

Contents
The genetic and cultural evolution of unsustainability
pubmed.ncbi.nlm.nih.gov/32292525/
link.springer.com/article/10.1007/s11625-020-00803-z
概要
人為的な変化は加速度的に進行し、地球上の生物の将来を脅かしている。これらの人為的変化の近接メカニズムはよく研究されているが(気候変動、生物多様性の損失、人口増加など)、進化的な因果関係はほとんど無視されてきた。
Anthroecological theory(AET)「人類生態学的理論/アンソロエコロジー理論」は、人為的な環境変化の究極の原因は、ニッチ構築と生態系エンジニアリングのためのマルチレベルの選択であると提唱している。
ここでは、この理論をロトカの最大電力原理と統合し、環境からのエネルギー抽出と遺伝的・技術的・文化的進化を結びつけ、人類の生態系収容力を高めるモデルを提唱する。
環境収容力は、集団が環境から獲得できる正味のエネルギーや、エネルギー入力から子孫の出力への変換効率といったエネルギー的な要因によって部分的に決定される。これらの要因は、すべての種においてダーウィンの遺伝的淘汰の下にあるが、人間においては、技術や文化によっても決定される。
もし個人や社会集団の環境収容力を高める能力に遺伝的または非遺伝的な変異があれば、環境収容力を高めるために淘汰や文化的進化が作用すると仮定する。さらに、この環境収容力の進化が、生態系の生物学的構成要素が自らの進化によって対応できるよりも速く起こる場合、持続不可能な生態系の変化が生じるという仮説を立てている。
はじめに
新石器時代の人口推移以来、人類社会はしばしば図1に描かれているように、持続不可能な連続体に沿って生きてきた(Ponting)2007;Gowdy and Krall2013)。脚注1この連続体の最も端にあるのが社会生態学的「崩壊」で、ここでは生態的要因と社会的要因の組み合わせにより社会の人口および/または発展が急速に低下することと定義されている(Cumming and Peterson2017)。社会生態学的崩壊の全体または一部を経験したと思われる文明には、マヤ人、ローマ人、ギリシャ人、イースター島とピトケアン島の文明、グリーンランドの北欧植民地、アメリカ中西部のカホキア、アナサジ人などがある(Lopinot and Woods)1993;Ponting2007;Hughes2011;Kennett and Beach2013)。これらのケースでは、人間システムは環境からエネルギーや物質を吸収する能力を超えて拡大し、こうした資源の制約が社会・政治的な対立と相互作用して、社会生態学的な崩壊を引き起こした可能性がある。このようなケースやその他のケースにおける正確なメカニズムや社会的要因と生態学的要因の相対的な比重については議論がなされており(Butzer)2012;Butzer and Endfield2012;Boersema2015;Cumming and Peterson2017)、私たちはすべての社会崩壊のケースが主に資源主導であると示唆するものでない。むしろ、私たちは単に、人間集団が繰り返し崩壊を経験しており、これらのケースの多くで、環境と資源の制約が関与していることを指摘するのみである。
図1
持続可能性と人間の非持続可能性の連続体
このような永続的な崩壊は特に顕著であるが、人類はその歴史の中で社会生態学的危機の状態も多く過ごしてきた。私たちは社会生態学的危機を、人間集団が環境からエネルギーや物質を収集する能力に近い、あるいはそれを超えて拡大し、集団とその生活水準を危機に陥れる状況と定義している。社会生態学的危機は崩壊につながる可能性もあるが、社会が回復する可能性もある。社会生態学的危機の例としては、紀元前108年から紀元後1910年までの1828年間(2000年のうち)、中国の少なくとも一つの省で飢饉が発生したこと、1315年から17年のヨーロッパの飢饉で住民が共食いに走ったこと、1696年から97年のフィンランドの飢饉で人口の4分の1が死亡したことが挙げられる(Ponting)2007)。いずれの場合も、危機が崩壊につながったわけではないが、私たちの目的からすれば、記録された人類の歴史の多くが危機的な状態で費やされてきたことだけが重要なのである。
社会生態学的危機や社会生態学的崩壊の状態に加え、人類は旧石器時代の残りの歴史の大半を非持続可能な状態で過ごしてきた。私たちは、非持続可能性を、人間が環境からエネルギーと物質をどんどん取り出して、それを維持と成長のために使う成長段階と定義している。非持続可能性は、経済成長(Daly)1991)と人口増加(Ehrlich and Ehrlich)2009)の両方に関連しており、社会生態学的危機につながる可能性がある。古代ギリシャ文明(Hughes)2011)や現代の西洋文明の成長段階は、非持続可能な状態で存在していると考えることができるかもしれない。新石器革命以降の人類文明は、なぜ社会生態学的危機と崩壊の時期を挟みながら非持続可能な時期が続いているのだろうか。本論文の目的は、なぜ人類が環境の非持続可能性、社会生態学的危機、そして崩壊に至る素因を持っているのかを理解するための概念モデルを提供することである。
人類生態学的理論(AET)は、人間の社会的・文化的進化が、現在地球システムにダメージを与えている生態学的危機の究極の原因であるという仮説を立てている(Ellis)2015;Ellis et al.2018 )。AETによれば、地球システムに対する人間の被害は、マルチレベルの集団選択プロセスを経て進化した社会文化的ニッチ構築の結果である。この理解では、人間は少なくとも農業革命以来、そしておそらく狩猟のために古代のサバンナを焼き払ったときから、生態系エンジニアおよびニッチ構築者として行動してきたと言える。この生態系エンジニアリングによって、「社会的規模」と「人口規模」が徐々に大きくなり、生態系エンジニアリングのレベルがますます高くなることが要求されるようになった。
