
Contents
www.ncbi.nlm.nih.gov/pmc/articles/PMC6470839/
2019年3月22日オンライン公開
概要
卵は、孵化までの胚の発達をサポートするために必要なマクロおよびミクロ栄養素のカプセル化された供給源である。その栄養素の完璧なバランスと多様性、高い消化率、そして手頃な価格により、卵は人間の基本的な食物として脚光を浴びている。
しかし、卵は長年にわたり、心血管疾患の発生を抑えるために卵の摂取を制限するよう栄養士に勧告されている。ほとんどの実験的、臨床的、疫学的研究は、卵がもたらす食事性コレステロールと血漿中の総コレステロールの増加との間に相関関係を証明する証拠はないと結論づけている。
卵は、高齢者や子供を含む成人にとって栄養価の高い食品であり、世界中で広く消費されている。一方、卵にはまだ解明されていない多くの生理活性物質が含まれていることが証明されており、これらの物質は疾患の予防や治療に役立つと考えられている。
本総説では、(1)鶏卵の主な栄養特性、(2)鶏卵の生理活性物質に関する新たなデータ、(3)鶏卵の組成に影響を与えるいくつかの要因について、様々な家畜種の卵の栄養価の比較も含めて概説していく。
キーワード:鶏卵、食品、栄養成分、生理活性、変動、消化、健康
1. はじめに
1968年、卵業界はアメリカ心臓協会の勧告に直面し、食事性コレステロールが高いと血中コレステロールが高くなり、その結果、心血管疾患のリスクが高まるとして、全卵の摂取を週3個未満にするよう奨励された。この勧告は、卵業界だけでなく、人々の食生活にも影響を与え、栄養価の高い食品を手ごろな価格で手に入れることができなくなった。
1995年までに、アメリカ国内のすべての食事に関する勧告を統一し、卵を再生するためのin vitroおよびin vivo研究を支援するための協調的な取り組みが行われた[1]。半世紀にわたる研究により、卵の摂取は健康リスクの増加とは無関係であり[2]、その高い栄養素含有量と多くの生物活性の点で、卵を食生活に取り入れる価値があることが証明されている[1]。
最近のいくつかの研究は、身体活動的な人々を含む人間にとって卵が有益な役割を果たすことを強調しており、また、何人かの著者は卵のコレステロールがあまり吸収されないことを実証している[3,4]。
その結果、卵の摂取が血中コレステロール濃度に大きな影響を与えることはない[3,4]。また、卵の消費者、特に6-24ヶ月の幼児は、非消費者に比べて、添加糖および総糖の摂取量が少なく[5]、これは卵の満腹効果と相関していると思われる[2,6,7]。
糖尿病、高コレステロール血症、高血圧などの代謝性疾患を患っている人は、食事からのコレステロール摂取に注意する必要があるが、卵が生涯を通じて健康全般に貢献することは、現在では十分に確立されている[8]。もうひとつの懸念は、卵アレルギーに関するものである。
卵アレルギーは一般的な乳児の食物アレルギーで、5歳未満の子どもにおける有病率は1.8%~2%と推定されている。卵に対する過敏症に関連する分子は、主に卵白に集中しており、オバルブミン、リゾチーム、オボムコイド、オボトランスフェリンが主な卵アレルゲンとなっている[9]。
また、卵黄由来のタンパク質もいくつか報告されている [9]。卵アレルギーは通常、生後5年以内に発症し、50%の小児が3歳までに卵過敏症を脱却します [10,11]。幸いなことに、ほとんどの場合、卵アレルギーの有病率は年齢とともに減少し [12]、通常、学齢期までに治癒する。
*
卵は、必須脂質、タンパク質、ビタミン、ミネラル、微量元素を含み [13]、適度なカロリー源(約140 kcal/100 g)、料理の可能性、低経済コストという栄養学的観点から特に注目されている食材である。
実際、卵はタンパク質、ビタミンA、鉄、ビタミンB12、リボフラビン、コリンの最も安価な動物性供給源であり、亜鉛とカルシウムの2番目に安価な供給源であると確認されている[14]。
幼児や成人にバランスの良い栄養素を提供するだけでなく、卵は無数の生物学的に活性な成分を含んでいる[15,16,17]。これらの成分は、卵の様々な内部成分に割り当てられている(図1)。卵殻とそれに密接に関連する卵殻膜は通常消費されないが、卵殻膜は食用に供されることを言及しておかなければならない(図1)。
世界の卵の平均消費量は62個/年(インド)から358個/人以上(メキシコ)[18]、アフリカ諸国ではさらに少ない36個/年/人([19])である。商品化されている食卓用卵は無精卵で、人間が食べるために世界中で特別に飼育された約30億羽の鶏によって生産されている。
図1 卵の構造
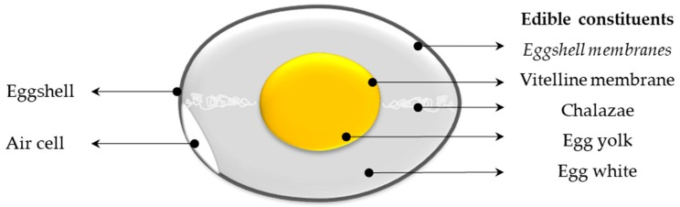
斜体:卵殻膜は食用になるが、卵殻と強固に結びついたままなので、通常は食用にしない
また、卵の成分は、特に卵を生の食材として摂取した場合、少量の卵タンパク質が同化されないものの、消化率が高いことが報告されている[20] [20,21,22].加熱した卵タンパク質の消化率が高いのは、加熱によりタンパク質の構造的な変性が起こり、消化酵素の加水分解作用が促進されるためと考えられている。
しかしながら、卵タンパク質の同化は加熱前処理により促進され、高いレベル(加熱卵白タンパク質で91-94%)に達するものの、部分的には不完全なままである。主要なタンパク質、本質的にはプロテアーゼインヒビターのオボムコイドや主要な卵白のオバルブミンは熱加熱に抵抗することは注目に値する[23,24]。
この観察は、卵由来タンパク質および卵白タンパク質の限定消化からin vitroで生成される多くの加水分解ペプチドが、人間の健康にとって興味深い生物活性を有し、したがって栄養補助食品として使用される可能性があることを知る上で特に興味深いものである[16]。
実際、それらのいくつかは、抗菌性、抗酸化性、および抗がん性の特性を示すことが示されている[25,26,27]。このように、多くの著者は、腸内のタンパク質由来ペプチドの重要性と、身体の免疫防御の第一線、免疫調節、および正常な身体機能におけるその実質的な役割を強調している[28]。
2. 卵の栄養素
卵のタンパク質は卵白と卵黄に均等に分布し、脂質、ビタミン、ミネラルは基本的に卵黄に集中している(図2)。卵の主成分は水分であり(図2)、繊維質がないことは特筆すべきことだ。卵のミネラル、ビタミン、または特定の脂肪酸の相対的な含有量は、国の基準によって異なる場合があるが [29]、水分、タンパク質、脂質、炭水化物などの主要な構成要素を考慮すると、世界的に同等であることに変わりはない。
卵の主要栄養素は非常に安定しており、卵白と卵黄の比率に左右されるのに対し、微量成分は鶏の栄養などいくつかの要因に影響される(セクション4.2参照)。全卵、生卵、産みたて卵において、水分、タンパク質、脂質、炭水化物、灰分はそれぞれ約76.1%、12.6%、9.5%、0.7%、1.1%を占める [30]。
図2 卵の可食部の基本組成
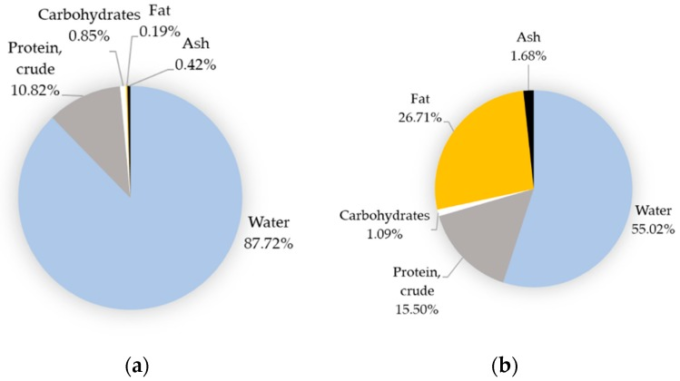
(a)卵白、(b)卵黄。なお、(b)については、卵黄・ビテリン膜複合体を指す結果である。Ciqualホームページ ciqual.anses.fr/ より2019/11/01に取得(フランス食品・環境・労働安全衛生庁。
ANSES-CIQUAL)
2.1. 大栄養素
2.1.1. タンパク質
卵白と卵黄にはタンパク質が多く含まれている。何百種類ものタンパク質が同定されており、胚の発生過程で時間的に必要な特定の生理的機能に関連している。これらのタンパク質の一部が区画特異的であることは、卵黄と卵白が異なる組織で形成されているという事実で説明できる。卵黄は基本的に肝臓に由来し、卵白は鶏の卵管で成熟した卵黄の排卵後に合成され分泌される[31]。
*
タンパク質の濃度は、平均して全生鮮卵100gあたり12.5gであり、卵膜を含む卵黄と卵白はそれぞれ100gあたり15.9gのタンパク質と10.90gのタンパク質を含んでいる。これらの値は、鶏の遺伝や年齢によって若干変更される(セクション4参照)。
プロテオミクスによる補完的なアプローチにより、卵殻を含む鶏卵では約 1000 種類のタンパク質が同定されている [32,33,34,35,36,37,38,39,40].
