゲノムは、染色体に沿った異なる配列要素が異なる独立した機能を持つという点で、卓越したモジュール構造である。そして、種は個々の生物からなり、生態系は個々の種からなる。
モジュール単位は、それが必要とする特定の条件によって機能が発揮される隔離された領域を提供する。しかし、たとえゲノムレベルであっても、またそれ以上のレベルであっても、隔離は完全にはできないのが普通である。生命の構成単位は、さまざまな方法で互いに連絡を取り合っている。このようにして、ゲノムレベルから始まり、細胞、器官、生物、そして生態系に至るまで、より低いレベルの組織からより高いレベルの組織が構築されていくのである。共通するのは、自分では予測できない外部の状況を感知し、それに応じて変化する能力である。
このユニット間のコミュニケーションはどのような性質を持っているのだろうか。それは一般に、機能的に任意な、さまざまな種類の組み合わせコードに基づいている。コンビナトリアルコードとは、相互作用する因子の適切なセット-組み合わせ-が、適切な場所、適切な時間、満足できる濃度で存在することである。この「満足」という言葉は比喩的にとらえる必要がある。なぜなら、ある結果をもたらす経路は複数存在することが多く、また濃度レベルにも許容範囲があるからだ。
人生はブーリアン的な冒険である。このようなコードの組み合わせの性質から、私たちはこれを「表現論的論理」と呼んでいる。つまり、遺伝子型が表現型につながるプロセスは、手を取り合ってフィットしなければならない分子の相互作用に基づく論理であり、それが起こると、他の分子の相互作用のカスケードが引き起こされる(図3参照)。あるシグナルが送信され、受信されると、発生カスケードは一回転する。しかし、シグナルが送信されなかったり、細胞がシグナルを受信する準備ができていなかったりすると、別の回転をする。組織学的なレベルでは、これは主に、細胞が検出する準備をするために、細胞の外部にあるシグナル分子が関与している。よくある例では、ある細胞から分泌されたシグナル分子は、拡散または循環して、適切なシグナル受容体分子を表面に持つ他の細胞のそばを通り過ぎる。シグナルが結合することで、受容体の内部構造が変化し、それが他の数十から数百の分子間相互作用のカスケードを引き起こす。最終的な結果は、受信側の細胞が発現する遺伝子の変化、あるいは発現レベルの変化、あるいは細胞の形状や機能における何らかの変化となる。
組合せシグナル伝達は、一種の”論理”であるのと同じ意味で”任意”なのである。同じ組み合わせの成分が、発生時や成体時に複数の文脈や異なる組み合わせで使われることが多く、物理的あるいは生理的に全く関係のない器官や形質であることも多い。したがって、シグナルや受容体分子の物理的性質、つまり分子構造には、変化した形質の物理的性質に関係するものは何もないのである。このように、線維芽細胞増殖因子(FGF)とその受容体分子(FGFR)と呼ばれるシグナルは、皮膚、脳、骨など、さまざまな場面で利用されている。しかし、これらの分子の構造には、やせっぽち、骨っぽけ、脳っぽさはない。重要なのは、これらの分子が皮膚や脳、骨を形成する細胞で発現し、検出されることである。これは、例えばヘモグロビンが酸素と結合したり、コラーゲンが繊維組織を作ったりするような、直接的な物理的機能とは異なるものである。
2万個以上のタンパク質をコードする遺伝子が、それぞれの場面で数十から数百の構成要素を含む複数の組み合わせで使われるからこそ、私たちは分化した生物と言えるのである。実際、私たちの体内の異なる細胞は、種と同じように互いに異なるので、細胞種と呼ぶことができる。例えば、脳と脳梁、足と胃は、自然界に存在するどの2つの構造物や種とも同じように物理的に異なるが、ハエとヘビとは異なり、同じ遺伝子のコピーを持つ細胞によって作られ、胚の中の共通の祖先細胞からハエとヘビの世代数ほど離れた細胞分裂をしないのだ。人間を構成する「細胞種」間の相互作用は、他のどのような生態系にも劣らないほど複雑であり、同じ属性の多くを共有している。複雑な協力的相互作用は生物の本質の中心であり、実際、30-40億年前の最古の細胞化石の集合体においてすでに生命の構成要素となっていた(Schopf and Kudryavtsev, 2010)。これらのシステムは、数十億年の時間をかけて少しずつ構築されてきたものであり、後述するように、これは重要なポイントである。
組織化された生命は、各要素がそれぞれの環境、つまり自分のレベルの外側にあるものを監視し、その状況に応じて対応することに基づいている。つまり、条件がユニットの状態に影響を与え、ユニットを変化させるのだ。監視と偶発的な反応は、ユニットが完全に「ハードウエア化」されているのではなく、通常、次に経験する条件が何だろうかを「知る」ことができないことを意味する。そして、その条件が発生する変化を決定し、それ故に将来の舞台を設定する。