ここでは、AETとロトカの最大電力原理(Lotka 1922)および進化生態学の概念との融合を試みる。ロトカは、生物が環境からエネルギーを取り出す速度を最大化するように自然選択が働くと主張し、後にH.T.オダムはこの仮説を最大電力原理と名付けた(Sciubba 2011)。ロトカが正しいとすれば、AETが説明する生態系エンジニアリングやニッチ構築は、自然淘汰を通じて作用する最大出力原理の結果として理解することができる。このエネルギー的な見方は、EllisのAETモデルと似ているが、私たちはエネルギー抽出を淘汰の対象として、また環境負荷の指標として用いている。この形式化により、AETの新しいテストとモデルが可能になるかもしれない。本論文では、まず、非持続可能性の進化に関する文献をレビューし、農学、進化心理学、AETからの先行アプローチを紹介する。次に、私たちのエネルギー的AETの概念モデルと環境収容力の進化との関係を説明する。最後に、エネルギッシュなAETが社会学における2つの現代的な問題の理解に与える影響について議論し、結論でこの論文を終える。
非持続可能性の進化に関する先行文献
農業、エネルギー、社会生態学的危機
本作と同様に、Gowdy and Krall(2013,2014,2014,139)人類の非持続可能性の原因を理解しようと試みた。彼らは、人間の超社会性(社会生物学的文献ではeusocialityと呼ばれる)が新石器時代の人口推移の結果として進化し、この超社会性が人新世の環境危機の究極の原因であると主張したのである。ガウディとクラールは、人間集団の超社会性が、選択の主体を個人レベルから集団レベルへ移行させることを可能にしたという。そのため、「農耕への移行に伴い、適応単位としての集団は、余剰生産という命令によって全く異なるゲシュタルトを構成するようになる(Gowdy and Krall2014, 139)」のである。この余剰生産への命令は、Gowdy and Krallのモデルでは環境危機につながるが、同じ表現型は、我々のモデルではLotkaの最大電力原理の表現として理解することも可能であろう。
もちろん、農業と社会生態学的危機を結びつけたのは、ガウディとクラールが初めてではない。トーマス・マルサスの農業と人口に関する古典的な著作とそれに対する反応は、私たちの知る限り、人間社会のエネルギー的な持続可能性に関する最初の研究である。1798年の『人口原理に関する試論』において、マルサスは、人口が指数関数的に増加し、農業収量の向上はせいぜい直線的に増加することを観察した(Malthus and Pullen)1989)。その結果、人間の食糧エネルギー需要が環境から食糧を取り出す能力を上回り、その結果、人口規模、生活水準、あるいはその両方が減少するという、マルサス的破局と呼ばれる事態が発生したのである。マルサスは、環境からのエネルギー抽出の増加率は、主として(技術よりも)資源に制限されており、人口増加率がエネルギー抽出の増加率を上回ると、社会・環境への危機をもたらすと主張した。
マルサスの考えは、18世紀以降、何度も異議を唱えられたが、最も有名なのはEsther Boserup(2014)であろう。Boserupは、農業生産性は主に人間が農業に捧げる技術や労働力によって制限されると主張した。彼女の考えでは、人口密度の上昇に伴い、農業のやり方は「農業集約化」によって変化し、労働力の増加の代償として収量を増加させることになるという。このような変化は、本稿で言うところの技術的・文化的進化に相当し、環境からのエネルギー抽出を増大させる。このエネルギー抽出の増大は、マルサスとは逆に、指数関数的に増加する人口を支えることができる。
Boserupの研究以来、緑の革命は、新しい作物品種と天然ガスから作られる肥料によって、主食用作物の生産性を向上させた。この革命は、20世紀後半の発展途上国における人口の急増に伴う社会生態学的危機を改善することができた(Ehrlich and Ehrlich 2009)。このように、Boserupの論文の概要と一致しているのは、人類はテクノロジーを使って環境からのエネルギー抽出を増やすことができ、このテクノロジーを介したエネルギー抽出が人口規模の増加を支え、農業におけるテクノロジー利用の増加を可能にする文化の変化(進化)に相応して発生したということです(Possas et al. 1996)。この過程は、ここで展開されたエネルギー的AET仮説と一致する。
BoserupとMalthusは、人間の人口における農業の役割を理解するための重要かつ対照的なレンズを提供し、GowdyとKrallは農業を人間の進化における重要な転換点として特定しているが、農業はもはや人間にとって優勢なエネルギー源ではない(Smil 2017)。そこで、私たちは、農業システムからのこれらの洞察と一致しながらも、より進化的でより一般的な、人間が使用するが人間の直接的な代謝に使用されないエネルギーと定義される体外エネルギーを取り入れた、人間社会生態系の概念的進化モデルの構築を目指す(Price 1995)。さらに、社会生態学的システムのエネルギー論に関する先行モデルに欠けている正帰還を提案し、ラチェット効果も統合して、社会生態学的危機への人類の進化を一方向的なものにする。
進化心理学と社会生態学的危機
Gowdy and Krall(2013,2014)が人間の非持続可能性を理解する鍵は農業であると考えたのに対し、Van Vugt et al.(2014)は非持続可能性のルーツを農業が発明される前の旧石器時代に見出した。Van Vugtらは、進化心理学的なアプローチを用いて、旧石器時代の人間の祖先淘汰が、人間を持続可能な暮らしに向かわせない心理状況を作り出したと主張した。例えば、財の絶対量よりも地位を重視する相対的価値観は、先史時代には適応的であったが、現代社会では持続可能性に不適応になっていると論じている。本論文では、Van Vugtら(2014)が述べた特徴を、文化進化によって利用される前適応と理解する。
人類生態学的理論とエネルギー
AETは、人間はその生物学的・文化的進化を通じて非持続可能性の素因を持っていると提唱している。具体的には、AETは、人間の非持続可能性は、ゲノムに作用するマルチレベルの選択プロセス(マルチレベル選択の関連概要はWaringら(2015参照)を経て進化し、文化や技術に作用する選択的・非選択的メカニズムと協調して発生すると提唱している。AETでは、このプロセスの結果、ニッチ構築と生態系エンジニアリングを行いやすい種が生まれ、これらのプロセスの規模は、個体数の増加とともに増加し続けている。この規模の増大は、人間のニッチ構築の傾向と相まって、人間の非持続可能性につながる(Ellis 2015;Ellis et al.)