*
卵黄は、68%の低密度リポタンパク質(LDL)、16%の高密度リポタンパク質(HDL)、10%のリベチンおよびその他の可溶性タンパク質、および4%のホスビチンを含む複雑な環境である。これらの成分は、卵黄タンパク質の約50%を占める顆粒と呼ばれる非水溶性タンパク質凝集体(乾物の19〜23%)と、乾物の77〜81%に相当する黄色透明な液体または血漿の間に分布している[41,42]。
アポリポタンパクB,アポビテレニン-1,ビテロジェニン,血清アルブミン,免疫グロブリン,オバルブミン,オボトランスフェリンが卵黄の最も多いタンパク質で,全卵黄タンパク質の80%以上を占めている[43].卵黄は、卵黄を包む細胞外タンパク質マトリックスを形成する2つの異なる層からなる卵膜と密接に関連している[44]。これらの膜は卵黄を他の卵の区画から物理的に分離し、卵黄の卵白への漏出を防止する。
*
卵白は脂質を含まないゲル状の構造で、主に水(約88%)[44](図2)、繊維状の構造タンパク質(オボムシン)、糖タンパク質(オバルブミン、プロテアーゼ阻害剤)、抗菌タンパク質(リゾチーム)、ペプチド(セクション3.1参照)で構成されている [33,45].卵白の平均容積は 30 mL(卵殻を含む重量 60 g の卵の場合)、タンパク質濃度は卵白中約 110 mg/mL と推定される。
卵白には全部で150種類のタンパク質が同定されており[35]、その中でも非常に豊富なオバルブミンは卵白タンパク質全体の50%を占めていることが分かっている。卵におけるこのタンパク質の生理的機能はまだ不明であるが、オバルブミンは鶏胚の成長に必須なアミノ酸を供給していると推測される。
このように、卵白のオバルブミンは、ヒトの栄養摂取のための貴重なアミノ酸源となる。卵白には、オバルブミンの他に、抗菌性リゾチームが濃縮されており、現在、抗感染症剤として多くの医薬品や食品の保存料として使用されている(3.1項参照)。
卵白の粘性は、基本的にオボムチンによるものである[46]。驚くべきことに、卵白はまた、非常に豊富な4つのプロテアーゼ阻害剤[47]の存在によって特徴付けられ、特に卵白がいくつかの食品調製物の原料として使用されている場合、卵成分の消化を遅らせる可能性がある。
2.1.2. 脂質
EU各国やアメリカの卵の成分表[29]を考慮すると、卵の総脂質量は全卵100gあたり8.7から11.2の範囲で比較的安定している。これらの脂質は卵黄にのみ集中し(図 2 および表 1)、ごく一部は卵細胞膜と密接な関係を保っている可能性がある [48,49]。
表1 卵脂質 1
| 名前 | 卵、全体、生 | 卵黄、生 | ||||
|---|---|---|---|---|---|---|
| 平均含有量(g / 100g) | 最小 値(g / 100g) | 最大 値(g / 100g) | 平均含有量(g / 100g) | 最小 値(g / 100g) | 最大 値(g / 100g) | |
| FA飽和 | 2.64 | 0.05 | 3.13 | 8.47 | 7.13 | 9.55 |
| FA 4:0 | <0.05 | 0 | –2 _ | 0 | – | – |
| FA 6:0 | <0.05 | 0 | – | 0 | – | – |
| FA 8:0 | <0.05 | 0 | – | 0.009 | – | – |
| FA 10:0 | <0.05 | 0 | – | 0.009 | – | – |
| FA 12:0 | <0.05 | 0 | – | 0.009 | – | – |
| FA 14:0 | 0.024 | 0 | 0.038 | 0.091 | 0.077 | 0.1 |
| FA 16:0 | 1.96 | 0.05 | 2.43 | 6.04 | 5.03 | 6.86 |
| FA 18:0 | 0.65 | 0.05 | 0.89 | 1.73 | – | 2.42 |
| FAモノ不飽和 | 3.66 | 0.05 | 6.73 | 11.9 | 10.2 | 13.8 |
| FA 18:1n-9シス | 3.51 | 3.03 | 3.65 | 10.4 | 9.69 | 11.2 |
| FA多価不飽和 | 1.65 | 0.05 | 3.39 | 4.07 | 3.33 | 4.66 |
| FA 18:2 9c、12c(n-6) | 1.38 | 1.18 | 2.7 | 3.28 | – | 3.62 |
| FA 18:3 9c、12c、15c(n-3) | 0.061 | 0.02 | 0.58 | 0.15 | – | 0.27 |
| FA 20:4 5c、8c、11c、14c(n-6) | 0.12 | – | 0.13 | 0.37 | – | 0.4 |
| FA 20:5 5c、8c、11c、14c、17c(n-3)EPA | 0 | – | 0.003 | 0.01 | – | 0.011 |
| FA 22:6 4c、7c、10c、13c、16c、19c(n-3)DHA | 0.09 | 0.045 | 0.18 | 0.25 | 0.11 | 0.46 |
| コレステロール | 0.398 | 0.344 | 0.423 | 0.939 | 1.280 | |
1 Ciqualホームページ ciqual.anses.fr/ より2019/01/17に取得(フランス食品・環境・労働安全衛生庁。ANSES-CIQUAL)。FA、脂肪酸、EPA、エイコサペンタエン酸(オメガ3脂肪酸)、DHA、ドコサヘキサエン酸(オメガ3脂肪酸)。2は入手不可。
脂質は、トリグリセリドとコレステリルエステルのコアからなり、リン脂質とコレステロールの単層で囲まれ、その中にアポタンパク質が埋め込まれた構造を持つ卵黄リポタンパク質の一部である[42]。卵の総脂質量を変化させることは非常に困難である。
卵の脂質の増加は、基本的に卵黄と卵白の比率の増加に依存するが、この比率は鶏の栄養にあまり影響されない。一方、脂肪酸組成は鶏の飼料に大きく依存する(セクション 4.2 参照)。この変動は、表 1 に脂肪酸(飽和、一価不飽和、多価不飽和)の最小値と最大値で示されている。
卵黄の飽和脂肪酸に対する不飽和脂肪酸(一価不飽和+多価不飽和)の相対量(全卵100gあたり5.31g対2.64g、表1)は、他の動物由来食品と比較して特に高いことが特徴的である。卵黄は、リノール酸(FA 18:2 9c,12c (n-6) )などの必須脂肪酸も豊富に含んでいる。
