
Contents
pubmed.ncbi.nlm.nih.gov/30311830/
Published online: 12 Oct 2018
概要
光は生活に必要なものであり、人工光は視覚性能や安全性を向上させるが、光が健康や環境に与える潜在的な影響についての懸念が高まっている。多くの研究から得られた知見は、タイミングを誤った光曝露がヒトの概日リズムを乱し、さらなる健康への影響を引き起こす可能性があることを示唆している。しかし、光による概日リズムへの影響に関する個々の実験研究では、光曝露の定義や結果を含め、様々な方法が適用されている。したがって、これらの結果を総合的に検討するためには、系統的なレビューが必要である。
また、ライフサイクルアセスメント(LCA)における光害、すなわち人工光の悪影響の評価方法を開発するためには、光が概日リズムに与える影響に関する科学的根拠のレビューが必要である。現在のLCAの実務では、人の健康や生態系への影響の観点から光害を評価する方法がない。そこで、光と概日リズムという 2 つの概念を検索し、系統的な文献調査を行った。
サーカディアンリズムについては、メラトニンとレム睡眠の用語を追加して検索した。文献検索の結果、128 件の論文が得られ、データ収集・分析の対象となった。メラトニン分泌に関する研究は 122 件、レム睡眠に関する研究は 13 件であった。メラトニン分泌に関する報告は、通常は管理された実験室環境での特定の光曝露を用いた研究(101 報)と、家庭や職場環境での一般的な光条件を用いた研究(21 報)に分けられた。研究は一般的に20代または30代の成人を対象に行われてたが、子供や高齢者を対象にした研究はごくわずかであった。128件の研究のうち39件は10人以下の被験者で実施されている。
より深い合成を行うための研究の質の基準は、最低20人の被験者のサンプルサイズと、光照射の詳細(スペクトルまたは波長、照度、照度、照度または光子密度)を提供することであった。その結果、メラトニンに関する13件の研究とレム睡眠に関する2件の研究が認定された。これら15の報告をさらに分析した結果、夜間に2時間の青色光(460nm)に曝露するとメラトニンが抑制され、最も短い波長(424nm、バイオレット)で最大のメラトニン抑制効果が得られることが示された。メラトニン濃度は、露光の停止から15分以内にむしろ急速に回復し、メラトニン分泌に光の露光の短期または同時影響を示唆している。
メラトニンの分泌と抑制は加齢とともに減少したが、光誘起の概日位相の進行は加齢とともに損なわれなかった。夕方、夜間、朝の光照射はメラトニン量の概日位相に影響を与えた。また、最長波長(631nm、赤色)や間欠的な光曝露でもサーカディアンリセット反応が誘導され、夜間に目を閉じて寝ているときの低照度(5~10ルクス)への曝露でもサーカディアン反応が誘導された。このレビューにより、LCAにおける光害の評価方法をさらに発展させることが可能となった。
キーワード: 光、人間の健康、メラトニン、レム睡眠、健康、概日リズム
序論
人工光は現代社会の必需品である。自然光が利用できないときに照明を提供し、暗闇の後に多くの機能を可能にする。屋内外の環境における視覚性能や視認性を向上させ、人間の安全性や快適性を向上させることを目的としている。また、人工光は、リラックスや美容のための光源として、また、覚醒度や生産性を向上させるための光源としても利用されている。
人工光の意図は有益なものであるが、エビデンスの蓄積により、人間(Dominoni er al)。2016)や環境(Irwin 2018)への悪影響が指摘されている。人工光は、人間、動物、植物、生態系のバランスに悪影響を及ぼすことが疑われている(Tähkämö er al)。 照明製品の潜在的な環境影響は、いくつかのライフサイクルアセスメント(LCA)研究で評価されている(例えば、Scholand and Dillon 2012; Tähkämö and Halonen 2015; Franz and Wenzl 2017; Tähkämö er al 2014のレビューを参照)。LCA 法は、あらゆる種類の製品システムやプロセスの潜在的な環境影響(人の健康への影響を含む)を体系的に評価するためのツールである。しかし、これまでのランプや照明器具のLCAでは、光による影響は除外されていたが、地球温暖化、酸性化、オゾン層の破壊、資源の枯渇、生態毒性、人への毒性など、多くの環境影響を引き起こす他のエネルギーや物質の流れの影響に焦点が当てられていた。
光害は、人工光によるすべての悪影響の総和である(Commission International de l’Eclairage 2014)が、LCA で使用されている現在の特性化モデルでは、人工光が人間や他の種や環境の一部に与える影響を定量化して評価することができないため、事前の LCA 研究からは除外されている。LCA における光害を評価するための評価方法を開発すべきであることは認識されている(Cucurachi er al 2014; European Commission-Joint Research Centre – Institute for Environment and Sustainability 2011)。