ここでは、人間は環境からのエネルギー抽出率を高めることで、環境収容力(規模)を高めるように進化してきたという仮説を立てている。この仮説はAETの仮説と同じだが、メカニズム(ニッチ構築や生態系工学)ではなく、プロセスの原因(エネルギー抽出の最大化)に重点を置いている点が異なる。つまり、ニッチ構築と生態系エンジニアリングは、人間が環境からのエネルギー抽出を最大化するための2つの可能なメカニズムである。この進化の基盤は、遺伝的なものだけでなく、文化的・技術的なものであるため、生態系が進化的に対応する能力を上回っている。このように、人間と人間以外の生態系は赤の女王のような共進化を遂げているが(Van Valen 1973;Stenseth and Smith1984)、人間の技術は生物的生態系の能力よりも速く進化している。
赤の女王仮説は、ある種が他の種または種群からの選択圧に負けないように共進化し、共進化相手(例えば、病原体とその宿主)の進化速度に差がある共進化システムを説明するためにしばしば引用される。ここでは、レッドクイーンを適応して、ある種(ヒト)と生態系の生物学的構成要素(生物圏)の共進化を含めることにする。したがって、人間の人口が増加するにつれて、人間集団は環境からより多くのエネルギーを要求するようになると仮定する。なぜなら、エネルギー資源は有限であり、熱力学の第2法則により、エネルギー資源はゼロサムであるため、つまり、人間が環境からエネルギーを消費すれば、他の生物が利用できるエネルギーは必然的に少なくなるため、生物環境に選択的圧力をかけることになるからだ。しかし、人類はテクノカルチャーの進化によって進化しており、生物環境が遺伝子の進化によって進化するよりもはるかに速いスピードで進化している。したがって、進化の速い寄生虫(人間)と進化の遅い宿主(生物環境)のアナロジーは適切である。環境そのものが人間の技術・文化の進化と共進化するのではなく、生態系を構成する生物集団が進化し、その結果、生物環境に影響を与えるということを意味していることに注意してほしい(ただし、Lovelock and Margulis1974,Levin1998,Swenson et al2000を参照)。
AETと本仮説の概念の違いはわずかであるが、その意味するところは重要な違いがある。エリスのAETは、人類は生態系エンジニアでありニッチ構築の専門家であるため、やや特殊であると提唱している。一方、本仮説は、その逆の仮説を提唱している。人間は、他のすべての種と同様に、環境からのエネルギー抽出を最大化する。他のすべての種と同様に、ヒトはこのプロセスにおいて、生態系の生物学的構成要素における共進化的反応によって対抗しているのである。人間は、エネルギー抽出における文化的進化の速度が、生物学的環境の共進化的反応を上回っているため、唯一ユニークな存在なのである。この見解では、人類の非持続可能性は、人類独自の高度な文化と、エネルギーの最大化を目指す非独自的な衝動が結びついた結果であり、同様の文化的進化を遂げた種は、同様の行動をとるであろうということだ。ニッチ構築と生態系工学(Ellis 2015)または農業(Gowdy and Krall 2014)は、単に私たちが環境からのエネルギー抽出を最大化するための手段である。
アンサステナビリティの概念モデル
エネルギーと選択
自然淘汰は、ある個体が次の世代に残す子孫の数(=適応度)を最大化するように働く。しかし、エネルギーと適応度は密接に関連している。適応度は、環境からエネルギーを取り込み、子孫に変換する生理的プロセスの結果だからだ。つまり、集団あるいは個体レベルでの成長と生殖には、環境からのエネルギー入力と、環境に対する高エントロピーの廃棄物出力が必要である。このような体力に関する生理学的な理解は、人間以外でもよく研究されており、生活史理論の多くの基礎となっている〔例えば、(Weiner)1992,Garland Jr and Carter1994,Ricklefs and Wikelski2002)〕。この文脈で、Pianka(1970)は、「…自然選択は通常、単位時間あたりに集まる物質とエネルギーの量を最大化するように作用する」と主張した。Brownら(1993)も同様に、フィットネスを「繁殖力、すなわちエネルギーを子孫に変換する率」とするエネルギー的な定義を提示した。この繁殖力は、環境からのエネルギー同化率と子孫へのエネルギー変換率の両方の関数であるとされた(ただし、(Kozlowski)1996)参照)。
Brownらの定義によれば、個体iの体力(F)は、環境から抽出したエネルギー(X)と子孫へのエネルギー変換効率(E)の関数である。
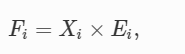 (1)
(1)
ここで、Fは一生涯の子孫数、Xはその個体が一生涯に摂取するカロリー、Eはカロリーあたりの子孫数である。このモデルでは、体力は個体レベルのものであるが、Xや Eに遺伝的な集団変異があるとして、集団レベルでも同じ論理が成り立つ。人間以外の生物が使うエネルギー源は体細胞だけなので、この体力に関する生理学的理解は、人間以外に適用してもトートロジーである。例えば、ライオンに適用すると、式(1)は、ライオンの子孫数は、食べたカロリーの総量に子孫数をかけたものを、食べたカロリーで割ったものであることを論証しているに過ぎない。しかし、遺伝的変異があれば、Xも Eも独立して選択される可能性がある。つまり、ライオンはより優れた採食能力(環境からの総エネルギー獲得量の増加)、またはより効率的な採食、消化、代謝など(効率の向上)を選択される可能性がある。ヒトは非ヒトと異なり、体外エネルギー源(化石燃料など)を利用することができ、そのエネルギー源を遺伝的適合性に変換することができる。例えば、ヒトはサバンナを燃やして獲物を収穫したり(Scherjon et al.)2015)、森林を燃やしてテラプレタを作ったり(Glaser et al.)2001)、化石燃料を改質して肥料を作ったりできるが、これらのエネルギー利用は全て非栄養的エネルギーを用いて間接的にフィットネスを向上させるものである。したがって、ヒトの場合、Xと Eは、ヒト以外の集団のように光合成と呼吸によって制限されることはない。