卵はコレステロールを多く含むため(全卵100gあたり400mg)、30~40年前に卵の摂取量は減少したが、1990年代に行われた多くの研究で、卵の摂取量と高濃度の形質膜コレステロールの間に相関がないことが報告されている[3,4]。
現在では、形質膜コレステロールの変動とそれに伴う心血管疾患リスクは、食品要因だけでなく飽和脂肪酸(食事性ミリスチン酸(14:0)およびパルミチン酸(16:0)など)摂取にも起因すると考えられている。
サルおよびスナネズミを用いたin vivoでの古い研究により、食事性14:0(ミリスチン酸)および18:2(リノール酸)が血漿コレステロールを調節する主要な脂肪酸であることが示されている-14:0は、血漿コレステロールを上昇させる主要飽和脂肪酸で、18:2は一貫して低下させる唯一の脂肪酸だった [50、51]。
卵では、14:0(ミリスチン酸、全卵100gあたり0.024g)は不飽和脂肪酸18:2(リノール酸、全卵100gあたり1.38g)に比べてはるかに濃縮されていない。これらのデータはすべて、卵が健康な患者の心血管疾患発症率の上昇と関連しないことを裏付けている。
しかし、食事性コレステロールの高応答者(人口の約15%から25%)では、卵の摂取量をコントロールする必要がある。このような人々の卵消費量の増加は、低応答者よりも血漿脂質に大きな影響を与えるからである。
2.1.3. 炭水化物
卵は繊維質を含まず、炭水化物の含有率は0.7%と低い。卵の炭水化物は卵黄と卵白の間に分布している(図 2)。グルコースは卵の遊離糖の中で最も多く(全卵100gあたり約0.37g)、卵白に多く含まれている(卵黄100gあたり0.18gに対して卵白100gあたり0.34g)[30]。
生卵白と生卵黄からは、微量のフルクトース、ラクトース、マルトース、ガラクトースが検出されている[30]。糖質は卵タンパク質にも多く含まれ、その多くは卵黄、膜、卵白を形成するために鶏の生殖組織から分泌される前に翻訳後糖鎖修飾を受ける糖タンパク質であることが分かっている。
2.2. 微量栄養素
2.2.1. ビタミン類とコリン
卵、正確には卵黄は、ビタミンC(アスコルビン酸)以外のすべてのビタミンを含む、ビタミン豊富な食品である。卵にビタミンCが含まれていないのは、鳥類がグルコースからのデノボ合成により、自身のビタミンCの必要量を満たすことができるためと考えられる[52]。
モルモット、サル、飛行哺乳類、ヒト、そして進化した一部のスズメ目鳥類を含むいくつかの動物種では、進化の過程でビタミンCを生成する能力が失われている[52]。その結果、家禽類ではなくこれらの後者の種は、ビタミンCの食事源(果物や野菜)に依存している。
卵黄はビタミンA、D、E、K、B1、B2、B5、B6、B9、B12を多く含み、卵白はビタミンB2、B3、B5を多く含み、ビタミンB1、B6、B8、B9、B12を多く含む(表2)。1日2個の卵を食べることで、人間が必要とするビタミンの10%から30%をカバーすることができる。
卵黄に含まれる脂溶性ビタミン(ビタミンA、D、E、K)の含有量は、鶏の食事に大きく依存することが注目される(4.2項参照)。これらのビタミンに加えて、卵はコリンの主要な供給源であり、コリンは基本的に卵黄に集中している(卵白の1 mg/100gに対して卵黄は680 mg/100g)[30,53]。
固ゆで卵は、牛レバーに次ぐコリンの主要供給源であり[54]、米国の食事におけるコリンの最初の供給源であることが報告されている[55]。食品中では、コリンは水溶性(遊離コリン、ホスホコリン、グリセロホスホコリン)と脂溶性の形態(ホスファチジルコリン、スフィンゴミエリン)の両方で見られ、すべてのライフステージにおいて細胞の維持と成長の両方で重要かつ多様な機能を有している。神経伝達、脳の発達、および骨の完全性において何らかの役割を担っている[54,56,57]。
表2 卵のビタミン1(平均含有量;μg/100g)
| 名前 | 卵、全体、生 | 卵黄、生 | 卵白、生 |
|---|---|---|---|
| ビタミンAまたはレチノール | 160 | 371 | 0 |
| ビタミンA前駆体またはベータカロチン | 0 | 88 | 0 |
| ビタミンDまたはコレカルシフェロール | 2.0 | 5.4 | 0 |
| ビタミンEまたはアルファ-トコフェロール | 1050 | 2580 | 0 |
| ビタミンKまたはフィロキノン | 0.3 | 0.7 | 0 |
| ビタミンC | 0 | 0 | 0 |
| ビタミンB1またはチアミン | 40 | 176 | 4 |
| ビタミンB2またはリボフラビン | 457 | 528 | 439 |
| ビタミンB3またはニコチン | 75 | 24 | 105 |
| ビタミンB5またはパントテン酸 | 1533 | 2990 | 190 |
| ビタミンB6 | 170 | 350 | 5 |
| ビタミンB8またはビオチン | 16.5〜53.8 2 | 27.2〜49.42 _ | 5.7–7.9 2 |
| ビタミンB9または葉酸 | 47 | 146 | 4 |
| ビタミンB12またはコバラミン | 0.89 | 1.95 | 0.09 |
1 農務省農業研究事業団(2014)より2019/01/17に取得した。USDA National Nutrient Database for Standard Reference, Release 27. www.ars.usda.gov/ba/bhnrc/ndl 2 [58]。
2.2.2. ミネラルと微量元素
卵はリン、カルシウム、カリウムが豊富で、適度なナトリウム(全卵100gあたり142mg)を含んでいる(表3)。また、銅、鉄、マグネシウム、マンガン、セレン、亜鉛などの必須微量元素をすべて含み(表3)、鉄と亜鉛の供給には卵黄が大きく寄与している。
これらのミネラルや微量栄養素の一部(Zn、Mg、Se)の欠乏は、うつ病や疲労[59]、病理学的疾患の発症と関連しているので、卵にこのようなミネラルや微量栄養素が存在することは非常に興味深いことだ。これらの微量元素の一部(セレン、ヨウ素)は、鶏の飼料によって濃度が著しく上昇する場合がある(4.2項参照)。
表3 卵のミネラルと微量元素(平均含有量;mg/100g) 1.