LCA における光害の評価手法を開発するためには、まず、人工光が環境に悪影響を及ぼす科学的根拠を評価する必要がある(Cucurachi er al)。 予備的な文献検索(Tähkämö 2017)に基づいて、光が人間の健康に与える影響、特に概日リズムに関連する研究が多く見出された。動物相、植物相、生態系に対する人工光の影響は、科学的な文献では複雑で矛盾していることがわかった。動物への影響は種や個体数によっても異なり、植物や生態系の連鎖に関する研究は少ない。人間の健康への影響も同様に複雑である(Tähkämö 2017)が、特に概日リズムの乱れとそのマーカーとしてのメラトニン排泄の低下を介して、深刻な健康への懸念と光害との間の関連性についての予備的な示唆がある(Bedrosian and Nelson 2013; Cho er al 2015; Reiter er al 2007)。これらは、様々な実験設定、研究方法、および光曝露の定義を用いた、ヒトの概日リズムに対する光誘発性の影響に関する増加しつつある個々の研究に基づくものである。しかし、我々の知る限りでは、光が概日リズムに与える影響に関する系統的なレビューは存在しない。このテーマに関する先行研究に関しては、Cho et al 2015)は85の論文をレビューし、メラトニンの抑制、睡眠および覚醒に関連した、ヒトにおける健康への有害な影響を発見した。Souman et al 2018)は、ヒトの覚醒度に影響を与える光に関する68の論文をレビューし、多色光の強度を高めると主観的な覚醒度が上昇することを見出したが、レビューした論文の多くでは有意な効果は見られなかった。そこで、本文献レビューでは、光がヒトの概日リズムに与える影響に関する研究を収集し、総合的にまとめた。
ヒトの非視覚的概日リズム反応
ヒトでは、光の非視覚情報は目で検出される。非視覚刺激は、メラノプシンと下垂体アデニル酸シクラーゼ活性化ポリペプチドを含む本質的に感光性の網膜神経節細胞(ipRGC)によって検出され、視床下部の視床上核(SCN)に直接伝達される(Hannibal er al)。 SCNは、ホルモン分泌(メラトニン、コルチゾールなど)や体温など、毎日繰り返される生理機能を組織するマスターサーカディアンクロックとして機能する。マスター・サーカディアン・クロックは、目で知覚される明暗の変化によって同期しているが、特に夜間の人工照明による明暗のパターンの変化によっても乱れることがある。夜間の人工光への曝露(ALAN)は、過去数十年の間に毎年3〜6%増加していると推定され(Hölker er al 2010)さらに空間、時間、強度においても、輝度と範囲において毎年2%以上の増加を続けている(Kyba er al 2017)。
非視覚情報の検出には、視覚情報の検出に比べて高い光量が必要である(Brainard er al)。 1988; Zeitzer er al 2000)。光の強さに加えて、光のスペクトルや光の照射時間も光の非視覚情報に対する反応に影響を与える。また、年齢、遺伝子、年表型(すなわち、朝型の行動形質)などの個人の資質も、非視覚的な反応を誘発する光刺激に対する感度に影響を与えるようである。
概日リズムは、メラトニンのレベルを測定することによって決定することができる。メラトニンは、人間の体に夜を知らせるホルモンである。メラトニンの分泌は一日を通して周期的に行われており、夜間(暗闇)に分泌量が多く、日中に分泌量が少なくなる。夜間のメラトニン分泌量が多いレベルは、すべての人に平等ではなく、年齢や性別によって異なる(Follenius er al)。 メラトニンは、唾液、血液、尿のサンプルから測定することができる。メラトニンの一次代謝物である尿中メラトニン(6-スルファトキシメラトニン)は、24時間の間、2~8時間ごとに採取される場合は、一晩あたりのメラトニン産生量と一日あたりのメラトニン産生量を推定するために使用されるが、概日リズムの位相変化に注目する場合は、血液(血漿)または唾液サンプルで一般的に行われるように、10~30分ごとのサンプリングをより頻繁に行う必要がある(Benloucif et al 2008)。血漿中のメラトニン濃度は、通常、唾液から評価されたものよりも約3倍高い。正常なメラトニン分泌の乱れは、ヒトにおけるいくつかの疾患、特にいくつかのタイプの癌、メタボリックシンドローム、および精神障害(Touitou er al 2017)と関連しているが、人工光への暴露とこれらの条件との間の因果関係は証明されていない。国際がん研究機関(IARC)は、概日リズムの乱れを含むシフトワークを、おそらくヒトに対して発がん性があると分類しており、夜間の光曝露ががんリスクに寄与する可能性があることを示している(Straif er al)。
夜間や夜間の光曝露は、正常なメラトニンの分泌を抑制したり、遅らせたり(位相をずらす)することがある。光曝露がメラトニンの概日リズムの位相に及ぼす影響は、位相シフトの大きさと方向を示す位相応答曲線(PRC)によって記述される。PRCは、典型的には、生物学的に早い夜の遅延中の光曝露と生物学的に遅い夜と早朝の間の曝露が概日位相を前進させることを示している。PRCは、様々な暴露レベルおよび持続時間について決定されている(例えば、Khalsa et al 2003; Kripke et al 2007; Revell et al 2012; Rüger et al 2013; St Hilaire et al 2012)。
メラトニン分泌に加えて、光曝露は睡眠覚醒サイクルに影響を与える。したがって、急速眼球運動(REM)睡眠タイミングはマスターサーカディアンクロック(Lee er al 2009)によって制御されているため、サーカディアンリズムの乱れは、脳波(EEG)を用いたREM睡眠パラメータの測定によっても検出することができる。
方法