不測の事態への対応は、あらゆるレベルの組織とあらゆる時間スケールにおいて、生命に内在する適応性を反映している。DNAから細胞、組織、生物、そして生態系に至るまで、状況の不測の事態に対応することができるのだ。もちろん、完璧なものはなく、うまく対応できない状況もある。「成功」という概念もまた、慎重にとらえなければならない。細胞は、アポトーシス(プログラムされた自己破壊)により、うまく適応できるかもしれない。これは、発生過程で複雑な器官を形成する際によく見られることで、最大限の協力をすることができる
分子レベルでは、非常に基本的な条件下で、自動的かつ非常に速い反応が起こることがある。頻繁に遭遇する条件に対する反応は、進化によって確立されたハードワイヤードな分子間相互作用である可能性がある。しかし、より高いレベルでは、より多くの時間が関係する。細胞は、例えば数秒や数分という時間スケールで変化することもあるが、もっと長いスケールで変化することもある。生物も同様に、その複雑さと、応答を生み出すのに必要な内部変化の数に関連した形で、素早く反応することもあれば、ゆっくりと反応することもある。Evoのスケールでは、何が起こるかは競争相手や仲間など、環境の多くの側面に依存し、変化は生殖時にしか起こらないため、変化は桁違いに遅くなる。
表2に示した各特徴は、生命の時間スケールにわたって作用しているが、その方法は多少異なっている。相互作用、モジュールの隔離、コーディングは、DNA配列の細分化、細胞、生物といった現代的な単位の間で行われるという点で、エコの時間スケールで発生する。エコスケールは、最も即時性があり、最も決定的なスケールである。しかし、エコ・スケールで起こる変化は、相互作用するユニットの短い寿命、すなわちデボ・スケールにおいて、大きな、短期的な変化をもたらす。ユニットが世代を超えてそれぞれの構成要素を伝達することによってのみ、エボスケールでの変化が蓄積されるのである。これは他の時間スケールよりはるかに遅く、また一部の変化のみが継承されるため、生命の基本特性はエコ時間とデボ時間のすべての違いを生むが、エボスケールには時々しか影響を及ぼさない。
以上のような生命の特徴は、すべて複数の構成要素の相互作用を伴うものであり、このことは、些細なことであると同時に深い意味をもっている。もちろん、ゲノムの機能領域、細胞、組織、器官、生物、あるいは種など、場合によっては多くの異なる要素間の協力的な相互作用なしには、ダイナミックなものは存在し得ない。個々の要素は単独で作用することはない。彼らは、空間と時間の適切な組み合わせで存在しなければならず、それぞれが相互作用の結果で重要である。彼らは協力する:co-operate。
サルやヒトのような生物間の意図的、意識的な社会的協力は、生命のまさに中核をなす協力のごく一部に過ぎず、したがって人類学を含む生物学のすべてに関連するものである。DNAは基本的に不活性であるため、ゲノムにコード化された機能はすべて、相互作用するユニット間のコミュニケーションに関係している。遺伝子の発現には、DNAと結合する産物(多くの場合、ゲノムから事前に発現している)が必要である。しかし、これはほんの始まりに過ぎない。図3に模式的に示したように、細胞間のシグナル伝達は生物の生命維持に不可欠なものである。実際、シグナルカスケードには数十から数百の遺伝子産物が関与しており、ほとんどの細胞分化はシグナル伝達の相互作用の組み合わせによって引き起こされている。このことは、バクテリアのような単細胞生物の内部機能や他のバクテリアとのコミュニケーションから、最も複雑な多細胞生物が発生、相互作用、生殖、恒常性の維持など生命のあらゆる機能を果たすまで、あらゆるレベルの生命体すべてに当てはまることである。
図は上から順に、中央の未分化な細胞の層で、いくつかのシグナル分子に対する受容体を提示している。シグナルは、この組織層の両端で分泌され、組織層を横切って拡散する。中央の細胞は、通過するシグナルの適切な組み合わせと濃度を検出すると、分化する。ここで、丸、三角、プラスの記号は、3種類のシグナル伝達因子を表している。しかし、中央の細胞はプラス記号を検出する受容体を持たないため、プラス記号を意識することはない。同様に、中央の細胞だけが丸と三角の受容体を提示し、他の細胞はそれらに気づかない。
このように部分的に隔離されたユニット間のコミュニケーションで重要なことは、情報が異なるソースから中心領域に来ること、そして情報がシグナル分子の特定の物理的特性ではなく、その組み合わせにあることである。また、シグナルは遺伝的にコード化されているが、作用は細胞外で行われ、シグナル伝達細胞の異なる発生源から遠隔にある。これはある意味でゲノム的だが、そのメカニズムは本質的に”ブール”あるいは”論理的”である。最後に、中心細胞の分化は、シグナルに応答する多くの遺伝子産物が同様にコンビナトリアルな相互作用をすることによって起こり、最終的に受容細胞のシグナル依存的な分化(図では膨張)をもたらす。