| 名前 | 卵、全体、生 | 卵黄、生 | 卵白、生 |
|---|---|---|---|
| カルシウム | 56 | 129 | 7 |
| 銅 | 0.072 | 0.077 | 0.023 |
| ヨウ素 | 0.021 | 0.18 | 0.002 |
| 鉄 | 1.75 | 2.73 | 0.08 |
| マグネシウム | 12 | 5 | 11 |
| マンガン | 0.028 | 0.055 | 0.011 |
| リン | 198 | 390 | 15 |
| カリウム | 138 | 109 | 163 |
| セレン | 0.030 | 0.056 | 0.020 |
| ナトリウム | 142 | 48 | 166 |
| 亜鉛 | 1.29 | 2.30 | 0.03 |
1 農務省農業研究事業団(2014)より2019/01/17に取得した。USDA National Nutrient Database for Standard Reference, Release 27 www.ars.usda.gov/ba/bhnrc/ndl および Ciqual homepage ciqual.anses.fr/ (French Agency for Food, Environmental and Occupational Health & Safety.)から取得。ANSES-CIQUAL)のヨウ素含有量を参照する。
2.3. 抗栄養因子
前述したように、卵の主要タンパク質にはプロテアーゼ阻害剤が含まれており、ペプシン、トリプシン、キモトリプシンなどの消化酵素を阻害することによって、卵タンパク質の適切な分解を遅らせる可能性がある。実際、卵白はオボスタチン、オボムコイド、オボインヒビター、およびシスタチンの主要な供給源である[47]。
さらに、これらの分子の一部(オボインヒビター、オボムコイド、シスタチン)は、プロテアーゼや胃液による変性に対して適度な抵抗性を与えると思われる多くのジスルフィド結合を有している。これらの抗栄養素の一部は調理過程で熱により部分的に変性し [20,22,23,24] 、消化性プロテアーゼによるタンパク質の分解を促進する可能性がある。
さらに、卵に多く含まれるビタミン結合タンパク質は、ビタミンへのアクセスを制限する場合がある。ビタミンB12(ビオチン)を結合するアビジンは、リガンドとタンパク質間の自然界で最も高い親和性を示している[60]。ビオチンは、アビジンとその結合したビタミンB8との間に形成される緊密な複合体によって、消費者にとってのバイオアベイラビリティが損なわれている可能性がある。
3. 卵の栄養補助食品
卵は単に栄養価の高い基本的な食品であるだけでなく、人間の健康にとって大きな関心のある多くの生物活性化合物(脂質、ビタミン、タンパク質、および由来の加水分解ペプチド)[16,61,62,63,64]を含んでいるという証拠が増えてきている。
精製タンパク質に対して行われたin vitro分析により、卵タンパク質は多様な生物活性を示すことから、大きな可能性を持っていることが明らかになった。生物活性が期待できる加水分解ペプチドの同定には、物理化学的アプローチ、分析的アプローチ、in silicoアプローチを組み合わせた様々なツール [65,66] が使用可能である。
卵タンパク質の多くは、胚に必須なアミノ酸を提供するだけでなく、ヒトを含む卵を食べる生物種にとって、まだ生理的な機能が特定されていないことは驚くべきことだ。卵タンパク質が腸の健康に貢献しうる幅広い抗菌活性を示すことに加え、消化の過程で自然に発生する可能性のある卵由来の加水分解ペプチドの生物活性をさらに明らかにするための多くの努力がこの数十年間になされている[20,22]。
興味深いことに、これらの生理活性ペプチドの一部は、変性した卵タンパク質を茹でた後に限定的にタンパク質分解した後に特異的に生成される[67]。これらの研究のほとんどはin vitroで行われたものであるが,この発見は多くの研究分野を開くものである.現在までのところ、卵タンパク質が胃の酸性pH、消化酵素、腸内微生物にどのように抵抗するのか、また、食事中に卵タンパク質分解酵素阻害剤が存在すると、消化酵素による卵タンパク質の分解がどのように妨げられるのかについては、ほとんど分かっていない。
タンパク質消化の動態は、タンパク質のペプチドへの加水分解から始まり、ジペプチド、そして最後に遊離アミノ酸へと完全に分解されるまでの順を追ったものである。しかし、一部の卵タンパク質(オバルブミン、オボムコイド)は部分的にしか消化されないことが知られており[20,22]、アミノ酸への完全分解を受けずに、いくつかの生理活性ペプチドが自然に生成される可能性が示唆される。
3.1. 抗菌剤
可食部に含まれる卵の抗菌成分は、基本的に卵白と卵膜に集中している。これらの抗菌剤は、対象とするタンパク質によって、抗菌活性、抗ウイルス活性、抗真菌活性、抗寄生虫活性を示すことがある(表4)。
表4 主な卵の抗菌性タンパク質
| 遺伝子ID/遺伝子記号 | 対象生物 | ローカリゼーション1 | 参考文献 | |
|---|---|---|---|---|
| 鳥類のベータディフェンシン11 | 414876 / AVBD11 | バクテリア | EW、VM | [ 76、77 ] _ _ |
| アビジン | 396260 / AVD | バクテリア | EW、VM、EY | [ 78、79 ] _ _ |
| ベータマイクロセミノプロテイン様 | 101750704 | バクテリア | EW | [ 76 ] |
| シスタチン | 396497 / CST3 | バクテリア、ウイルス、真菌、寄生虫 | EW、VM、EY | [ 75、80、81、82、83、84、85 ] _ _ _ _ _ _ _ _ _ _ _ _ |
| ガリン | 422030 / OvoDA1 | バクテリア | EW | [ 86、87、88、89 ] _ _ _ _ _ _ |
| 免疫グロブリンY | – | EY | [ 90 ] | |
| リゾチーム | 396218 / LYZ | バクテリア、ウイルス、菌類 | EW、VM、EY | [ 67、91、92 ] _ _ _ _ |
| 卵白アルブミン関連タンパク質X | 420898 / OVALX | バクテリア | EW、VM、EY | [ 93 ] |
| オボグロブリンG2/TENP | 395882 / BPIFB2 | バクテリア | EW、VM、EY | [ 94、95、96 ] _ _ _ _ |
| 卵抑制剤 | 416235 / SPIK5 | バクテリア | EW、EY | [ 97 ] |
| オボムチン(アルファおよびベータサブユニット) | 395381 / LOC395381(アルファ)
414878 / MUC6(ベータ) |
バクテリア、ウイルス | EW、VM | [ 74 ] |
| オボトランスフェリン | 396241 / TF | バクテリア、ウイルス | EW、VM、EY | [ 98、99、100 ] _ _ _ _ |
| ホスビチン | 424547 / VTG1 | バクテリア | EY | [ 101 ] |
| プレイオトロフィン | 418125 / PTN | バクテリア | EW | [ 76、102 ] _ _ |
| 卵黄膜外層タンパク質1 | 418974 / VMO1 | バクテリア | EW、VM、EY | [ 76 ] |
1 可食部。EW:卵白、EY:卵黄、VM:硝子体膜。
卵の抗菌作用は、いくつかの殺菌または静菌メカニズムに依存する。中には、細菌壁と相互作用して、細菌の透過と死滅を誘発する強力な作用を持つものもある(リゾチーム、鳥類β-ディフェンシンなど)。その他の分子の効果は、いくつかの微生物の増殖に必要な鉄(オボトランスフェリン)やビタミン(アビジン)のバイオアベイラビリティを低下させたり、感染症の悪性因子である微生物プロテアーゼ(オボインヒビター、シスタチン)を阻害することによって、むしろ間接的である [68].これまでに文献に記載されている様々な卵の抗菌性分子を表4に示す。
興味深いことに、そのうちのいくつか(AvBD11, OVAX, avidin, beta-microseminoprotein) はヒトゲノムに発現していないことから[69]、これらはヒト腸管病原体に対する強力な抗感染剤となり、腸管宿主免疫を強化できる可能性が示唆された。
*
これらの卵タンパク質やペプチドに加え、外因性プロテアーゼによる部分加水分解後に放出される可能性のある卵由来ペプチドの抗菌活性を報告するデータも増えてきている。リゾチーム[70,71,72,73]、オボトランスフェリン[25]、オボムシン[74]、シスタチン[75]から得られるこのような加水分解ペプチドは、幅広い抗菌活性を示すことが分かっている。
3.2. 抗酸化活性