光による概日リズムの乱れ(メラトニン分泌の抑制と位相シフト、レム睡眠パラメータへの影響を含む)に関す る査読付き科学雑誌論文を対象に、系統的な文献検索を行った。Brocke er al)。 (2009)が示唆するように、文献レビューは、(1)レビュー範囲の定義、(2)トピックの概念化、(3)文献検索の実施、(4)見つかった文献の分析と合成、(5)研究課題の計画を立てなければならない。この文献レビューの意図は、研究のギャップを見つけることではなく、光がヒトの概日リズムに与える影響に関する既存の科学的証拠を分析することであるため、最初の 4 つのステップは本レビューに適用されるが、5 番目のステップは適用されない。
範囲
調査範囲は研究成果、すなわち既報の研究成果であった。文献検索は、概念的整理を用いて、一定の概念(代わりに歴史的整理や方法論的整理)に分けて行った。文献の対象者は一般の学者を想定しており、文献検索は文献の網羅的な網羅性を目指した。
概念
検索は、光と概日リズムの 2 つの主要な概念に焦点を当てて行われた。そのため、光に関連する検索用語は(「光曝露」or「人工光」)概日リズムに関連する検索用語は(「概日」or「メラトニン」or「レム睡眠」)とした。検索語は論文のタイトル、要旨
、キーワードで検索した。検索用語は、「光」が曖昧であること、慣用句やフレーズで頻繁に使用されていることから、「光」のみを含まないようにした。
文献検索
人間の健康と照明の分野における潜在的に関連性のある研究をすべて網羅するために、文献は4つのデータベースで検索された。PubMed、Scopus、ProQuest、Web of Scienceで検索した。検索は2017年11月1日に実施した。検索を実行し、検索結果を組み合わせ、重複を除去した結果、556本の論文が得られた(図1)。論文は、タイトル、要旨
、最終的に全文を基に関連性があると判断され、128件の関連論文が得られた。
図 1. 検索語と論文の選択による文献検索プロセス
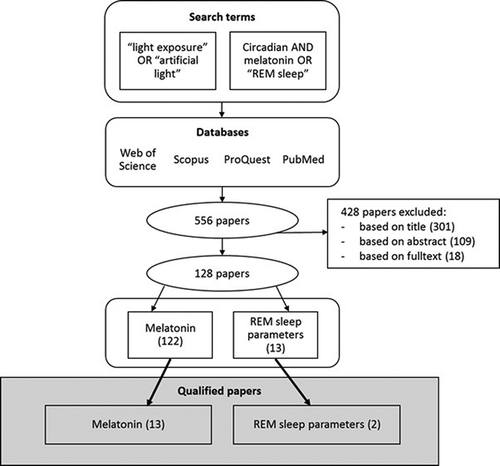
分析の対象とした論文の包含基準は以下の通りである。(1)学術論文(学会論文や書籍の章を除く)(2)独自の実験研究を発表した論文(レビューや他で発表されたデータに基づく研究を除く)(3)健康なヒトを対象とした研究(患者や動物を対象とした実験を除く)(4)測定された光曝露レベル(光レベルが主観的に評価された研究や全く測定されなかった研究を除く)(5)英語で書かれた論文、(6)メラトニンレベルまたはレム睡眠のいずれかまたは両方を測定した論文(概日リズムやヒトの健康に関連するその他のパラメータを除く)とした。人工光源と自然光源の両方からの光曝露(ディスプレイを含む)が含まれていた。
研究数が多く、質、研究方法、曝露の詳細に多様性があるため、より詳細な分析のために最高品質の論文を選択するために、追加の品質基準を決定した。品質基準は、(1)最低20名の被験者、(2)与えられた光曝露のスペクトル特性(スペクトルパワー分布(SPD)または波長)(3)照度、照度および/または光子密度で示された曝露のレベルである。その結果、15編の論文が採択された。
結果
データ収集
128 のフルテキスト論文から収集したデータはに記載されている。収集されたデータには、被験者の数、性別および年齢、光暴露の強度、持続時間、タイミング、スペクトル特性およびランプの種類、光レベルの測定点、研究の場所(実験室 vs. 典型的な生活環境)メラトニンサンプルの種類、および/または提供されたレム睡眠パラメータが含まれている。多くの研究は、体格指数、食事および視力などの被験者の特徴に関する情報を提供し、特定の生活様式を有する被験者に対して実験を制限した;例えば、アルコール消費量がないかまたはほとんどない、メリディアン横断旅行がない、夜勤労働がない、カフェイン摂取量がないかまたはほとんどない、喫煙がない、薬または薬物がない;および/または特定の生活様式を維持するように被験者に推奨し、典型的には実験に先立って規則的な睡眠/覚醒サイクルを維持するように指示していた。実験前の行動についての参加者の選択またはそれらの指示の詳細は、文献レビューでは収集されなかった。
- (SM1)は、メラトニン分泌に影響を与える特定の意図的な光曝露に関する101件の研究を提示している。
- (SM2)は、SM1の意図された暴露とは対照的に、一般的な光条件での21の研究から収集されたデータを列挙している。
- レム睡眠パラメータ(時間、%、潜伏時間)に関する13件の研究の詳細は、(SM 3)に記載されている。
これら3つの論文リストのうち、品質基準を満たした論文のみを「適格論文の分析」の項で詳細に分析している。
メラトニン分泌に影響を与える特定の光照射
意図的な光曝露がメラトニン分泌に及ぼす光誘発性の影響に関する研究(SM 1参照)は、1980年から 2017年の間に発表された。被験者数は4~72人で、典型的には1研究あたり5~20人であった。101件の研究のうち約29件が10件以下のサンプルサイズで実施された。被験者の年齢は典型的には20歳から30歳で、15歳未満の子供や50歳以上の高齢者を対象とした研究はごくわずかであった。光強度レベルは、照度(lx)照度(W/m2)および/または光子束(光子/m2/s)または非常にまれに輝度(cd/m2)で示された。露光の光レベルは0から数千ルクスまでの範囲であり、特定されるべき典型的な範囲はなかった。10ルクス以下の薄暗い光または暗闇(0ルクス)は、頻繁にコントロールまたは比較として使用されていた。光量が測定された場所は常に記載されなかったが、それがあったとき、それは最も頻繁に被験者の目(また、角膜、額)または目の近く(首、手首の周りのペンダント)で測定された。また、通常は視線の方向をコントロールして測定する場合には、視線の方向で測定していることが示されることもあった。被験者の目の状態は、頻繁に表示されたり、制御されたりするものではなかったが、例えば、被験者が意識を保っている場合、点での注視を求められている場合、読書を許可されている(開いている)場合など、プロトコルに基づいて推測することが可能であった。被験者の目が拡張されたのは8つの研究であった。瞳孔の拡張がメラトニン分泌の結果としての反応を増加させることは明らかである。