パスツールは、「チャンスは準備された心を好む」と言った。同様に、細胞もその準備が整っていなければ、シグナルに反応することはできない。具体的には、与えられた時間と場所で関連するシグナルに適したシグナル受容体のセットを発現させなければならない。受容体とは、細胞にシグナルがあることを「知らせる」ものであり、シグナルに応答するための時間である。シグナルが検出されると、活性化を待つ他の遺伝子産物の間で相互作用のカスケードが始まり、最終的には状況に応じた遺伝子発現の変化がもたらされる。ある遺伝子が活性化するためには、隣接する制御配列や、ゲノム上の他の遺伝子によってコードされた制御タンパク質など、DNAの他のセグメントやモジュールからの情報が必要であり、おそらく異なる染色体間の物理的接触によっても、遺伝子は互いにシグナルを交換する。つまり、シグナルを検出する前に、あるいはその準備として発現パターンが生じるのだ。細胞は、拡散性のあるシグナル分子を分泌したり、表面に受容体を発現させたりすることでシグナル伝達を行う。細胞は、コラーゲンなどの細胞外マトリックス化合物のような、環境の物理的な構造的側面を検出することができる。また、塩分濃度などの化学的環境にも応答する。

臓器は生命の生理的側面を管理するために協力し合っている。ホルモンや生殖はその一例である。植物でも、根から枝に至るまで、異なる細胞間でシグナル伝達を行っている。また、生物のレベルでは、非社会的な生物でさえも、交尾のためにコミュニケーションをとっている。植物と受粉媒介者、捕食者と被食者などは、匂い、色、動きなどのシグナルによって相互作用しているのだ。
このような協力的な現象は、生命の起源に遡る無限後退の結果である。なぜなら、現在の応答性は、応答する側が有効であり、有効化する側も以前は有効であったはずだからだ。したがって、受精卵は単なるDNAではなく、発現した遺伝子や適切な化学環境などの複合体である。この意味で、生命の起源がデノボ(de novo)である以外、生命にデノボ(de novo)なものはないのである。
私たちは非常に一般的なことを述べていた。なぜなら、これらはあらゆるレベル、あらゆる時間スケールの生命に適用される一般原則だからだ。この原則は、人間や霊長類、あるいは哺乳類に限定されるものではない。また、個々の生物間の社会的相互作用に限定されるものでもない。それらは、生命におけるより高次の種類の協力の一つである。また、これらの原則は相互作用の進化に限定されるものでもない。もちろん、進化はその時々の生物の存在にとって不可欠な側面ではあるが。
実際、これらの原則の一般性を見るには、自分自身の研究について考えるのが一番である。上記の原則は、些細な意味ではなく、生命について、人類学者にとっては私たち自身を含む霊長類の生命について理解しようとしていることの深い特性として当てはまる。それは骨格系だろうか?これは、モジュール構造(脊椎骨、四肢骨、歯)を作るための反復的なパターニングによって構成されており、発生中の組み合わせシグナル伝達によって達成される(寄与する遺伝子は、Hox、Bmp、Fgf、Shh、その他のパターニングシステムが知られている)。各骨の中には、骨ン、層、骨膜などのサブモジュールと、骨芽細胞や破骨細胞などの特殊な細胞ユニットが存在する。骨細胞や骨膜上のビタミンD受容体などが、栄養素、貯蔵すべきカルシウムの要求や供給、カルシウムイオンの捕獲に関連する遺伝子ファミリーの発現、あるいは骨の成長や改造のために細胞が受ける物理的ストレスに応答することで、シグナル伝達は生涯にわたって継続される。
もし、栄養代謝、目と視覚の性質、あるいは神経の発達と脳機能に興味があるのなら、表2の原理はすべてすぐに適用可能であり、少なくとも暗黙のうちに実行されていると考えてよいだろう。あるいは、栄養と環境適応に関係するのは、分節化された腸、その断面、排泄物(膵臓)、絨毛、幹細胞クリプト、免疫細胞成分などだろうか?あるいは、歯の本数、両顎の配置、歯牙と歯根への分割など、歯列の性質とその進化?さらに、より抽象的なレベルでは、同じ原理が行動にも適用される。
重要なのは、これらの原理が漠然とした一般的な方法で適用されるというだけでなく、研究の設計(例えば、どの遺伝子を調べるか、遺伝的変異を調べるためのマウスモデルの設計、比較組織学とその進化の理解)にも、結果の解釈にも利用されるということである。これらは、単なる付随的な性質ではない。どのようなテーマであれ、たとえ生物レベルであっても、現代の研究はおそらくこれらの原則を適用しており、おそらく正式に明言はしていないだろう。
このような協力的な相互作用の基本的な性質について、新しく発見されたものは何もない。それらは非常によく知られている。個々に、あるいはその場しのぎの方法で、少なくとも2世紀にわたってその応用が行われていた。しかし、生命の特徴としてのその偏在性と基本的性質は、競争という考え方に、時にレーザーのような強さで焦点を当てる時代には、十分に評価されていないかもしれない。
明らかに、機能的な理由による差動増殖は、失敗した分子間相互作用から、同胞間の相互作用、複雑な生態系の力学まで、あらゆるレベルで適用可能である。しかし、自然淘汰の洗練されたマルチレベルの概念は、素晴らしい説明の選択肢を提供する一方で、選択的説明へのコミットメントを回避させ、他の説明との比較において検証不可能なほど開放的であるため、捉えどころのないものになる可能性もある。ここで重要なのは、ダーウィン的競争以外の生命の側面は、あらゆるレベルの物事に対する単純な選択的説明を見つけようとするあまり、見落とされてしまうということである。