私たちは、人類はXと Eを増加させるように選択されてきたと仮定する。これは、高いXを好む遺伝的・行動的形質に作用する個人選択と、遺伝子、文化、技術により、環境からより迅速にエネルギーを吸収し、そのエネルギーを適応力に変換する集団を好む集団選択の組み合わせにより生じたのではないかと考えている。このような選択が、体細胞X(食料など)と体細胞外X(化石燃料など)の両方を増加させるように作用してきたと仮定する。私たちは、人類はEよりも Xを最大化するよう優先的に選択されてきたこと、これが社会生態学的危機への偏向の原因であり、このXへの優先的選択はEの変化に対する熱力学的限界のために起こると仮定する。さらに、資源効率向上の選択でさえ、環境への影響を増大させる可能性がある。資源がより効率的にフィットネスに変換されるようになると、人口規模が増大し、その結果、資源の絶対的な使用量が増加することになる。これは、ジェボンズのパラドックスや定常経済学の考え方に類似しており、エネルギー効率の向上でさえ、経済成長の増大を通じて環境への影響を増大させる可能性があることを示唆している(Daly and Cobb Jr)1994)。
個体レベルでは、人間の社会的行動において、エネルギー抽出を最大化するための選択の影響を見ることができる(Penn)2003)。環境からより多くのエネルギーを蓄積し、物質的な所有物の形で社会的に示すと、集団内での地位が向上し、おそらく交尾の成功率が高まる(Kruger)2008;Sundie et al.2011)。これは、エネルギー獲得の間接的な物質的指標(例えば、物)にも、エネルギー獲得の直接的な物理的指標(例えば、男性の魅力の指標としての体格(Swami and Tovée)2005)にも当てはまるかもしれない。もちろん、物質的な所有物や地位の総体的な蓄積には、環境コストがかかる。私たちは、ステータスをもたらすもの(車、家、宝石、服など)を作り、手に入れるためにエネルギーと材料を使う。この資源の使用は、環境に対して高いエントロピーを持つ廃棄物を放出する結果となる。したがって、社会生態学的危機を理解する一つの方法は、個人レベルで働く祖先の選択の結果であり、体現されたエネルギーの蓄積を通じて表される社会的優位性を支持することである(Odum1996)。この物質的・身体的エネルギー蓄積への適応は、環境抽出への事前適応であり、後の文化進化によって利用される(Van Vugt et al. 2014)。
また、遺伝的選択は集団レベルでも働くかもしれない(Wilson and Sober 1994)。この場合、より効率的に資源を蓄積できる個体の集団は、できない、あるいはしない集団と競争することになる。資源の抽出効率や資源の利用効率に遺伝的(遺伝的)変異があれば、これらの形質が選択されることになる(次節で非遺伝的メカニズムを検討する)。式1のように、集団の適応度は、集団が環境からエネルギーを抽出する能力と集団の変換効率の関数である。例えば、社会的知性の遺伝子変異により、より効率的に狩りをする狩猟採集民の集団は、たとえその形質が個人レベルではコストを示すとしても、そのような形質を持たない集団よりも選択されるかもしれない。遺伝的集団選択の可能性と限界についての議論はHenrich(2004)を参照されたい。
環境から集団や個体へのエネルギーの抽出量(X)と、抽出されたエネルギーを子孫に変換する効率(E)を最大化するために社会組織は存在するのである。これは、エネルギー的なフィットネスの定義があれば、社会組織は集団のフィットネス(Wilson and Sober)1994)や集団を構成する個体のフィットネス(Nowak et al.)2010)を最大化するために存在するという主張と同じである。ユニークなのは、その意味合いである。もし社会組織が集団の適応度(または集団のメンバー個人の適応度)を最大化するように働き、適応度が環境からのエネルギー抽出に依存するならば、社会組織は環境からのエネルギー抽出を増大させる結果となる。ここでもまた、Xか Eのどちらかの増加が環境の非持続可能性につながり、したがって、社会組織は社会生態学的危機の進化的原因であると言えるのである。
文化・技術の共進化
マルチレベル選択理論は、自然界で観察される表現型、特に社会的表現型は、個体とより高いレベルの社会組織の両方に同時に作用する選択過程の結果として理解することができると主張している。マルチレベル選択理論では、集団レベルと個体レベルの分散と選択圧のバランスが、結果として生じる遺伝子型と表現型を決定するのである。マルチレベル選択は、生物学では昆虫の社会行動を説明する手段として、Nowakら(2010)、社会科学では親環境行動を含む人間の社会行動を説明する手段として(Bellら)2009、Zefferman and Mathew2015、Richersonら2009,Safarzyńska and van den Bergh2010,Safarzyńska et al.2012,Waring et al.2017,Brooks et al.2018).」
多段階選択説の伝統的な弱点の一つは、祖先の人類集団が集団レベルの選択に必要な集団間の遺伝的分散の維持に適していないように見えたことである(Henrich)2004)。しかし、淘汰は遺伝子の伝達だけにとどまらない。文化的集団選択では、遺伝子ではなく文化が遺伝的変異の源泉となり、ヒトの集団レベルで選択が働くと仮定している。文化継承と遺伝継承では伝達性や分散の発生に違いがあるため、集団間での文化的変異の維持はより妥当であり(Henrich)2004;Bell et al.2009;Waring et al.2015)、人間の形質を説明する手段として集団レベルの文化選択が魅力的である。