SM1では、曝露の時間的特徴は、パルス状の断続的曝露から数時間にわたる一定の曝露まで様々であった。曝露の持続時間は、典型的には1時間から5時間であった。研究では、同じ日の昼間、夜の間、または翌日の間に、様々な繰り返しの手順が含まれていた。曝露の詳細なタイムスケジュールは表に収集されなかった。暴露は、研究の明確な大部分(101研究のうち71)は、夕方および/または夜に光に被験者を公開して、一日のすべての時間でスケジュールされていた。
SM 1に示されたほとんどの研究(101件中76件)は、実験室または他のよく管理された施設(模擬生活施設、病院、研究施設)で完全に実施された。わずかな研究(101件中4件)は、作業環境で実施されたものであり、一部の研究では、被験者が、全体的に(自宅での測定)または部分的に(実験室での測定)典型的な生活環境(職場と自宅を含む)での生活を継続することを許可したものもあった。また、特定の実験場所が明示されていない場合もあるが、例えば、家庭環境では厳密に管理された光環境が得られないなど、プロトコルに基づいて想定することが可能である。
SM 1 に記載されている研究では、唾液、血液(血漿)または尿サンプルからメラトニンレベルを評価しており、唾液と血液サンプルが最も頻度が高かった。メラトニンサンプリング中の被験者の姿勢はメラトニンレベルに影響を与える可能性があるが(Deacon and Arendt 1994)すべての研究で記載されていないため、SMの表には収集されていない。
光曝露のスペクトル特性のレベルは、研究では非常に異なるレベルで示された。多色光と単色光の両方が使用され、研究の一部ではいくつかの異なる単色光で実験が行われた。光曝露のスペクトルパワー分布を提供した研究や、放射線の正確なピーク波長(半ピーク帯域幅を持つ)を示した研究は非常に少数であった。ヒトの概日リズムへの影響は光曝露のスペクトルに依存するため、スペクトル特性のこのような正確な測定は、結果を正確に評価するために必要である。ケルビン(K)で表される相関色温度(CCT)は、非常に異なるスペクトルによって得られる光の色の大まかな表現である。CCTは照明設計には有用な測定値であるが、健康関連の実験では使用されない。
研究は様々な光源を使用して行われた。ほとんどの場合、蛍光灯(FL)と発光ダイオード(LED)が使用されたが、一部の研究では昼光から放電灯、白熱灯まで、いくつかの異なる光源を使用した。また、一部の研究では特定の波長の放射線を遮断するためにフィルターが適用されていた。
メラトニン分泌に影響を与える一般的な光曝露
SM2には約21件の研究が掲載されているが、これは、あらかじめ定められた特定の光の照射量を実験するのではなく、通常の生活環境(家庭、職場、学校)に存在する光の条件を測定または制御したものであるためである。したがって、光源の種類やそのスペクトルの詳細は、SM 2に記載されている研究では頻繁に提供されていなかった。研究は1994年から 2017年の間に発表されたものである。サンプルサイズの範囲は5~528人と大きく、21研究のうち4研究は最大10人の被験者で実施されていた。被験者は通常20歳から40歳であり、SM1で提示された研究よりもやや高齢であった。1つの研究は、未就学児(Akacem er al)。2016)および1つは高齢者(Obayashi er al)。2014)の成人を対象として実施された。光レベルは一般的に1日24時間測定され、長期的な光曝露プロファイルについての洞察を提供した。
レム睡眠に影響を与える光曝露
光のレム睡眠パラメータへの影響を評価した研究は、SM 3に収集されている。これらの研究は、光曝露がレム睡眠時間、レム%、および/またはレム睡眠潜時に与える影響を研究したもので、1995年から 2017年の間に発表されたものである。被験者数は8~30人で、13研究中6研究はサンプルサイズ10以下であった。被験者の年齢は一般的に20~30歳の範囲であり、高齢者を対象とした研究は1件のみであった(Münch et al 2011)。光曝露の強度は数十ルクスから数千ルクスの範囲であり、ある研究では5ルクスと10ルクスの影響を比較し(Cho er al 2016)別の研究では電子書籍からの光(32ルクス)と紙の本からの反射光(1ルクス)の影響を比較した(Chang er al 2015)。光曝露は、パルス光を使用した1つの研究(Zeitzer et al 2014)を除き、典型的には1日の任意の時間帯に2〜8時間の連続的な光曝露であった。被験者は典型的に目を開けたまま覚醒していた。2件の研究(Cho er al 2016;Zeitzer er al 2014)のみが、睡眠中の被験者を夜間に光に曝露した。
資格のある論文の分析
表1は、サンプルサイズが 20 以上の基準に適合し、光曝露の強度(照度、照度または光子密度)とスペクトル特性(SPD または波長)に関する情報を提供している論文の詳細を示したものである。ここでは、光曝露の影響を受けたメラトニン分泌やレム睡眠の主な結果を、それぞれの論文を簡単に紹介する。
表1.