ここまで、表2に示された原則のうち、一つを除いてはすべて簡単に議論してきた。偶然性”の役割はどうだろうか?これはより微妙な問題であることがわかった。表面的には、高度な組織は”偶然”では生じないという考えそのものに反するように思えるからだ。しかし、それは誤認であり、同時に偶然はおそらく通常考えられているよりも重要であり、別の扱いを受けるに値するからだ。結局のところ、私たちは偶然を、異常な出来事や少ないサンプル、あるいはメンデル分離のような確率的プロセスによる、データ中の単なる統計的ノイズと考えることに慣れており、(実験データとしては厄介である以外に)長期的な結果をもたらさないのだ。しかし、これは生命において偶然が実際に果たしている役割を正しく描写していない。
確率論、淘汰、進化偶然は広く存在するが、微妙な側面である。この用語は、ある可能性に対する確信の度合いを示すために、かなりカジュアルに使われることが多い。例えば、生命が他の場所からではなく、地球上に発生した「確率」はどの程度だろうか。コインが表か裏のどちらを出すかわからないが、十分な実験を行えば、表が出る「本当の」確率を推定することができる。この推定値は、裏返しの回数が多ければ多いほど、より良い推定値となることが分かっている。コインとコイン投げの物理学から、十分に制御された条件下では、コインは100%の確率で表が出ることが分かっているからだ(Weiss, 2009)。
細胞間の分子シグナル伝達も同様である。シグナル伝達の成功には、シグナル分子が受容体分子の前を通過し、実際に結合する確率が関わってくる。信号伝達の偶然性を理解する上で重要な前提は、コイン投げと同様、実際に知っているかどうかは別として、シグナル分子と受容体分子の数、位置、結合親和性が与えられた場合、結合事象が起こる根本的な確率が本当に存在する、ということである。そうであれば、シグナル伝達における偶然の要素は統計的ノイズの源であり、例えば、細胞、受容体、シグナル分子の数が多ければ多いほど、正常に発達する組織に対する偶然の影響が均等になる可能性がある。
しかし、これは決して生命における偶然性のすべてではない。実際、生命とは偶発的な偶然の産物である。生物の細胞から細胞への記憶による継承は、世代から世代への継承も意味する。例えば、遺伝子の発現パターンなど、細胞が受け継ぐものの中には変えることができるものもある。しかし、伝達されたDNA配列の変異は変えられない。受け継いだ細胞は、受け継いだものを使って活動するのであって、受け継いだかもしれないものを分配して活動するのではない。ある世代から次の世代に移ることは、独立して何度も実験を繰り返すこととは違う。人生において、偶然の影響は累積されることがある。子孫の受けなかったものは、王家の書庫で永遠に消え去る。
あらゆる発達段階には、偶然の役割が含まれる。胚はダイナミックに階層化された細胞発生の木である。受精卵から始まる細胞分裂によって、細胞の「運命地図」が作られ、後続の細胞は次第に多能性(幹細胞的)ではなく、特定の組織や器官の役割に特化していく。突然変異やその他の偶然の影響は、発生運命地図の影響を受けた枝にある後続の全細胞に影響を及ぼす可能性がある。このような事象は、後に器官系全体に影響することもあれば、ごく一部の細胞にのみ影響し、その組織は基本的に正常な範囲に収まることもある。この範囲には、環境因子や細胞分裂の確率的側面などによる他の偶然の事象が寄与している。実際、臓器に正常範囲というものが存在するのは、遺伝的変異と体質的変異の両方があるからだ。成人では、身長、血圧、血糖値、肝臓や肺、脳や脳梗塞の大きさや生理機能などがそのような分布を持っている。しかし、個体内では、発生時の記憶を伴う階層的な遺伝が行われるため、多くの細胞が関与しているからといって、このような形質に対する偶然の影響が「均等に」なる必要はない。階層的な現象では、異なる細胞は、コイン投げのような独立した実験を構成しないのである。
一段高い生命の時間的組織では、集団や種の相互作用は常に偶然の影響を受け、ある世代での出来事が次の世代に大きな影響を与えることがある。これは進化の核心的な特徴である。なぜなら、淘汰は同時代に存在し、競合するものに対して働くからだ。偶然の出来事が遺伝子型の頻度に影響を与えると、それは生物内の細胞間の対立遺伝子とその影響、そして将来の適応を決定する生物間の進化的な遺伝子の流れの一部となるのだ。各世代はそれ自身の実験である。
実際、ほとんどの進化は、種内の突然変異、組換え、ドリフト、環境(獲物や捕食者など他の種を含む)の変化など、偶然の効果に大きく影響されると考える理由がある。偶然性は、ダーウィンが生命は歴史的な現象であると認識する上で最も重要な要素であった隔離との分岐の、十分に評価されていない側面である(Weiss and Buchanan, 2009)。