表1は、人間の非持続可能性を生み出すために作用する選択のレベルと、その作用の速さを示している。選択の強さは、形質における遺伝的変異と、その形質と適応度との関連性の関数である(Henrich)2004;Freeman and Herron2007)。選択の速度は、その強さと世代時間の関数である(Van Valen)1973)。文化的集団の選択は、文化的変化が表現型の比率を変えるために生物学的生殖を待つ必要がなく、非ランダムな誘導突然変異によって新しい文化的革新を集団に加えることができるため、遺伝的形態の選択よりも速くなることがある(Pereault)2012;Brooks et al.2018)。
表1 energetic-AETで動作する多値選択機能
Xを増加させる行動的・心理的形質に対する遺伝的・個人レベルの選択が、遺伝子-文化共進化(Feldman and Laland)1996;Ambrose2010)の一種の事前適応をもたらし、Xをさらに増加させる技術をもたらすと仮定する。技術-文化進化は文化集団選択(または他の非遺伝的伝達手段)により広がる(Cavalli-Sforza and Feldman1981;Mesoudi et al.2004;Claidière et al.2014;Jordan2014).その結果、技術は、自然システムが自然淘汰によって対応できるよりもはるかに速く変化する可能性がある(Wilson)2012)。技術や技術的文化を持たない種では、Xと Eの変化は、進化的時間に、あらゆる進化に対応して進化する生物学的システム内で起こるだろう。つまり、文化や技術がない場合、システムの生物学的構成要素はすべて、ある種の変化速度が別の種に適応できるような、時間の経過に伴う遺伝子頻度の緩やかな変化に限定される。人間の場合、文化の進化が速すぎたために、他の種が対応できなかったのかもしれない(Perreault)2012)。したがって、私たちの社会生態学的危機を理解するもう一つの方法は、遺伝的でない急速なプロセスによってKを増加させる私たちの能力の結果であると言える。この技術の急速な増加は、人間以外のシステムの生物学的構成要素が共進化的に対応するよりも速く、人間のシステムが変化するという持続不可能な状況を生み出している。
遺伝子の進化(遺伝子頻度の経時変化)になぞらえて、技術文化の進化とは、ある技術の使用頻度の経時変化であると考えることができる。この頻度は極めて急速に変化することがある。例えば、20世紀初頭、1900年に米国で登録された1万台の自動車の使用頻度は、20年後には1000万台まで増加した(Nakicenovic)1986)。私たちは、自動車を多用した社会は、そうでない社会よりもこの時期に大きな経済成長と人口増加を経験し、環境からより多くの資源を取り出し、環境収容力を高めたのではないかと仮定している。このように、20世紀初頭、米国は技術の変化を通じてXを増加させ、この変化は非人間系が共進化を通じて対応できるよりもはるかに急速に起こったのである。
20世紀初頭の米国では、この技術的な媒介によるXの増加が、米国内での生存と生殖を高め、結果として人口増加をもたらした。しかし、人口が増え、豊かになるにつれて、より多くの人口とそのエネルギー的(経済的など)な期待を支えるための新しいエネルギー源が必要とされた。20世紀には、Xと Eの積を維持するという課題があったが、米国はそれを達成した。しかし、20世紀が進むにつれ、人口が増加し、Xと Eの増加に専念できる頭脳の数が増え、技術革新の速度が上がり、エネルギー抽出の速度が上がり、人口を支える社会の能力が上がった。この正のフィードバック・ループが、エネルギー的AET仮説の核心である(図)2)。
図2
Hypothetical positive feedback mechanism in energetic AET which leads to unsustainability
最大出力の原理
Xを増大させるための遺伝的・文化的進化の結果が、ロトカの最大電力原理(Lotka)1921,1922)である。最大出力原理とは、開放系は環境から有用なエネルギーを吸収する速度を最大化する傾向があるというものである。システムは必ずしもエネルギー獲得量を最大化するわけでもなく、エネルギー損失量を最小化するわけでもないが、その代わりに、システムは環境から吸収する使用可能な電力(エネルギーを時間で割ったもの)を最大化する(Hall)2004)。これは、前節で述べた議論と同様で、Xはレート(寿命あたりのエネルギー)であるからだ。したがって、進化がXを最大化するように働くということは、選択が環境からのパワーを最大化するように働くということであり、ロトカの原理と矛盾しない。なお、OdumとPinkerton(Odum and Pinkerton)1955;Odum2007)はLotkaの研究を拡張し、彼らのモデルは効率についてここで紹介したもの(Hall et al.)1986)とは異なる結論を導くかもしれない。ここでは、Odumではなく、Lotkaの感覚に基づく最大出力原理を使用する。
ロトカ(1922)はこう書いている。
もし、生物のシステム全体が実際に利用しているエネルギーを超える利用可能なエネルギーを供給できる源が提示されれば、適切に構成された生物に、システムを通るエネルギーの総流量を増大させる機会が与えられる。…考えられるあらゆる事例において、自然選択は、物質と利用可能なエネルギーの未利用残滓が提示される限り、有機システムの総質量を増大させ、システム内の物質の循環速度を高め、システム内の総エネルギー流量を増大するように作用するだろう」(147-148)
そこでロトカは、環境からエクセルギーXを吸収し、N個のサブシステムを維持するシステムを想定している。人類は技術によってXを繰り返し増加させることができ、その結果Nを増加させてきた。ロトカによれば、「未利用の物質と利用可能なエネルギーが存在する限り」そうし続けることが期待されている。持続不可能性が生じるのは、人類が未利用の物質とエネルギーの新しい供給源を見つけるために、ますます洗練された手段を用いるように進化してきたためだ。最大電力原理の実証的な検証は限られているが、批判(Månsson and McGlade)1993)と同様に現存する(Hall)2004;Cai et al.2006;DeLong2008;Li et al.2013 )。