| 筆頭著者 | 年 | 被験者数(f、m) | 被験者の年齢:平均または中央値±SDまたはSEM(範囲) | (明るい)光の強さ | 薄暗い光の強さ | で測定された強度 | 曝露時間 | 露出のタイミング | 目の状態 | ロケーション | 測定されたメラトニン(唾液、血液、尿) | レム睡眠パラメータ | 波長、SPD、色、CCT | 光源タイプ(FL、LEDなど) | *) 追加情報 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ブレイナード | 2001年 | 72(37f、35m) | 24.5±0.3a(18–30a) | 0 µW / cm 2 ; 0.01 µW / cm 2 ; 1.5 µW / cm 2 ; 2.3 µW / cm 2 ; 3.1 µW / cm 2 ; 6.0 µW / cm 2 ; 12.1 µW / cm 2 ; 24.2 µW / cm 2 ; 42.2 µW / cm 2 | 目の高さ | 1.5時間 | 夜2h–3h30 | 開いて拡張 | ラボ | 血液 | 単色、420〜600 nm(420、440、460、480、505、530、555、575、600 nm) | キセノンアークランプ | 55人の白人、9人のアジア人、4人のアフリカ系アメリカ人、3人のヒスパニック、および1人の未知の民族 | ||
| タパン | 2001年 | 22(4f、18m) | 27±7a(18–45a) | 0.7〜65.0 µW / cm 2 | 目の高さ | 0.5時間 | 個別に設定されたタイミング:23.30〜02.30時間の夜(概日時間CT 16〜18) | 開いて拡張 | ラボ | 血液 | 単色:ラムダ最大424、456、472、496、520、および548 nm; 半帯域幅13nm | メタルハライドアークランプ | |||
| Cajochen | 2006年 | 11 + 10(11m(調査1); 5f、5m(調査2)) | 25.3±3.6a(研究1); 28.4±9.9a(研究2) | 12.1μW/ cmで2 460nmで、10.05μW/ cmのための2 550 nmのため、2.8 * 10 ^ 13光子/ cmで2 / sの460および550nmでのために | <15lx(周囲); 0 lx(ライトなし、コントロール) | 視線の角度 | 2時間 | 2130〜2330時間 | 開いた | ラボ | 唾液 | 単色、460nmまたは550nm(+ -10nm HPBW)でフィルタリング | アークキセノンランプフィルター | 研究1 =軽い研究; 研究2 =概日研究 | |
| Sletten | 2009年 | 21 | 若い23.0±2.9a以上65.8±5.0a | 6個の* 10 ^ 13個の光子/ cmで2 /秒(456 NM:26.4μW/ cmで2 ; 548 nmで:21.7μW/ cmの2) | 0 lx | 2時間断続的 | 薄暗いメラトニン発症(DLMO)の8.5時間後、朝06-08h | 開いて拡張 | ラボ | 血液 | 単色ブルーラムダ最大456nm; 緑548nm; 最大帯域幅の半分10nm | 水銀ランプ、メタルハライドアークランプ | 11人の若い+10人の年配の大人のうち、7人は両方の光条件にさらされ、3人は青い光だけにさらされました。もともとは11人の若者と15人の高齢者でしたが、サンプルが不足しているかメラトニンのリズムが検出できないため、5人の高齢者が除外されました。 | ||
| クローリー | 2010年 | 33(23f、10m) | (15–17a) | 156.2μW/ cmで2または540.8μW/ cmで2 | 目から18インチの距離で | 1時間 | 朝(週末の起床時) | 開いた | 自宅(睡眠)、実験室(DLMOの評価) | 唾液 | 454〜484 nm | 導いた | 16は通常のスケジュールにさらされ、17は特定の光条件にさらされました | ||
| グーリー | 2010年 | 48 | (18〜30 a) | 2.52 * 10 ^ 11から1.53 * 10 ^ 14光子/ cmで2 / sの、0.04から27 LX(460 nm)を0.6 -375 LX(555 nm)を(完全に16個の異なる照度レベル) | 目の高さ | 6.5時間 | 夜(約22:00–05:00 h) | 開いて、拡張 | ラボ | 唾液、血 | 青460nm、または緑555 nm; HPBW 10〜14 nm | キセノンアークランプ | 各露光で24(青と緑) | ||
| シャーキー | 2011 | 25(13f、12m) | 21.8±3a | 〜225 lx | <1 lx | 角膜 | 1時間 | 目覚めた後の朝 | 目覚めていると仮定 | ホーム(モニタリング週)、ラボ(概日位相評価) | 唾液 | 短波長(青)470 nm FWHM〜20 nm | LED | ||
| アップルマン | 2013年 | 21(11f、10m) | 22.5±3.9a | 40 lx、40 µW / cm 2 | 角膜; 手首 | 2時間(朝)、3時間(夕方) | 朝、夕方 | 起きているときは開いており、眠っているときは閉じていると想定 | ラボ | 唾液 | 青色LED(ラムダ最大476±1 nm、FWHM 20 nm)またはガラスフィルタリング<525 nm | LED、フィルタリングゴーグル | オレンジ色または青色光のゴーグルを着用して、朝2時間、夕方3時間の露光量を測定しました。角膜で光レベルが40lxに設定されたゴーグルに取り付けられた青色LED | ||