偶然性の偏在も、適者生存や自然淘汰が過大評価されがちな重要な理由の一つである。カエデの木から落ちた種が突風にあおられて池に落ちたとしても、成木になる可能性はない。サメが魚の群れを一網打尽にし、行く手を阻む魚をすべて食べてしまうとき、生き残るのは泳ぎの速い者でも、サメが来るのを見て泳ぎ切った者でもなく、幸運な者たちである。そして、運の遺伝子は存在しない。ある対立遺伝子が自然淘汰の影響を受けない場合、集団におけるその頻度は、繁殖力と死亡率の偶然性、つまり遺伝的ドリフトによって、時間とともに変化する可能性がある。しかし、これは非常に大きな「もしも」である。遺伝的ドリフトを、コイン投げに見られるような機能しない統計的ノイズと考える人は、遺伝的ドリフトを広く誤解している。しかし、それは、生命が偶発的な一方通行の歴史であるという事実そのものが、誤った認識なのである。一旦失われた対立遺伝子は、進化の地下室から戻ってくることはない。

しかし、生命の歴史は非常に長く、遺伝的ドリフトは現在の変異の重要な要因であり、おそらく最も重要な要因である。たとえ自然淘汰がそのバリエーションを形成したとしても、生殖成功の偶然の側面も存在する。全体として、ほとんどの選択は非常に弱いので、偶然はしばしば選択に匹敵する強さを持つ。進化の過程で、選択によって形成されたはずの変異は、生き残ったものよりも偶然に失われたものの方が圧倒的に多い。実際、ゲノムには、消滅した現代の変異ほど「適合」していない重要な変異が存在している可能性が高い(Lynch, 2007; Weiss, 2010c)。実際に存在した集団では、そこから採取した実用的なサンプルはもちろんのこと、統計的に淘汰を検出することができなかったというのは、おそらく不気味である。その証拠は間接的に、そして遡及的にしか存在しないのである。
チャールズ・ダーウィンは時代の産物であり、科学においては重力の法則のような普遍的な「自然の法則」を特定していた時代であった。確率的な概念が存在する一方で、ダーウィンは進化を、生物が持つかもしれないわずかな利点さえも検知して有利にする、鷹揚な自然選択によって推進されるものと考えていた(Weiss, 2004)。しかし、形質が何百もの遺伝子に影響され、そのうちのシグナル伝達カスケードが重要な例である場合、集団内の各成分の変異は、強い選択でさえ、競合する個体間の表現型の違いにのみ作用し(Weiss and Buchanan, 2003)、ほとんどの個体にごくわずかに作用するだけである。
選択によって形質が大きく左右される場合でも、ほとんどの寄与する変異株は選択と同じくらいドリフトによって進化する(Weiss, 2009; Weiss and Fullerton, 2000)。ヒトの場合、文化の緩衝効果によって、この事実はさらに顕著になる。例えば、行動に対する文化的制約は、たとえ行動形質が遺伝的裏付けを持っていたとしても、その形質の発現を抑制し、そうでない場合よりも自然淘汰の目に止まりにくくすることができることを意味する。
このことは、鳥やハチ、そして人間における重大な事実だが、あまり理解されていないようだ。生命は、ダーウィン以来ずっとイメージされてきたような、重力のような一貫した、持続的な、決定論的な力ではない。ミツバチの場合を除けば)女王蜂がすべてではない。また、生物学的形質の因果関係の複雑さは、その進化が主に競争によるものではないことを意味している。淘汰が存在することに疑問の余地はないが、その主な効果は、非協力的な変異、つまり、生命の根幹である協力的相互作用を阻害する変異を取り除くことにあるのだろう。細胞が特定のシグナルを認識できないために胚が死ぬのは、通常の意味でのダーウィン的なものではない。この死は分子の非機能(他の分子との非協力)のせいであり、種の中の他の個体との「競争」という意味では、ごく間接的なものである。しかし、人生において意味のあることがすべて競争という言葉で定義されてしまうと、競争という言葉は意味を失い、何も表現できなくなる。この真理を理解することは、人間学や人間問題に関して言えば、特に重要なことである。戯画化されたダーウィン的な考え方は、自然の前での固有の価値についてであり、歴史は、人間社会が個人の固有の価値についての信念に基づいて行動することに正当性を見出すときに生じる危険性を示しているのである。
淘汰がどこにでもあるとしても、淘汰を見つけることは難しい
私たちが今描いた絵が正確なものであるならば、機能遺伝学と進化遺伝学の双方に帰結がある。ある形質(病気、生理、形態、あるいは行動)の変異の原因となっている遺伝子を特定することは、問題である。あまりにも多くの遺伝子が個々に小さな影響を及ぼしているため、ほとんどの状況下では検出されないのである。また、多くの遺伝子が関与する形質の漸進的進化は、検出可能な痕跡をほとんど残さないかもしれない。
現在のゲノムデータから過去の適応淘汰の”サイン”を探すための統計的手法は数多く存在する。基本的に、適応選択は新しく生じた、あるいは新しく有利になった対立遺伝子を好む。これらの対立遺伝子は頻度を増し、隣接する配列変異株も一緒に高い頻度で運ばれてくる。このプロセスの特徴は、配列変異の平均レベルよりも低いDNAの伸張である。淘汰がなくなると、突然変異や組み換えによる変異によって、この特徴はいずれ崩れるが、少なくともしばらくの間は残る。