最大出力原理の最も関連性の高い例は、おそらく新石器時代の革命からきている。Bowles(2011)は、狩猟採集民と初期の農民のエネルギー効率を比較し、単位労働量あたりに抽出されるエネルギーの効率は、農民よりも狩猟採集民の方が高いことを明らかにした(Ponting)2007;Smil2017も参照のこと)。このことから、Bowlesは非エネルギー的な要因が農耕への移行を説明すると結論づけた。しかし、ロトカによれば、自然淘汰は最大効率(単位投入量あたりのエネルギー)よりも最大出力(単位時間あたりのエネルギー)を好むという。初期の農民は、狩猟採集民よりも単位時間あたりに環境からより多くのエネルギーを取り出すことができ、これが新石器革命の成功を説明する。つまり、ある年(時間)において、農耕社会は狩猟採集社会よりも多くのエネルギーを取り出すことができ(Ponting)2007;Smil2017)、このエネルギーを体力に変換することができたのである。したがって、農耕への移行は、労働効率やエネルギー効率に基づく説明とは矛盾するものの、最大出力原理とは一致する。
ロトカは、自然淘汰がシステムを通じて電力の流れを最大化するように作用するからといって、電力の流れを増大させる表現型の分散がないために、必ずしも進化が追随しないことを規定し、この分散を「生成的影響力」と呼んでいる。私たちは、人間の技術と文化が、人間が最大電力原理に従うことを可能にする主要な生成的影響であることを提案する。
人間以外の生物種では、代謝系は種を超えて高度に保存されており、多様な分類群に同じ分子経路が存在している。例えば、呼吸は植物、菌類、動物で類似している。この保存性は、呼吸効率に世代間の影響(分散)がないためで、その結果、ある世代が次の世代を生み出す効率に熱力学的な限界がある。その結果、Eの進化は制限される。同様に、人間以外では、ある種でXを増加させる適応が他の種に選択圧として作用するような共進化によってXの進化が制限される。
非体系エネルギーを用いることで、人間はXと Eをこれらの制限から解放した。しかし、エネルギー変換の効率よりも、システムに流入するエネルギーを取り込む能力にばらつきがあるのではと仮説している。エネルギー効率は熱力学的な限界によって制限されるが、エネルギーの取り出しは、文化や技術による急速な進化のため、共進化反応の影響から解放される。つまり、エネルギー効率への進化よりも、エネルギー取り出しへの進化の方が、システムにとって容易であったということである。例えば、世界の木材採取量の増加率は、木材燃焼のエネルギー効率の増加率を上回ると予想される。他の燃料についても同様の仮説を立てることができる。
環境収容力の進化
キャリングキャパシティモデル
人間の環境収容力(K)は固定された量ではなく、人間がその一部である地球システムの創発的特性である(Sayre 2008;Chapman and Byron2018)。つまり、Kは人間にとって外生的なものではなく、気候、生物多様性、生物地球化学サイクルと相互作用する人間の技術や行動によって決定されるものである。過去数世紀にわたり、人間は、環境からのエネルギーと物質の抽出量を増やし、それらのエネルギーと物質をより効率的に繁殖力と生存力に変換する技術を駆使して、Kを繰り返し増加させてきた。つまり、環境収容力は集団レベルの表現型であり、淘汰の重要なターゲットであり、ヒトにおいては遺伝子、文化、技術、環境の相互作用から生まれるものである。つまり、トップダウンの規制がない場合、ヒト集団は遺伝的・文化的進化によってボトムアップの規制を克服するよう選択されてきたという仮説である。
より具体的には、t+1世代がt世代の子孫だけで構成される(つまり、)t世代が t+1世代を生んで死ぬ)離散型人口増加系を想定する。この場合

ここで、Ntは時刻tにおける母集団の大きさ、Fは式1で定義される体力である。したがって、母集団の大きさは単純に全個体の体力の合計である。式1より代入すると

つまり、次世代における個体数は、個体群全員が取り出したエネルギーの総和に、各個体の変換効率を掛けたものである。母集団がKであると仮定すると、Ntは Nt-1に等しく、Kは Ntに等しい。したがって

個体レベルでは、Xか Eのどちらかに遺伝的変異があると仮定する。この遺伝的変異が選択されると、両者は正の関係にあるため、Kが増加する。
この場合、Xや Eが高い集団はKを増加させ、その結果、集団規模を拡大させることになる。他の集団に比べて人口規模が大きくなることで、社会的支配が可能になり、Xを増加させるための遺伝子や技術の普及率が高まる可能性があることを示唆している。
種が環境収容力を変化させる能力は重要である。Kが外生的に決定される場合、環境からの物質とエネルギーの抽出を増加させるような淘汰は働かない。しかし、Kが内生的に決定される場合、つまり、ある種が文化や技術を通じてそのKを決定できる場合、Kを増加させる文化的または遺伝的選択が存在する可能性があると考えられる。Kが増加すると、今度はその新しいK まで集団が拡大し、再び集団に選択的圧力がかかることになる。しかし、集団はより大きくなり、集団認知力も高まり、Kの増加にさらに適応するようになる。これは、母集団のサイズが大きくなるにつれて、母集団における突然変異の総頻度が増加することと類似している。集団の規模が大きくなるにつれて、任意の有益な突然変異が起こる確率は各世代で増加する。同様に、人間の人口規模が大きくなると、イノベーションに専念できる頭脳の数が増えるため、イノベーションの確率も高くなる。
図3
技術革新が急速に進み、人口が増加するとKの変化率は減少する
積載量シミュレーション
この議論を説明するために、人口増加、エネルギー抽出、環境収容力の進化を追跡する簡単なシミュレーション人口モデルを作った。このモデルは、環境からのエネルギー抽出が環境収容力に取って代わるという、標準的な離散的人口成長モデルに基づいている。したがって