| シェラッパ | 2013年 | 30(16m、14f) | 25.2±3.1a(20–31a) | 通常、25〜32 lx(27、29.8〜27.6 lx)。2.27、6.6、および2.52 * 10 ^ 16フォトン/ m2 / s(380〜780 nm) | <8 lx; 闇; | 目の高さ | 2時間(1.5時間の薄暗い光、2時間の暗闇、2時間の露光) | 夕方:21:30–23:30 h(薄暗い18–19:30 h、暗闇19:30–21:30 h、明かり21:30–23:30 h) | 開いた | ラボ(想定) | レム% | 与えられたSPD、6500、2500、および3000 Kの多色光(注:実際には5984、2481、および2581 K) | CFL、INC | CFL 6500 K、CFL 2500 K、INC 3000 K、ただし実際にはCFL 5984 K、CFL 2481 K、およびINC 2581 K | |
| フィゲイロ | 2014年 | 23(17f、6m) | 31.1±11.1a(18–51a) | 明暗の露出は、朝と夕方に、早いグループと遅いグループの対数ルクスで与えられたデイズメーターで測定されました。 | 朝2時間、夕方3時間 | 朝と夕方、時間はベースラインDLMOに依存 | 開いていると仮定 | ホーム(露出)、ラボ(テスト用) | 唾液 | 青色LEDとオレンジ色のメガネに与えられたSPD。LEDλmax= 476±1nm、半値全幅〜20 nm、オレンジ色のメガネ:525nm未満のほぼすべての光放射をフィルタリング | 導いた | 11後期、12初期クロノタイプ | |||
| ホーミエン | 2014年 | 24メートル | (21–28a) | 10 ^ 13光子/ cmで2 / S(赤); 2500 lx(白) | 目の高さ | 6時間の連続赤色光、断続的(約1分のオン/オフパターン)赤色光または明るい白色光 | 夜、約23〜05時間 | 開いた瞳孔径を測定 | ラボ | 唾液 | 赤631nm狭帯域幅、クールホワイト4000 K | 赤はLED、白はFL | |||
| サンダー | 2015年 | 29(13f、16m) | 69.7a | 280 lx、920 mW / m 2 ; 240 lx、710 mW / m 2 ; 140 lx、392 mW / m 2 | 手首(連続光測定)、角膜(朝の露光) | 5時間 | 朝/日:08–13時間; 午後13〜18時間 | 開いた | ホーム | 唾液 | 与えられたSPD:午後の光のために青が豊富で、青が抑制され、別の青が抑制された | 導いた | |||
| チョ | 2016年 | 23メートル | 22.26±2.65a(21–29a) | グループA:5 lx、グループB:10 lx | 目の高さ | 8時間 | 夜(一晩)23–07時間 | 寝ている間閉まっている | ラボ | レム%、レム睡眠潜時(分) | ピークラムダ463.6nm、5779 K | 導いた | 当初は30人の被験者が採用されましたが、品質管理プロセス中に7人が除外されました | ||
| Geerdink | 2016年 | 42(23f、19m) | 21.4±6.5a | 300 lx、3.0 W / m2(青); 250 lx、0.5 W / m 2(琥珀色) | 角膜 | 0.5時間 | 目覚めた直後の朝 | 目覚めていると仮定 | ホーム | 唾液 | 青(ピーク470 nm)、または琥珀色(ピーク590 nm) | 導いた | |||
| Najjar | 2016年 | 39(25m、14f) | 26.4±5.06a | 1700〜1805lx連続または夜間パルス | 0.6〜1.9 lx(日中および一定の姿勢) | 異なる間隔(0.004および0.4 Hz)で1時間連続または2ms点滅 | およそ02–03時間の夜 | 開いた | ラボ | 唾液 | 同様の広域スペクトル白色光(夜間露光); 補足情報で提供されるSPD | 日中の薄暗いためのFL; 広域スペクトル400Wブルーメタルハライド(UVフィルター付き)。点滅光用キセノンランプ | 31はさまざまな間隔で断続的な光にさらされ、8は連続的な光にさらされます |
メラトニン分泌やレム睡眠に影響を与える光曝露に関する研究で、少なくとも 20 人以上の被験者が参加していること、光曝露の強度とスペクトル情報(照度、照度または光子密度、SPD または波長)が与えられていること、などの品質基準で評価されたもの。光強度とは、照度(lx)照度(W/m2)光子密度(photons/cm2/s)輝度(cd/m2)を指す。略語
CCT=相関色温度(ケルビン、Kで表す)FL=蛍光灯、INC=白熱灯、LED=発光ダイオード、SPD=スペクトルパワー分布。
Brainard et al 2001)は、420から600 nmの波長で夜(02-03:30 AM)にフルフィールド、単色光に72人の被験者を露光した。被験者の瞳孔を拡張し、血液サンプルを暴露の前後に収集した。各被験者は、暴露の間に少なくとも1週間で1つの波長の少なくとも7つの異なる放射照度にさらされた。フィットしたアクションスペクトルは、446〜477 nmの波長範囲で最も効果的なメラトニン抑制を示す627個々の実験の合計から収集したメラトニン抑制データから構築された。このように、本研究は、ヒトの概日性光受容を媒介する単一の光色素であり、視覚応答を司る桿体細胞や円錐体細胞の光色素とは異なる波長領域の放射線に対してピーク感度を持つことを発見することに貢献した。少なくとも3.1 µW/cm2の放射照度でメラトニンの有意な抑制が見られた。