しかし、淘汰のサインを探す経験は、むしろ落胆させられるものだった。少なくとも、皮膚の色、高地への適応、その他多くの形質のような世界的な変異の一部は、最近になって適応的に進化したように思われるので、何らかの徴候が期待できるはずだ。生物学的に意味のある「ヒット」はいくつかあるものの、対象となる形質の変動のごく一部を占めるに過ぎないのである。数え方にもよるが、ゲノム上に何万もの遺伝子とその何倍もの機能単位があるにもかかわらず、統計的に選択された痕跡を探すためにゲノムを巡回しても、わずか100程度の痕跡しか発見されないのである。そして、検出されたシグネチャーは、主に地理的に局所的ではあるが、未知の事象や適応を反映しているように見える。また、先に述べたように、進化には非適応的なプロセスも多く存在する。
重要なのは、選択の痕跡がないからといって、選択がなかったことにはならないということである。この文脈では、特定の遺伝的貢献が見つからないことは否定的な結果ではない。むしろ、生物を構成する身体的・行動的形質の分子的基盤について、私たちが知っていることを肯定的に確認するものなのである。知れば知るほど特異的でなくなることは、自然界に広く存在する多遺伝子協調性の深い重要な帰結である。このような「否定的」な結果に問題があるとすれば、科学雑誌、資金提供者、公共メディアは、複雑さではなく、ドラマチックに演出でき、議員でさえ理解できる具体的な「発見」に関心があるということである(例えば、否定的結果についての議論は、Jennions and Moller, 2002を参照して下さい)。このことは、力のような淘汰や、形質の扱いやすい遺伝的構造を呼び起こすような、高揚した「まあまあ」の物語を抑制するものでなければならない。しかし、このようなストーリーは生命科学に多く、特に人間の生物医学、人類学、そして社会科学でさえも語っていることである。
原理のモデル化
私たちは、あらゆるレベルの生命に適用できると思われる一連の原則を示し、これらの原則は、生物学のあらゆる側面に関する研究を整理し、解釈するための枠組みとして利用できる、つまり、日常的に利用されていることを提案した。生物人類学も例外ではない。ヒトや霊長類の進化、形態、遺伝子の進化や機能に関連する分子データの解釈、ヒト以外の霊長類の観察によるデータ、マウスモデルから得られた知見のヒトへの適用など、研究者はそれぞれの原則を適用し、異なる状況下で動物がどのように、なぜそのように振る舞うのか、遺伝子の進化はどうだったのか、地球人類の歴史について、理論を構築している(ごく一部の例ではある)。
私たちは、霊長類の頭蓋骨の発達と変異に関する研究を行っているが、その例を挙げて、これまで述べてきた原則を明確に適用する方法を説明することができる。私たちは、頭蓋骨の発達における本質的なプロセスを特定するために、実験結果を予測し、それに基づいたコンピューターシミュレーションモデルを開発している。例えば、頭蓋板の発生を、適切な環境に到達するまで未分化な一連の移動性細胞(移動する神経堤細胞や中胚葉細胞など)の相互作用のプロセスという観点からモデル化した。頭蓋板は、チューリングプロセス、「反応-拡散」プロセス、「活性化因子-抑制因子」プロセスなど、さまざまに知られる定量的プロセスによって確立される。基本的に、最初は未分化な場において、細胞は活性化物質と抑制物質と呼ばれる2つのシグナル伝達物質を分泌する。これらの物質は、最初は一様に分布しており、異なる速度で拡散する。細胞は局所的な活性化剤の量に反応し、より多くの活性化剤を自ら生産・分泌する。また、局所的な抑制剤の量に反応し、活性化剤の生産を抑制し、抑制剤の生産を増加させる。この濃度の相互作用により、活性化と抑制が繰り返されるゾーンの波が発生する。活性化因子の相対濃度が十分に高くなると、細胞は歯、尖頭、毛髪、腸絨毛などの特殊な構造に分化する。骨や歯は骨化するため、活性化因子と抑制因子の波動は固定化されるのだ。
このシミュレーションでは、可動性のある未分化細胞に加えて、これらの細胞がモニターする活性化因子-抑制因子のプロセスをモデル化しており、これは比喩的に間葉系細胞の下の硬膜層で起こっていると考えることができる。これは、因子の空間的な密度と拡散に関する微分方程式を用いて行われる(例えば、Bar-Yam, 1997; Murray, 2002)。さらに、毛細血管の成長についてモデル化され、テストされたプロセスに従って、その領域は脈管形成によって侵される(Sun et al. 2005;Milde et al. 2008)。再び部分的に隔離された信号の送り手と受け手として、毛細血管の「先端」細胞は、酸素が少なく、他の毛細血管細胞が存在しない領域に向かって移動する。毛細血管先端細胞は、成長するにつれて、自ら酸素を分泌する。