ここで、Rは固有成長率で式1 のFに等しく、Xmaxは系のエネルギー収容力,Xtは全人口のエネルギー抽出量である。つまり、Xmaxと Xtは、標準的な離散的人口成長モデル(式)3、4のように)におけるKと Ntに代わるものである。Xmaxは、環境中のエネルギーと、そのエネルギーを取り込むための技術の両方によって決定される。したがって、Xmaxは技術革新によって変化する可能性がある。私たちは、Xmaxの変化は人口規模の一次関数であると提案するが、Xtが Xmaxに近づくとXmaxはより変化しやすくなるとも提案する。すなわち、集団が抽出するエネルギーの総量が抽出可能な最大エネルギーに近づくと、Xmaxを増加させようとする選択的圧力が生じるのである。そこで、時刻tにおける技術革新の確率Pを次のように定義する。

係数0.01は、1人あたり1世代に1回の確率でイノベーションが起きると仮定していることを示す。このモデルでは、0から100までの乱数を生成し、その乱数とPtを比較する。Ptが乱数を上回ればイノベーションが起こり、Xmaxは10%増加し、それ以外はXmaxは一定に保たれる。Xmaxの変化は正で永続的であると仮定する。式1と式3を式5に代入し、Xtについて解くと次のようになる。

ここで、FとEは前述の式1 で定義された通りである。ここでは、Eを1kcalあたり4×10-7の子孫に固定し、Xiを1世代あたり5×106kcalに固定すると仮定している。これらの仮定から、R(またはF)は2である。
図4は、10回のシミュレーションの結果である。当然ながら、エネルギー収容力Xmaxは時間とともに増加し、環境への負荷が増大していることがわかる。しかし、Xmaxの増加率も時間の経過とともに大きくなっている。図5は、図3に描かれたシミュレーションから得られたXmaxの平均値を、ベストフィットの指数方程式で示したものである。世代数とXmaxの関係が指数関数的であることから、エネルギー抽出量の増加率自体が時間の経過とともに増加することが示唆される。生物生態系が共進化して、このエネルギー抽出量の増加を抑制することができなければ、持続不可能な状況に陥ることになる。
図4
技術革新に伴うエネルギー収容力の進化に関するシミュレーション・モデルの結果
図 5
Change in energetic carrying capacity over time
このモデルには、寄生虫や病原体など、人間の個体数を制限するようなトップダウンの要因は含まれていないことに注意が必要である。寄生虫や病原体は、生物システムが人間システムの技術・文化的進化に「追いつく」ための一つの方法であるのかもしれない。
エネルギッシュなAETの応用例
エネルギーと社会生態学的崩壊
私たちが提示したモデルは社会生態学的崩壊そのものを説明するものではなく、むしろ社会生態学的崩壊につながる可能性のある非持続可能性や社会生態学的危機に対する人間の性向を説明することを目的としている。しかし、社会生態学的崩壊の概念に対するいくつかの批判は、私たちの議論を明確にするのに役立つかもしれない。Tainter(2006)は、社会生態学的崩壊における生態学的変化の役割は、歴史的に裏付けがないと論じている。その代わり、Tainterは古代社会の崩壊は主に社会的失敗によるものだと考えている。例えば、紀元前3千年後半にウル周辺のシュメール人が崩壊したのは、灌漑の不備と農業状況の悪化を認識できなかった指導者の失敗が原因であるとテインター氏は考えている。このように、テインターは、人口増加と環境エネルギーの採掘がシュメール崩壊の主要因であることに異論を唱えるかもしれない。しかし、環境収容力の進化という文脈で表現すると、土壌の塩類化によって環境収容力が低下し、それが土壌管理の不備の結果であるとテインターは示唆しているのである。私たちの見解では、持続不可能な灌漑の実施によってKが増加したため、Kの減少が可能になっただけだ。つまり、社会は技術を使ってXmax(つまりK)を増加させ、それに応じて人口が増加する。場合によっては、社会がこの新しいテクノロジーを使って資源を採りすぎてしまい、人口がKをオーバーシュートして、森林伐採や獲物の絶滅といった悪影響を及ぼすかもしれない。また、Xを増加させるために使用されたシステムが持続不可能であったため(例えば、シュメールにおける乾燥地農業)、人口がKをオーバーシュートせず、Kが人口規模よりも減少する場合もある。いずれの場合も、社会生態学的崩壊の「生態学的」が重要である。
私たちは、人間集団が環境収容力の進化に関連した2つの問題に直面していることを提案する。第一に、熱力学的な制約により、Kの継続的な増加が困難になり、最終的に人間集団が環境収容力を超え、集団が崩壊する事態に至る可能性がある。第二に、Kの増加は永久的ではなく、環境からのエネルギーや体内エネルギーの持続不可能な抽出に依存する可能性がある。この場合、人口はKをオーバーシュートしないかもしれないが、それでもKが低下するとKを超え、崩壊を経験する可能性がある。
Butzer(2012)は、社会生態学的崩壊は、10年~100年単位での経済・財政の衰退から始まり、10年単位での経済危機と連動して促進されると提唱している。Butzerは、これらの経済・財政状況の多くは、農業生産性の低下や人為的劣化などの生態学的なものであり、その他のものは社会経済的なものであると提案している(経済恐慌、外国からの攻撃)。しかし、上記のエネルギー的AETの観点からは、Butzerの言う前提条件やトリガーのほぼ全てがエネルギー的・生態的なものである。これは、経済活動とエネルギーの間の熱力学的な関連性によるものである(Georgescu-Roegen)1975;Daly1991)。つまり、すべての経済活動はエネルギーの散逸を意味し、Butzerが社会生態学的崩壊の引き金として「経済恐慌」を挙げるのは、人間社会が近年のように環境からエネルギーを引き出し、そのエネルギーを経済活動に転換することができなくなったことを意味する。同様に、Butzerが「外国からの攻撃」を崩壊の刺激とするとき、ここで提案するエネルギー的AETモデルは、侵略と紛争を、環境から大量のエネルギーを抽出し、それによってKを増加させるメカニズムとして捉えているのである。
エコモダニズムと環境保護主義者のパラドックス