消化管における長期的な酸化ストレスは慢性的な腸の障害を引き起こす可能性があり、卵の抗酸化物質を含む食品由来の抗酸化物質の腸の健康に対する可能性を調査することに関心が高まっている。鶏卵には、ビタミン、カロテノイド、ミネラル、微量元素に加え、オボトランスフェリンなどの主要な卵白タンパク質 [103,104,105,106] を包含する多くの抗酸化化合物が含まれている。
を含む卵黄タンパク質[113]、オボムコイドおよびオボムコイド加水分解物[111,112]、オボムチン加水分解物および由来ペプチド[112]、およびホスビチンを含む卵黄タンパク質のネイティブフォームまたは加水分解ペプチドとして[98,99,104,105,107,108,109,110]。
これらの分子のほとんどはin vitroで生成されたが、ブタモデルで行われたいくつかのアッセイは、炎症性サイトカインの産生を減少させる卵黄に由来するタンパク質の有益な効果を明らかにした[114]。著者らは、卵黄-タンパク質を食事に補充することは、腸の酸化ストレスを軽減するための新しい戦略である可能性があると結論付けている[114]。
3.3. 抗がん性分子
食品由来のタンパク質やペプチドが、がん疾患の予防や治療にも有益であることを示すデータはごくわずかである[26]。卵白リゾチームの腫瘍抑制活性については、実験腫瘍を用いたいくつかの研究により確認されている。
その効果は、本質的に免疫増強に依存している[115]。オボムシン(βサブユニット)およびオボムシン由来のペプチドも、細胞毒性作用および免疫系の活性化を介して抗腫瘍活性を示した[74]。
卵トリペプチドの抗がん作用[27]やオボトランスフェリン由来の加水分解ペプチド[116]も発表されている。この分野の情報は非常に少ないが、このような活性を調査し続けることは価値があるかもしれない。卵のプロテアーゼ阻害剤に関する研究 [47] からは、エンドウ豆のような豆類を含む他の食品に存在する同様の分子が、大腸化学予防剤として期待されていることから、興味深いデータが得られるかもしれない [117]。
3.4. 免疫調節活性
いくつかの卵タンパク質は、免疫調節活性を持つ可能性がある。中でも卵白リゾチームは、炎症性腸疾患の治療薬として有望視されている。大腸炎ブタモデルにおいて、リゾチームは動物を大腸炎から有意に保護し、炎症性サイトカインの局所発現を減少させる一方で、抗炎症性メディエーターの発現を増加させることが実証された[118]。
オボムシン、カラザイ、卵黄膜のタンパク質分解によって生成される硫酸化糖ペプチドは、in vitroでマクロファージ刺激活性を示すことができる[74]。卵白のプリオトロフィンなどのサイトカインは、炎症反応の生成と解消に極めて重要な役割を担っている。
ヒトでは、プリオトロフィンは、リンパ球の生存を促進し、免疫細胞の化学走性を駆動することが示されている[119,120]。しかし、ヒトの腸における卵白プレオトロフィンの潜在的な免疫調節活性の生物学的意義は、非常に推測されたままである。一方、卵黄ビテロジェニン加水分解物[121,122]は、消化酵素による部分分解の後、何らかの貴重な免疫調節活性を発現する可能性がある。
3.5. 抗高血圧活性
世界的な高血圧の有病率と重要性(12億人以上)を考慮すると[123]、この多因子疾患を制御する方法を見つけるための研究がますます進行している。人口レベルでは、血圧の長期的な制御の最も重要な要因は、ナトリウムおよびカリウムの摂取量と、レニン-アンジオテンシン-アルドステロン系の重要性である。
降圧作用を有する卵由来ペプチドの多くは、アンジオテンシン変換酵素(ACE)に対する阻害活性を示す。この酵素は、アンジオテンシンIから活性型血管収縮物質であるアンジオテンシンIIへの変換を促進する。
卵黄由来のペプチドは、オボトランスフェリンや卵白の加水分解物とともに、降圧活性を有するものがいくつか文献に記載されている [113,124]。これらのペプチドの中には、3つのアミノ酸しか含まないものもある[27,127]。
これらのトリペプチドの中には,生体内で活性を示すものがあり,高血圧ラットに経口投与したところ,血圧を有意に低下させたことから[128],循環器疾患の発生を抑制するのに役立つと考えられている[127,129].
4. 卵の品質に影響を与える要因
4.1. 遺伝学
卵質の選択は、採卵鶏の販売を行う企業の繁殖戦略において重要な要素である。実際、消費者は卵殻の強い高品質な製品を求めており、同時にコストを削減し、微生物汚染物質を排除した卵を保証し、飼育システムの受容性を向上させる[130,131]。
卵の品質を向上させるためのほとんどの選択戦略は、殻の物理的特性(および衝撃に耐える能力)、卵重量の安定性、卵白の品質、および卵黄の割合に焦点を当てている。卵白品質とは、基本的に鮮度を反映する卵白の高さ、および消費者の毒素感染リスク(サルモネラ症)に関連する可能性のある微生物の増殖と生存を防止する能力を指す。
最近、いくつかの著者は、様々な選択系統における卵白の高さ/pHの違いを報告し[132]、特定の形質に関する選択が、卵白の高さを増加させる一方で、卵黄、卵白および殻の割合を変化させたことを裏付けている。卵黄と卵白の重量の中程度の高いばらつきは、選抜系統と伝統的な系統を比較した場合にも観察された[133]。
卵白は、その高い粘性、卵の貯蔵により徐々にアルカリ性になるpH(7.8から9.5)、および無数の抗菌性分子(セクション3.1参照)の存在により、細菌にとって非常に好ましくない培地である。実際、卵白の抗菌力は中程度に継承されることが実証されている[134]。
卵タンパク質とペプチドに関しては、褐色卵と白色卵、あるいは系統の違いによって特定の分子の相対的な存在量に多少の差があることが報告されているが、主要な卵タンパク質は基本的に変化しない[135,136]。
4.2. 栄養と飼育システム
産卵鶏の栄養、飼料の特性(栄養素の構成、エネルギー含量だけでなく、飼料のテクスチャーとプレゼンテーション)、および一日を通しての飼料供給形態は、卵重量だけでなく、卵黄と卵白の割合にも影響する[137]。子豚の飼料の質は、基本的に産卵開始時に卵重量に影響を与えるが、産卵期間全体を考慮すると、その重要性はかなり低くなる [137]。
食事の特性には、カルシウムの供給レベルとその特定のサイズが含まれる。特定の形態の食事性カルシウムによって、鶏は一日の終わりにカルシウムに対する特定の食欲を示し、それが蓄えられ、殻形成が行われる夜間にさらに同化される [138]。
卵重は鶏の一日のエネルギー消費量に影響される。高エネルギーの飼料とリノール酸の供給は卵重を増加させる。この効果は産卵開始時(22-32週)に特に顕著であり、高齢の鶏ではあまり顕著ではない [139]。卵重はまた、食餌タンパク質のレベルによっても増加し、いくつかの研究では、メチオニンが鶏の食餌中の存在と卵重との正の相関があることから、主な制限アミノ酸であることが明らかにされている[140]。
さらに、産卵鶏が伝統的にトウモロコシ、小麦、大豆粕を与えていることを考慮すると、エネルギー摂取量はタンパク質源に依存する。飼料中の抗栄養因子(プロテアーゼ阻害剤およびコンビシリン、グリシニン、クルシフェリンといった消化性プロテアーゼに対して高い耐性を持つタンパク質 [141])の存在は、鶏による飼料全体の消化率およびその後の卵重量に影響を与える可能性がある。
しかし、卵の主要成分の含有量は比較的安定しており、その変動は本質的に、非常に対照的な組成を示す卵黄と卵白の割合に依存する(セクション 2 参照)。一方、卵黄の脂肪酸組成、ビタミンや微量ミネラルなどの微量栄養素(セクション 2.2 参照)、カロテノイドの含有量は非常に変動しやすく、飼料組成に直接依存する [137]。
*
トリグリセリドやリン脂質に含まれる卵の脂肪酸組成は、鶏の飼料に含まれる脂肪酸の組成を直接的に反映している。一方、鶏の飼料に飽和脂肪酸が含まれていても、卵黄の脂質組成にはあまり影響がない。鶏の飼料中の飽和および不飽和脂肪酸含有量は、魚、チア、亜麻仁 [142,143]、オリーブ、または大豆油などの高度な不飽和脂肪酸を示す油または飼料を含めることによって変更できる(レビューについては文献 [144] を参照されたい)。
例えば、鶏の飼料にオリーブ油を含めると、卵黄にモノ不飽和脂肪酸(特に、オレイン酸含有量)を取り込むのに有利であり、一方、土壌油は不飽和n-6脂肪酸(リノール酸)を増加させる [144]。より最近では、微細藻類または亜麻仁を用いた餌の濃縮によって得られた結果から、これらの化合物が卵黄のn-3脂肪酸の含有量を増加させる(それぞれ3倍以上および4倍以上の増加)可能性があることが明らかにされている[145]。
マリーゴールド抽出物粉末[146]、Schizochytrium微細藻類[147]、プレバイオティックとプロバイオティックの組み合わせ[148]などでも、卵黄の多価不飽和含量を増加させる同様の傾向が観察されている。結論として、人間の健康にとって興味深いいくつかの不飽和脂肪酸を卵に濃縮することは比較的容易である。
課題は、卵黄の技術的・感覚的品質に影響を与えることなく、多価不飽和脂肪酸の含有量を増加させる動物性および植物性の供給源を特定し、食品産業(卵製品)の要件と消費者の需要の両方に適合させることにある。