間葉系細胞は活性剤と(模擬)酸素の相対的なレベルを監視し、両者が適度に高い領域で、間葉系細胞は移動を止め、骨を形成する骨芽細胞へと分化する(Christley et al. 2007)。実際の標本から観察すると、新生児期やそれ以前の頭蓋骨は、頭蓋板の原初的な起源から発したかのように放射状に整然とした格子構造をとっていることが示唆される。新しい骨は、格子の端から芽を出すように将来の縫合の方向に形成され、格子の空間は時間の経過とともに埋まっていく。図4は、骨の写真から格子状の構造を示したものである。図5は、上述したこのプロセスの様々な層について、発生中の4つの期間にわたる画像のタイムシーケンスセットで、現実的なパターンの血管形成の上に、非常に現実的なパターンの骨形成を生成している。また、図6は高解像度CTスキャンで観察された実際の骨の詳細と、シミュレーションによるものである。この作品では、多くの種類の遺伝子を列挙する必要はなく、私たちの一般的な原理でプロセスを列挙している。
シミュレーションは、今回のように、実際の基礎となるプロセスを模倣することなく、臨場感を生み出すことができるので、シミュレーションが説得力を持つためには、実験による検証が必要である(当研究室で進行中の作業である)。しかし、現在では、着色パターンから、毛、羽、歯の形成、そして植物に至るまでの構造において、実際に直接的な分子的サポートを受けたシミュレーションのかなりの歴史がある(Kondo and Asai, 1995; Meinhardt 1995, 1996, 2003; Bar-Yam, 1997; Jung et al, 1998; Jiang et al., 1999; Murray, 2002; Salazar-Ciudad and Jernvall, 2002, 2004; Salazar-Ciudad et al., 2003; Lin et al., 2006; Maini et al., 2006; Cui et al., 2010; Green et al., 2010)が挙げられる。シミュレーションでは、特定のシグナル遺伝子を特定することはできないし、何種類の遺伝子が関与している可能性があるのかもわからない。しかし、いくつの主要な相互作用が必要で、それがいつ起こるかを示唆することはできる。そして、それは表2の原則に基づいて行われる。この種のモデリングは、これらの原則を説明する建設的な方法を例示していると考えている。
これとは別に、私たちが開発した進化的シミュレーション・プログラム(Lambert et al. 結果はまだ出ていないが、今述べたプロセスをモデル化し、頭蓋形状のワイヤーフレーム・シミュレーションに融合させ、頭蓋形状に対する方向性選択の対象となる孤立した子孫系統において、その形状を中立的かつ個別に進化させようというのである。
これまでに紹介した原理は、記憶を伴う遺伝、隔離、モジュール組織、コーディング、偶然性、適応性、協力、偶然性など、いずれも頭蓋骨の発達に関与しており、それぞれが私たちが構築したモデルの不可欠な特性となっている。膜性骨成長の格子状の性質は、何十年もの間、一応観察されてきた。私たちのシミュレーションでは、その格子状の形は、頭蓋の大きさや形が大きくなるだけでなく、同時に血管のような格子状のネットワークに関しても重要な方向性を持っているのである。しかし、これもまた、何世紀にもわたる生物学の観察と実験が暗黙のうちに明らかにしてきたように、生物学のどんな問題にも適用できる原理を、日々の研究生活の中で説明したにすぎない。
結論 家族の中でそれを維持する
遺伝子制御の複雑さは、20世紀初頭、近代遺伝学の創始者たちによって明示的に理解された。私たちは、最近の知識がいかに詳細を明らかにしているかを味わっていただいた。その基本的でよく認識されている性質にもかかわらず、私たちが概観した種類の点が、進化の語り部において、和らげるような、ましてや根本的な役割をほとんど果たしていないのはなぜだろうか。おそらくいくつかの理由があるのだろう。歴史的に見れば、単純なメンデルの因果関係の概念が研究課題を支配してきたのだが、その当時は、単純な主要作用を超える技術が存在しなかったのである。自然淘汰は、生命の多様性を(原理的に)説明できる単一のメカニズムを提供し、他の刺激的な信条と同様に、微妙な点や代替案が明示的かつ経験的に知られたり、関連する用語で評価されたりする前に、中心的なものとして確立されたのである。実際、ダーウィンの中心的な考え方の多くは、隔離、新しい変異の偶然の生成による分岐、そして、発生した状況下で十分に相互作用・協調しないものを取り除いた後の記憶による遺伝によって、成立する。自然淘汰による微調整は、ある意味で付加的なものである。

遺伝子が生物学的形質に関与していることは知られていたが、どれくらいの数が関与し、どのように機能しているかは分かっていなかったし、知ることさえできなかった。進化論は、遺伝子を前提条件とするもので、遺伝子は変動し、その変動は淘汰されるが、その変動が実際に何をもたらすかは、ほとんど比喩の域に留まっていた。
社会的にも、科学的にも、進化論が生命の起源とその非常に複雑な構造に関する従来の説明に対して投げかける挑戦は、注目を集めた。もしこれが、ダーウィンがハトやトウモロコシの選択的品種改良から推定できた普遍的な法則なら、同じ原理を行動や社会に外挿するのに才覚は必要ないだろう。人類学に携わる私たちは、適応的な進化のシナリオが、しばしばほとんど純粋に推測の域を出ないという明白な事実に対して、十分な冷静さを持ち合わせていない。遺伝子について実際に分かっていることとは裏腹に、科学的にも社会的にも快適な競争的世界観を想定し、それを無批判に受け入れることがあまりにも容易だったのである。