(Thapan et al 2001)は、光誘発性夜行性メラトニン抑制のスペクトル感度のための作用スペクトルを作成することを目的として、合計215例の単色光に曝露された22人の被験者を対象にメラトニン抑制を測定した。被験者は、ピーク波長(424, 456, 472, 496, 520, 548 nm)を変化させ,0.7から65.0 μW/cm2の範囲の放射照度で、夜間(23:30から02:30 hの間)に30分間の光パルスを受けた。被験者は、光曝露なしのベースラインの夜と瞳孔を拡張した状態で光曝露を行った2つの夜を含む、3つの連続した夜(19-07時間)の1から16のセッションを完了した。各波長を5〜8回の照射レベルで試験し、各照射レベルを3〜7人の被験者で試験した。メラトニンレベルは、光照射前(90分前と15分前)光照射中と照射後(光照射開始から1.5時間、照射2時間後から15分ごと)および光照射前2日間の尿から採取した血液から評価した。血漿中のメラトニンは、すべての波長で照射量の増加とともに抑制され、メラトニン抑制と光曝露との間に強度依存的な関係があることが明らかになった。統計的に有意なメラトニン抑制効果が得られたのは、424, 456, 472, 496, 520, 548 nmで、それぞれ1.9, 2.0, 1.8, 3.0, 7.0, 7.2 μW/cm2の最小照射照度であった。照射応答曲線をフィッティングすることにより、作用スペクトルを生成した。作用スペクトルのピークは、視覚感度関数(フォトピクティックまたはスコトピック)のピークに比べて短波長領域に多く位置しており、短波長領域での新たな光色素の存在を示唆していた。メラトニンは点灯後30〜45分後に最大に抑制されたが、30分間の露光が続いたことに注意しなければならない。30 分間の露光では、夜行性メラトニンの長期的な抑制効果は認められなかったが、メラトニン濃度は非常に急速に回復した(露光開始から 1 時間以内)。興味深いことに、よく引用されるフィットした作用スペクトル曲線とは対照的に、最大のメラトニン抑制効果は、試験された最も短い波長(424 nm)で発見され、続いて、順に、456 nm、472 nm、500 nm、520 nm、および548 nmであった。作用スペクトルの湾曲した形状は、459 nmでピークを持つロドプシンテンプレートに照射応答をフィッティングし、レンズの密度を補正した結果であった。
www.my-craft.jp/html/aboutled/led_hachou.html
(Cajochen et al 2006)の研究では、24人の被験者を12人の2つのグループに分けて研究した。光研究では、12人の男性被験者を、薄暗い環境光(<15lx)の中で30時間コントロールし、習慣的な就寝時に8時間睡眠させた。概日研究では、女性および男性の被験者12名を、夕方から16時間、ベッドで一定のリカンベント姿勢で管理され、最初に<2 lxで1.5時間、次に0 lxで2時間、次いで光曝露(青、緑または無光)で2時間、最後に<2 lxの多色光で1.5時間曝露し、これらの薄暗い、暗い、明るい、および再び薄暗い光の条件の後、被験者を睡眠に供した。メラトニンを唾液から測定した。夕方の青色光(460nm)への約2時間の曝露は、それ以前の低メラトニンレベルと比較して、メラトニンを有意に抑制した(12.0 pg/mLから8.8 pg/mLへ)が、緑色光(550nm)では抑制しなかった。
(Sletten et al 2009)の研究の目的の1つは、メラトニン抑制が加齢とともに減少することがわかっているため、概日位相の前進における青色光の有効性と、その反応が加齢とともにどのように異なるかを評価することであった。実験の1週間前と光照射前後の血漿から薄光メラトニンの発現(DLMO)を評価するために血液を採取した。約11名の若年成人と約10名の高齢者を対象に、実験の1週間前から実験期間中まで一貫した睡眠覚醒スケジュールを維持した。3日間の実験プロトコルは、ベースラインの夜、光照射の夜、および刺激後の夜から構成されていた。すべての若い被験者と10人の高齢者被験者のうち7人は、2つの実験セッションに参加した。高齢者被験者(3)の残りの部分は、青色光のみを露光した。2時間の露光は、早朝に発生している彼らのDLMOの8.5時間後に開始するように個々に時間を設定した。被験者の瞳孔は、露光中に拡張した。相の進行の大きさは、光曝露前後の血漿メラトニンリズム位相マーカーの差から評価した。サーカディアンの位相は、両年齢群ともに青色光と緑色光の両方で進んでおり、緑色光よりも青色光の方がわずかに進んでいたが、有意ではなかった。若年者の位相の進行は、高齢者(DLMO 2136 ± 0074 h、n = 15)と比較して、統計的な方法ではなかったが、より大きかった(DLMO 2104 ± 0043 h、n = 11)。彼らは、光誘発性概日位相進度が加齢とともに有意に損なわれていないことを発見した。
Crowley and Carskadon (2010)は2つの実験を行ったが、そのうちの実験2はこの更なる分析に含まれている。実験2では、週末の朝の短波長光がサーカディアンリズムに与える影響を、33人の若年者(15-17a)を対象に調査した。実験は2週間実施し、典型的な光曝露(「TYPICAL」)と特別に設計された光曝露(「LIGHT」)の2つのグループに分けて、被験者間デザインとして実施した。両群とも自宅での睡眠時間は平日に比べて1.5時間遅く、週末の起床時間は平日に比べて3時間遅く、週末の睡眠時間は平日に比べて1.5時間長くなった。被験者は、唾液からDLMO相を評価するために、各週末の前(金曜日)と後(日曜日)に研究室を訪れた。TYPICALグループの14人の被験者とLIGHTグループの14人の被験者は、分析のために両夜に使用可能な唾液のDLMO相データを持っていた。LIGHT群は、30%の強度レベル(4人の被験者)または100%レベル(10人の被験者)のいずれかで光曝露を受けた。DLMO位相は、46±34分で実験2のTYPICAL週末と38±28分でLIGHT週末に遅延したが、有意ではなかった。DLMO位相は光強度30%群と100%群で差がなく、これら2群のデータを合わせた。一般的に観察されている週末の睡眠スケジュールを維持した後、若年者の概日位相が遅れていると結論づけられた。週末の短波長光への朝の曝露は、本研究の対象者において概日位相を安定化させなかった。