*
卵に含まれるセレン、ヨウ素、そして低いが鉄、亜鉛、フッ化物、マグネシウムなどの微量ミネラルの含有量も、鶏の飼料を増やすことで増やすことができる [149]。平均的なセレン卵の含有量は卵あたり約 5 µg であるが、鶏に 0.3 から 0.5 mg のセレン(セレノメチオニンまたはセレン強化酵母から)/kg を供給すると、3 から 6 倍(卵白で 12 倍、卵黄で 4 倍)になり 30-40µg/ 卵に到達することができる。このような卵の濃縮により、人間の一日所要量の50-70%が供給される[150]。
*
同様に、鶏の栄養は、卵の親油性ビタミン(A、D、E、K)または水溶性ビタミン(葉酸、B12、パントテン酸、および低倍率ではリボフラビン、チアミン、ビオチン)を強化する手段である。ビタミンAは30,000IUのレチノールを与えると10倍、ビタミンD3は15倍(2500IUと15,000IUのD3を与えた鶏で2〜5〜34μg/100g)に増加することが分かっている。
卵黄中のビタミン E 含有量は、基礎飼料の含有量と飼料供給量に応じて 3 ~ 20 倍に増加する。水溶性ビタミンの場合、食餌供給量の増加による増加の程度は低く、葉酸、リボフラビン、コバラミンでは2倍以上、それより低いレベルでは、チアミン、ビオチン、パントテン酸、ピリドキシン、ナイアシン[137]である。
卵黄の色(黄色/オレンジ色の濃淡)も、食事に含まれるカロテノイドの含有量によって決まる[137]。鳥類のカロテノイド(ルテイン、キサントフィル、ゼアキサンチン)の主な供給源は、トウモロコシ、ルツェルン、花(マリーゴールド)、パプリカエキス(赤カロテノイド)で、よりオレンジがかった黄色の黄身に対する消費者の需要に応えるために鶏の飼料に取り入れられている。
卵黄の視覚的側面を高めることへの関心の他に、卵黄中のカロテノイドの高い含有量は、視覚的性能を高め、加齢黄斑変性症のリスクを低減するという人間の健康に対する肯定的な発生を有する可能性がある[151]。
*
自由飼育の鳥類は、基本的な餌に加えて、草、昆虫、ミミズを利用できるため、一部の卵の微量栄養素の含有量もわずかに異なる場合がある。例えば、同じような従来型の飼料を与えた場合、自由飼育では、総トコフェロール量、α-トコフェロール量、ルテイン量が、それぞれバッテリーケージおよび有機システムに比べて有意に高くなる[152]。
逆に,脂質と総ステロール含量には有意な差は認められなかった[152].卵白の高さと卵黄の色の減少は、従来のケージと放し飼いシステムの卵を比較した場合にも観察された[153]。しかし、一般的に、慣行飼育の卵は、化学添加物を含む可能性があるため、カロテノイドやビタミンをより多く含んでおり、そのような行為は有機飼育では行われていないことが分かっている。
これと並行して、放し飼いシステムでは、環境微生物の存在によって鶏の免疫システムがより困難になると考えられるため、卵黄中の免疫グロブリンYの含有量も増加すると考えられる(最初は、乳児に対する母親の初乳と同様に、ひよこに何らかの受動免疫を提供するために)。
さらに、鶏が環境微生物にさらされた場合、卵白の抗菌能力もわずかに変化する可能性があることを示すものもある[154]。結論として、産卵鶏を放し飼いシステムで飼育することで、卵の抗菌力が世界的に向上する可能性がある(セクション3.1参照)。
4.3. 生理的状態
卵の生産と品質は、鶏の年齢、ストレス、免疫状態などの生理学的状態に大きく影響される[155]。卵重は主に鶏の年齢と遺伝によって50gから70gまで変化する(セクション4.1参照)。現代の群れでは、産卵周期(20〜80週齢)を通して卵重は62〜66gに制限されている。
卵重は卵の格付け(小、中、大、特大)に用いられる主要な基準である。高齢の鶏で観察される卵重の増加は、卵白と卵黄の平均重量の増加および卵黄の相対的割合の増加と関連している[156]。鶏の年齢は卵殻強度の低下、卵白高度の低下(卵白高度が高いほど鮮度等級が高い)[153]、卵黄破裂の高い発生率と頻繁に関連している卵膜強度の低下とも関連している[157]。
この後者の観察は、高齢の鶏で卵黄の割合が増加することに起因すると思われる。しかし、卵の化学的組成については、鶏の年齢によって卵の脂肪酸組成(ドコサヘキサエン酸およびアラキドン酸)に若干の違いがあることを示すものもあるが、結果はかなり議論のあるところである[158]。しかし、これらの卵質の加齢による変化は、産卵サイクルの終わりに商業用鶏で起こる生理的変化や一次代謝異常と一致している[159]。
*
病気や感染症は食欲不振や生理的な障害を引き起こし、それによって動物の成長、産卵、卵質(卵殻変形、卵殻欠陥、卵白の減少など)に影響を与えるため、産卵成績も一般鶏の健康状態に左右される。衛生的な卵質に影響を与える産卵鶏で最もよく見られる微生物は、Salmonella Enteritidis、マイコプラズマ、伝染性気管支炎ウイルス、鳥インフルエンザウイルスである[160]。
鶏の福祉と卵の品質に関するもう一つの大きな問題は、システムの種類に関係なく、産卵鶏のほとんどの飼育システムで見られる家禽赤ダニである[161]。採血性であるこのダニは、苦痛、貧血、卵生産量の減少、卵質の低下など、家禽の宿主福祉に劇的な影響を及ぼす[162]。
アカダニの蔓延は、最近の鶏の飼育法の変更(非ケージシステムの支持)、殺ダニ剤耐性の増加、気候温暖化、蔓延を制御する効率的で持続可能なソリューションの不在の結果として、増加すると予想されている [163]。また、サルモネラ菌を含む食品媒介病原体の媒介となる可能性があるため、公衆衛生上も大きな懸念材料となっている[164]。
これらの微生物の中でも、Salmonella Enterica Enteritidisは、卵白の中で生存し [165]、4℃や25℃での数週間の保存後でも [166]、食品媒介性疾患の原因となるため、卵消費者にとって最も危機的な存在と言える。
卵の消費に関連する主要な病原体であることに変わりはないが [167]、卵の生産チェーン全体におけるサルモネラ・エンテリティディスの蔓延を制御するために、当局がかなりの努力を払ってきた。サルモネラの検出とモニタリング [169]、プレハーベスト対策の確立 [170]、管理と衛生対策 [171]、一定の条件下での洗浄による卵の汚染除去 [172] など、養鶏におけるサルモネラ管理プログラムの実施により、報告されたサルモネラ症患者数は減少し続けている [168]。
欧州の法律では、卵の洗浄を認めていない(欧州委員会規則(EC)No 589/2008)。「クチクラなどの物理的障壁を損傷する可能性があり、細菌による経殻汚染や水分損失を助長し、それによって、特にその後の乾燥や保存条件が最適でない場合、消費者へのリスクを高める可能性があるため」であることは注目すべき点である。
4.4. 卵の保管と熱処理
殻付き卵は消費者が使用する前に室温またはできれば冷蔵庫で保管される(卵は産卵後28日までは「新鮮」とみなされる)。卵の貯蔵条件は、食品産業にとって有用ないくつかの技術的特性を高める物理化学的変化や、卵白の抗菌特性の変化など、深い内部変化を引き起こす可能性がある [173,174,175,176] (図 3)。
これらの変化は、卵黄と卵白の間の水交換と、卵殻の孔からの水と二酸化炭素の損失から生じ、2つの卵殻膜の間に発達する気泡の増加を誘発する(図3a)。アルバムの高さは保存期間とともに減少し、一方、アルバムのpHとホイップ量は増加する[136]。
これと並行して、卵の貯蔵中に卵膜が緩んで強度が低下し、卵黄の形状・指数に影響を与える(卵黄は扁平になり、直径が大きくなる)[173]。これらの後者の変化は、炭水化物やグルコース [177]、タンパク質 [178,179,180]、ビタミン、微量元素などの成分の卵白/卵黄交換を促進する [181]。
さらに、保存期間と条件は、タンパク質の分解[175,179,180]とその抗菌力の低下[175]に関連している。しかし、卵白と卵黄を構成する脂質、ビタミン、ミネラルについては、タンパク質を除いて、保存中の変化・変性について記載された情報はほとんどない。
これらの変化が卵黄と卵白のそれぞれの機能的、栄養的、技術的特性(起泡性、乳化性など)にどのように影響するかをさらに調査することは興味深いと思われる。最近のデータでは、卵黄の抗酸化活性は6週間の小売店での保存期間中、ほとんど変化しないことが実証されている[182]。これらの鮮度基準の変化はすべて、冷蔵条件と比較して室温で加速される。
図3 卵の保存に伴う物理化学的変化(産みたて卵と室温で2週間保存した卵の比較)
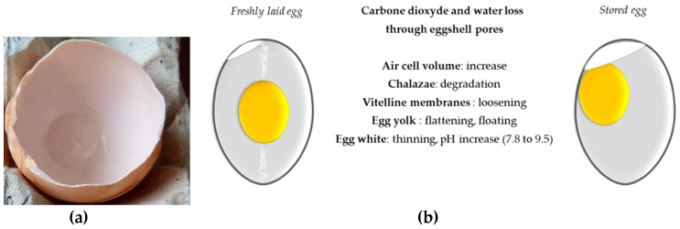
(a)気泡、(b)保存中に発生する主な変化
保存に加えて、卵の栄養素は調理中にも変化することが予想される。生卵、ゆで卵、ゆで卵を比較したところ、ミネラルやビタミンの変性は認められなかった(表5)。
表5 調理によって変化する卵の特性と主成分の一覧表 1.