民族学者は長い間、文化は独自の条件で進化し、理解可能なプロセスを反映していると思われる様々な側面があることを認識してきた(例えば、Boyd and Richerson, 1985; Cavalli-Sforza and Feldman, 1981; Harris, 1968; Laland et al, 2010; Lumsden and Wilson, 1981; Weiss, 2010b; Weiss and Hayashida, 2002; White, 1969) しかし、そのプロセスについてはコンセンサスが得られておらず、ダーウィニアンダー由来の自然選択と遺伝の単位(例えば”ミーム”)の概念をむしろ直接的に移転することはしばしば正当化できず、無数の強い主張あるいはほとんど確たる証拠のない仮定さえ導き出されてきた。もし遺伝学者が、細胞生命の組織化の複雑さを無視して、仮説のシナリオをあまりにも単純化しすぎたとしたら、遺伝子とミームの間のつながりはさらに希薄になる。
唯物論的な見方をすれば、現代人は文化とともに、また文化のために進化してきたのであり、私たちの解剖学や生理学には数え切れないほど多くの点でそれが反映されている。行動に影響を与え、その結果、文化の側面に反映されるいくつかの遺伝子、例えば、マラリア抵抗性の遺伝子(Verra et al. 遺伝子と文化の共進化には、おそらく私たちが様々な方法で説明してきたような原理が関わっているのだろう(Durham, 1991)が、文化には明らかに独自のプロセスがあり、生物学から直接移行させることはできない。なので、文化が進化し、言葉、動詞、親族集団、トーテム集団、狩猟、領地、職業、あるいはプロセスのような、部分的隔離、相互作用、コード化などを伴う相互作用する機能的活動のような社会単位がある一方で、遺伝子、細胞、生物とあまりに近い類推はおそらく危険だろう。いずれにせよ、そのテーマは明らかにここでの範囲外である。

赤の王室には王と王妃がおり、その他にも多くの親戚がいる(図7参照)。君主制は、社会が大きな拡大家族であり、協力し合うことで生き残る集団である場合にのみ機能する。一人でなく、家族の中でやっていくことで、人生はどこまでも続く競争にならずに、この地球上で成功してきたのである。競争は重要だが、社会的な協力や競争という問題が登場するずっと以前から、生命ははるかに協力的だった。
ここで紹介した原則が私たちが考えているほど普遍的なものであれば、最も還元的なものから最も全体的なものまで、あらゆるレベルで生命の本質を理解するのに役立つはずだ。また、世界中の研究所で毎日行われているような、研究の設定や結果の解釈の指針にもなる。記憶と変異を伴う遺伝がある限り、これらの一般的な特徴は異なる時間スケールで適用され、遺伝した形質の差異的な増殖と多様化という進化が存在する。その意味で、ダーウィンの考え方は、基本的なプロセスとしては一般的である。
しかし、別の見方をすれば、そろそろダーウィン的な制約の少ない考え方を卒業する時期が来ているのかもしれない。科学が解明する限り、私たちがここにいるのは、共通の祖先からの降下という歴史的なプロセスのおかげである。生命とは、死と解体が起こる実存主義的な現象である。私たちは皆、死ぬだけでなく、生命の歴史の中でほとんどの生物は、外側から、あるいは内側から生きたまま食べられ、被害者にとっては不愉快なことでも、原因にとっては好ましいことでも、何らかの形で受け継がれることによって、死を迎えたのである。しかし、競争以外の要因も、生命の性質やその変化に影響を与えており、少なくとも自然淘汰や共通祖先と同じぐらい根本的なものである。複雑な協力関係や偶然性は、競争よりもさらに広範な要因に含まれる。これらの要因は、日常的に正当な理由よりも推測に頼った外挿が行われている深い進化のタイムスケールよりもはるかに短い時間スケールで作用している。日常生活の性質は全く異なるものであり、その違いを理解することは、私たち自身や私たちを取り巻く生物界を理解する上で重要である。
謝辞
『年鑑』への論文執筆を快くお引き受けくださった編集部の方々、そして、自らの考えをより良いものにするために有益なコメントをくれた3名の査読者の方々に感謝する。著者らは、変さらによって彼らを満足させたとは言えないが、彼らの指摘に応えようとしたことは事実である。議論は十分に幅広く一般的なものであり、完全な引用リストは不可能だが、彼らは主要なポイントを示すために代表的な論文を引用するように努め、引用した論文には他の関連文献への参照も含まれている。引用できなかった、あるいは知っている著者には、一般的に謝るしかない。図4,5、6は、現在の著者の他に、Penn State大学のJoan Richtsmeier、Kazuhiko Kawasaki、Neus Martinez-Abadias, Christopher Percival、Mt Sinai Medical CenterのEthylin Jabs、YingLi Wangなどの研究グループが発表(引用)および実施中の研究結果を反映したものである。