エネルギッシュなAETは、エコモダニズム対技術的悲観主義の議論や、相互に関連する環境保護主義者のパラドックスに影響を与える。環境保護主義者のパラドックスとは、自然システムのウェルビーイングが低下しているにもかかわらず、人間のウェルビーイングは向上しているように見えるという観察である。Raudsepp-Hearneら(2010)は、環境保護主義者のパラドックスを定義し、その説明の可能性を4つ評価した。相互に排他的でない4つの説明の可能性は以下の通りである。(1)人間の幸福は不適切に測定されている、(2)人間の幸福は食糧関連の生態系サービスにのみ依存しており、そのサービスは増加している、(3)技術によって人間の幸福と生態系サービスが切り離された、(4)タイムラグによって現代の人口が将来の減少から隔離された。彼らの分析に対する完全な批判は本稿の範囲外であるが、Raudsepp-Hearneらは、人間の幸福は不適切に評価されてはいないが、他の三つの仮説を否定することはできないと主張した。
あるいは、エネルギー的AETは、人類が環境に対するエネルギー需要を増大させることによって、環境の擾乱による社会的な悪影響を回避してきたと提唱している。説明(3)に反して、テクノロジーは人間の生態系サービスへの依存を切り離したのではなく、どの生態系サービスに依存するかをシフトさせ、依存度を高めてきたのである。具体的には、人間は、自然システムからのエネルギーフローの減少を補うために、化石燃料、自然エネルギー、原子力の形でエネルギー補助金を利用してきた。例えば、人間は天然ガスに含まれるエネルギーを利用して、肥料生産のための窒素を減らしている。一人当たりの一次エネルギー消費量が、1971年の一人当たり石油換算1336kgから2014年の一人当たり石油換算1920kgに増加したように、エネルギー抽出量が人口増加よりも速く増加したため、人間のウェルビーイングは増加した。その結果、人間のウェルビーイングは向上した。
今後の方向性
競合する社会のエネルギー抽出
図1に示されるように、人間は一般的に非持続可能性の連続体の中で生活しているという指摘は、一部の先住民社会で観察されている持続可能性と矛盾しているように思えるかもしれない(Trosper)2002;Campbell and Butler2010)。しかし、これは本仮説の帰結である。エネルギッシュなAET仮説をゲーム理論の言葉で表現すると、人類社会の進化的安定戦略は、エネルギー抽出率の最大化と増加であるという仮説である。したがって、一部の先住民社会のように、環境からのエネルギー抽出量が増加しない持続可能な社会は、一時的で進化的に不安定な戦略であり、より抽出量の多い戦略と競合して取って代わられる、と考えられる。つまり、エネルギー抽出率が高く、持続可能性が低い社会が、エネルギー抽出率が低く、持続可能性が高い社会と競争してきたという仮説である。一般的な進化、特に文化的進化は価値中立的であるため(Sommers and Rosenberg2003)、持続可能な文化が進化的に不安定であることは問題かもしれないが、この仮説が否定されたわけでもない。
病原体の進化との融合
ここで論じたモデルは、人間の環境収容力がボトムアップの要因、具体的には環境からのエネルギー抽出によって制御されることを想定している(Hopfenberg)2003)。2020年のSARS新型コロナウイルス型の世界的流行が示すように、人間は病原体によるトップダウンの人口調整も受けやすく、人間の病原体は、人間のテクノカルチャーのように急速に進化できる生態系の数少ない部分の1つである可能性があるため、その影響を受けやすい。このように、人間の環境収容力は、環境からのエネルギー獲得と疾病の両方によって制限される可能性がある。
図2のようなフィードバックループに病害の進化を組み込むことが今後の研究者によって試みられるかもしれない。他の条件が同じであれば、人口密度の増加に伴い、病原体の毒性も増加すると考えられる(van Baalen and Sabelis1995;Lively2006;Mennerat et al.2010;Borovkov et al.)しかし、病原体の病原性の進化は複雑であり、病原体と宿主の両方の生態学的・遺伝学的要因や両者の相互作用の歴史に依存している(Knolle1989;Antia et al.)さらに、病気の細菌説、ワクチン接種、抗生物質の開発は、多くの人間社会が病原体にどう対応したかという技術文化的な変化を表しており、この変化は人口密度や環境からのエネルギー抽出と正の相関があると思われる。このように、病原体が人口増加やエネルギー採掘を制限する働きをするのか、あるいは技術革新や人口密度が病原体のトップダウン的な人口調節能力を奪う働きをするのか、今後の研究者にとって興味深いところであろう。
結論
社会がどのように非持続的な状態から持続可能な状態へと移行していくのかを理解することは、21世紀の環境社会科学において最も重要な問題の一つであろう。このような研究は現在も行われているが、そもそもなぜ人間社会は一般的に非持続可能な状態にあるのかを理解することにはあまり重点が置かれていない。
私たちは、人間社会が非持続可能な状態に陥りやすいのは、遺伝的・文化的な遺伝的変異に作用するマルチレベルの選択プロセスを通じて、環境からのエネルギー抽出率を最大化するように進化してきたからであると提案する。
具体的には、個人レベルでの遺伝的進化が行動の事前適応を生み出し(Van Vugt et al)2014)、文化的集団選択と協力して、単位時間当たりのエネルギー抽出量を増やす形質や技術が好まれるようになる。
このエネルギー抽出量の増加は、人口規模の増加を支えるために利用され、この人口規模の増加は、集団が新しいエネルギー抽出手段を革新する能力をさらに向上させ、人類の持続不可能性の究極の原因である正のフィードバックループにつながる(図)2参照)。
備考
- この命題に対する反論は、後述の6.1節で述べる。
- なお、私たちは「危機」という言葉をこのように具体的に定義して使っているにすぎず、価値観の相違を意図して使っているわけではない。