| 名前 | 卵、丸ごと、生、新鮮 | 卵、丸ごと、ソフトボイルド | 卵、丸ごと、ゆで卵 |
|---|---|---|---|
| エネルギー(kcal / 100g) | 140 1 ; 143 2 | 142; 143 | 134; 155 |
| たんぱく質(g / 100g) | 12.7; 12.56 | 12.2; 12.51 | 13.5; 12.58 |
| 炭水化物(g / 100g) | 0.27; 0.72 | 1.08; 0.71 | 0.52; 1.12 |
| 脂肪(g / 100g) | 9.83; 9.51 | 9.82; 9.47 | 8.62; 10.61 |
| FA飽和(g / 100g) | 2.64; 3.126 | 3.11; 3.11 | 2.55; 3.267 |
| FAモノ不飽和(g / 100g) | 3.66; 3.658 | 4.42; 3.643 | 3.57; 4.077 |
| FA多価不飽和(g / 100g) | 1.65; 1.911 | 1.28; 1.904 | 1.03; 1.414 |
| コレステロール(mg / 100g) | 398; 372 | 222; 370 | 355; 373 |
| 塩(g / 100g) | 0.31 | 0.2 | 0.31 |
| カルシウム(mg / 100g) | 76.8; 56 | 150; 56 | 41; 50 |
| カリウム(mg / 100g) | 134; 138 | 164; 138 | 120; 126 |
| セレン(µg / 100g) | 30 | 23.8 | 7.01 |
| ビタミンA、レチノール(µg / 100g) | 182; 160 | 132; 160 | 61.5; 149 |
| ビタミンD(µg / 100g) | 1.88; 2.0 | 1.28; 2.0 | 1.12; 2.2 |
| ビタミンE(mg / 100g) | 1.43; 1.05 | 2.17; 1.04 | 1.03; 1.03 |
| コリン(mg / 100g) | 250; 293.8 | – | 230; 293.8 |
1 出典 フランス食品・環境・労働安全衛生庁。
ANSES-CIQUAL フランス食品成分表バージョン 2017. Ciqual ホームページ ciqual.anses.fr/ より 2019/01/11 に取得; 2 斜体で、Department of Agriculture, Agricultural Research Service (2014) より 2019/01/18 に取得. USDA National Nutrient Database for Standard Reference, Release 27 and USDA Database for the Choline Content of Common Foods, Release 2 [30]; FA, fatty acid.
実は、参照元によって矛盾しているように見えるデータもある(CIQUAL対USDA、表5)。いずれにせよ、多価不飽和脂肪酸、セレン、ビタミンA [21]の量は、調理によって、特に固ゆで卵では減少する傾向があるようだ(表5)。
*
特に、タンパク質は、その相対的な量は調理によって影響を受けないにもかかわらず、調理によって大きな構造変化を起こしていることがわかる(表5)。このタンパク質の変性は、卵白抗タンパク質酵素のような抗栄養因子を不活性化するのに有効であるが、抵抗性の高いタンパク質を変性させ、消化管でのプロテアーゼ活性を促進させる可能性もある。
また、卵タンパク質の消化率の高さは、小児の卵に対する過敏症の抑制に寄与すると考えられる[183,184]。一方、調理によって、遊離芳香族アミノ酸、ルテイン、ゼアキサンチンに関連する卵黄の酸素ラジカル消去能(抗酸化能)が著しく低下すること[182]、また卵黄の脂質にも影響を与えることが明らかにされている[185]。
これらの観察は、卵の消化率およびそれに関連する栄養および栄養補助食品の品質を評価するためには、食品マトリックスおよび卵の調理方法を考慮することが非常に重要であることを裏付けている[186]。
結論として、多くの分子が調理によって影響を受ける可能性があるため、人間の健康に対する卵の調理の有益性とリスクのバランスを評価することはかなり困難であり、同時に、加熱工程は卵タンパク質の消化率を高め、潜在的に新しい生物活性ペプチドを明らかにするかもしれない[187、188]が、卵の調理によって消費者のトキシ感染の原因となり得る病原体を除去することも言及するに値する。
結論として、これらのデータを考慮すると、卵に関連する栄養学的および栄養補助食品の効果を最大限に維持するためのアドバイスとして、卵白は(抗栄養因子と潜在的な病原菌を不活性化するために)調理され、卵黄は(ほとんどのビタミン、脂質、微量栄養素、いくつかの生物活性(抗酸化)分子を維持するために)基本的に生のままのポーチドエッグまたは半熟卵を消費するようにするとよいだろう。
4.5. 鳥類家畜種間のばらつき
食卓用卵の市場は、どの国でも鶏卵が圧倒的に多い。しかし、アジアの一部の国では、アヒルの卵も広く消費されている。ニワトリは扱いやすく、飼育も簡単で、年間320個近い卵を産むように何十年も前から選抜されてきたのだ。逆に、ガチョウ、七面鳥、アヒルなどは、季節的に産卵するため、衛生面や飼育環境に最も厳しいとされている。鶏の卵も適度な大きさで、大きすぎず、ウズラの卵より大きく、後者はグルメな食材として時々食べられている。
*
伝統的な家畜種の卵の組成は共通の特徴をもっているが [189,190] 、エネルギーの面ではいくつかの大きな違いがあり、それは主に卵黄と卵白の相対的な割合の変化で説明できる(図 4)。鶏、うずら、アヒル、ガチョウ、七面鳥の卵のエネルギー(kcal/100g)は、それぞれ 143, 158, 185, 185, 171 である。
タンパク質の相対量は種間で安定している(約 13%)が、脂質の割合は 9.5% (ニワトリ)から 13% 以上(アヒル、ガチョウ)へと変化しており(図 4、これがそれぞれのエネルギー値の変化の大部分を説明している)、その結果、卵のエネルギー値が大きく変化している。
アヒルやガチョウの卵黄は、鶏卵と比較して相対的に脂肪含量が高く、卵黄の割合が高い[189]。これと並行して、卵黄の脂質プロファイルは、種によっていくつかの特異性を示す[191,192,193]。全体として、アヒルの卵の組成はガチョウの卵の組成に似ており、これは系統学的に近接していることと矛盾しない。
図4 5種の家禽類における卵の組成のばらつき
* *
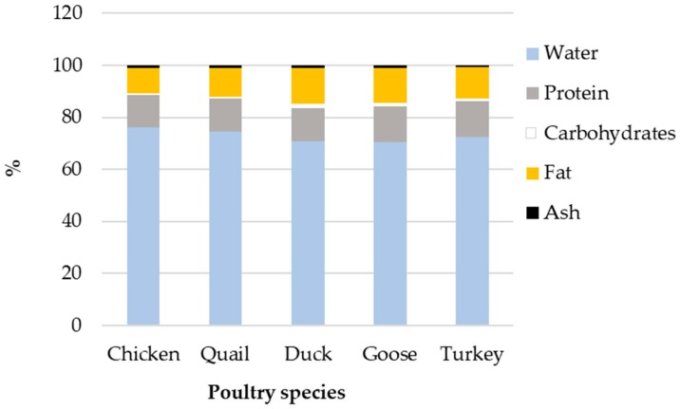
農務省農業研究事業団(2014)より2019/01/17に取得した。
USDA National Nutrient Database for Standard Reference, Release 27.www.ars.usda.gov/ba/bhnrc/ndl/
また、鶏卵のミネラルと微量元素の含有量は、通常、他の種、特にアヒルやガチョウの種で観察されるものより低いことも注目される(表6)。ビタミン類についても同様の傾向が見られる(図5)。ただし、卵に含まれるビタミンや微量元素の変化は、主に飼料組成に依存することに言及しなければならない。したがって、これらの違いは、卵にこれらの化合物を保持する鶏の遺伝的能力よりも、むしろ鳥の飼育条件を反映している可能性がある。
図5 様々な家畜種の卵のビタミンプロファイル
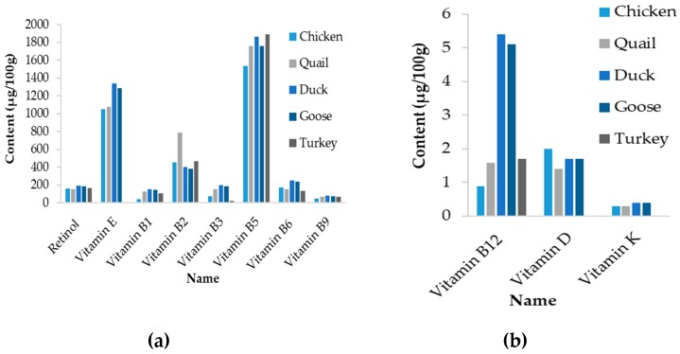
(a)存在量の多いビタミン類、(b)存在量の少ないビタミン類。なお、七面鳥の卵のビタミンE、D、Kの濃度は不明であった。農務省農業研究事業団(2014)より2019/01/17に取得した。
USDA National Nutrient Database for Standard Reference, Release 27.www.ars.usda.gov/ba/bhnrc/ndl/
表6 鶏卵、ウズラ卵、アヒル卵、ガチョウ卵、七面鳥卵に含まれる卵のミネラルと微量元素の比較(平均含有量;mg/100g)
| 名前 | 鶏 | ウズラ | アヒル | ガチョウ | 七面鳥 |
|---|---|---|---|---|---|
| カルシウム | 56 | 64 | 64 | 60 | 99 |
| 銅 | 0.072 | 0.062 | 0.062 | 0.062 | 0.062 |
| 鉄 | 1.75 | 3.65 | 3.85 | 3.64 | 4.1 |
| マグネシウム | 12 | 13 | 17 | 16 | 13 |
| マンガン | 0.028 | 0.038 | 0.038 | 0.038 | 0.038 |
| リン | 198 | 226 | 220 | 208 | 170 |
| カリウム | 138 | 132 | 222 | 210 | 142 |
| セレン | 0.0307 | 0.032 | 0.0364 | 0.0369 | 0.0343 |
| ナトリウム | 142 | 141 | 146 | 138 | 151 |
| 亜鉛 | 1.29 | 1.47 | 1.41 | 1.33 | 1.58 |
農務省農業研究事業団(2014)より2019/01/17に取得した。
USDA National Nutrient Database for Standard Reference, Release 27.www.ars.usda.gov/ba/bhnrc/ndl
これらの化学化合物以外に、生理活性分子の変動も報告されている。実際,卵白と卵黄のプロテオームの比較解析により,どちらかの種に特異的に関連するタンパク質[194,195,196,197]と,存在量に差を示すタンパク質(オボトランスフェリン,リゾチーム,ガリン)[196]があることがわかった。
これらの特徴は、鳥類の由来によって卵白抽出物の総合的な生理活性(抗菌活性および/または抗酸化活性)に影響を与えると考えられ、少なくとも一部の細菌株(Bacillus subtilis、Staphylococcus aureus、Pseudomonas aeruginosa、Escherichia coli)に対して、鶏卵由来の卵白は七面鳥、鴨、雁由来の卵白と比べて最高の抗菌力を保持することが示された[91]。さらに、アヒル、ガチョウ、七面鳥の卵白に含まれるオボムコイド(化学変性や熱変性に強いプロテアーゼ阻害剤)の存在量が少ないこと [196] は、タンパク質消化率の高さに関連している可能性がある。
*
結論として、卵の化学組成におけるこのような違い、さらにその生物学的活性分子のいくつかにおける違いは、鶏卵と比較して、他の種からのいくつかの卵タンパク質の栄養補助食品的価値の増加または減少と相関している可能性が高い。
5.結論
古くから、卵は人間にとって栄養価の高い食品とされ、世界中で広く消費されている。その消費量は、肉を使わない食事(ベジタリアン)を始めたり、肉の摂取量を大幅に減らすオクシデントの消費者が増えていることを考慮すると、今後も継続的に増加すると予測される。
我々の消費様式と食習慣のこの変化は、肉類摂取と消化器系癌および心臓血管系疾患との関連リスクに関する多くのデータ、および菜食主義を賞賛する研究の増加によって動機づけられている[198,199,200,201]。並行して、食肉生産様式に関する倫理的な懸念や環境問題にも後押しされている [202]。
また、卵の消費量には国によってかなりの格差があり [18]、特に中央アフリカでは一人当たりわずか36卵/年という低さであることも強調されている [19]。発展途上国における卵産業の発展は、人間の栄養・健康や経済にとって大きなチャンスとなるかもしれない。
*
基本的な栄養素の他に、卵は潜在的な栄養補助食品の大きな供給源でもある。卵白および卵黄/ビテリン膜には、これまでに合計550種類のタンパク質が同定されており、そのうち生理学的な機能は、現在までにわずか20種類しか明らかにされていない。
このことは、消化管における卵タンパク質の動態を評価する研究が行われていない現状を考慮すると、卵にはまだ多くの未知の活性が存在し、さらなる研究に値することを示唆している。このような研究は、卵タンパク質の生体内での可能性と、その結果として生じる加水分解ペプチドをより良く理解するのに役立ち、食品および製薬に基づく研究において他の食品で使用されてきた動的胃モデルを用いて容易に理解することができるだろう[203,204]。
これらのin vitroモデルは、胃の消化の生化学的および機械的側面を模倣している。人工唾液、食品を崩壊させる圧縮力、連続的な胃排出のシミュレーション、ヒトの胃と同様のpHプロファイルを生成する胃液分泌を組み込んでいる。また,胆汁酸塩と腸内酵素を含み,現実的な時間依存性で順次作用し [205] ,腸内細菌叢を追加することによって改善される可能性がある。
このモデルは,栄養素の生物学的アクセス性を評価したり,食品マトリックスの構造変化を研究するために,すでに幅広い研究で利用されている。このような実験戦略は、食事中の卵の調理法(生か加熱か)が生理活性ペプチドの生成に与える影響を研究し、ヒトの健康に対するその生物学的意義をより良く理解するための有望な方法となると期待される。
腸の健康は、宿主のゲノム、栄養、ライフスタイルの相互作用に依存し、正常な脳機能や精神的健康に寄与していることは、現在ではよく知られている[206]。
資金提供
本研究は、外部からの資金援助を受けていない。
利益相反
著者は利益相反のないことを宣言する。