(Gooley et al 2010)は、コーン光受容体が光反応の概日性調節に寄与しているかどうかを試験した。約48名の被験者が、時間的な手がかりのない実験室環境で9日間のプロトコルに参加した。最初の3日間は通常の睡眠・覚醒スケジュール(8時間睡眠、16時間覚醒)を190 lx未満で維持した後、3 lx未満に調光した。これは、強制的な覚醒とそれに続く8時間睡眠機会の50時間の一定のルーチンに続いた。次いで、拡張瞳孔を有する被験者は、ランダムな順序で16の異なる照度レベルでの2つの波長(青460または緑555nm)のいずれかの狭帯域幅光曝露の6.5時間に曝露された。被験者は90分間連続して光を凝視し、その後10分間、そうでなければ暗い部屋の中で自由な凝視が許可された。この夜間の光曝露は、メラトニン抑制と概日位相リセットのための用量反応曲線を決定するためであった。メラトニンレベルは、一定のルーチンと光曝露中の唾液サンプル、および睡眠中と覚醒中の両方の血液から評価した。夜間光曝露の開始時には、緑色光と青色光はメラトニンの抑制に等しく効果的であったが、その後の光曝露時には、緑色光に対する分光感度は青色光と比較して指数関数的に低下していた。このことは、コーン光受容体(λmax = 555 nm)が、光曝露の開始時および低照度では非視覚応答に実質的に寄与するが、長期または高照度の光曝露中には寄与しないことを示唆している。メラノプシンが高強度および長期のサーカディアン応答に関与していることが示唆された。その結果、狭帯域幅の緑色(555 nm)光(24ルクス以下)への短時間(90分未満)の曝露は、青色(460 nm)光(2ルクス以下)の同程度の光束と同等の効果がある可能性があることが示された。
(Sharkey et al 2011)は、25人の被験者を無作為に、起床後の朝1時間、青色光(ピーク470nm、225ルクス)または薄暗い光(<1ルクス)を受けるように割り付けた。ベースライン1週間後、被験者は6日間、個別に固定された7.5時間の高度な睡眠スケジュールを維持した。光曝露は頭に装着した光計で測定した。DLMOを評価するために唾液を採取した。6日後、両群間で概日位相に有意差は認められなかった。両群ともに概日位相が進んでいた:平均DLMOは、薄暗い光群で1.5±1.1時間、青色光群で1.4±0.7時間進んでいた。青色光群では、覚醒後 90 分間に短波長光を多く受けても、薄暗い光群と比較して位相シフトは促進されなかった。また、概日光曝露量(短波長加重照度)や覚醒エピソード全体の光学的照度には、薄暗い光群と青色光群の間に差は見られなかった。著者らは、青色光が薄暗い光よりも大きな位相前進を誘発することが見出されなかったため、青色光暴露のタイミングが位相前進を誘発するのに最適ではなかった可能性を示唆している。
(Appleman et al 2013)は12日間の研究を行い、21人の被験者を無作為に2つのグループに分け、朝2時間短波長(青色、ピーク476nm)の光を照射し、夕方3時間オレンジ色のメガネでフィルターをかけた光(<535nm)を照射した(前進群)か、逆のタイミングで、朝はオレンジ色のメガネ、夕方は青色の光を照射した(遅延群)かのどちらかを選択した。被験者は最初の5日間は通常のスケジュールを維持し、その後の7日間は固定された睡眠スケジュール(ベースラインから90分前倒し)に加えて、朝と夕方の光照射を受けた。この研究から、概日位相の変化の方向は固定睡眠スケジュールではなく明暗暴露によって決定され、概日位相をシフトさせるためには朝夕の光暴露の両方をコントロールする必要があることが示された。90 分間の固定睡眠スケジュールで 7 日後には、サーカディアン位相は事前群で 132±19 分、遅延群で 59±7.5 分進んでいた。測定した平均光曝露値(概日刺激(CS)概日照度(CLA)対数ルクスで表される光学的照度)には、両群間で統計的な差は生じなかった。
(Chellappa et al 2013)は、夜間の青色に富む多色光が睡眠に与える影響を研究した。彼らは30人の被験者の脳波を測定し、レム%データを提供した。被験者は、18時から 19時30分までは薄暗い光(8 lx未満)19時30分から21時30分までは暗闇(0 lx)21時30分から23時30分までは明るい光(約28 lx)の中に置かれた。これは、定格と測定されたCCT値の間に顕著な違いを示している。スペクトル特性をより正確に表現するために、彼らは先行研究(Chellappa et al 2011)でランプのSPDを提供したが、これは2500Kのコンパクト蛍光灯のとがったSPDと3000Kの白熱灯の滑らかなSPDの間の違いを示している。最初の3回のNREM-REM睡眠サイクルにおける睡眠構造には、光条件に関して有意な差は見られなかった。レム%は6500K光と3000K光の両方で18.2±0.9%、2500K光では17.9±1.0%であったが、光条件間では有意差は認められなかった。
(Figueiro et al 2014)は、固定された高度な睡眠-覚醒スケジュール上で、朝-クロノタイプと夕方-クロノタイプが光曝露に対して異なる反応を示すかどうかを研究した。彼らは、13日間(ベースライン評価6日間、光曝露と設計睡眠スケジュール7日間)の高度な睡眠・覚醒プロトコルを2回、1回は光曝露を前進させた場合、1回は遅延させた場合の研究を行った。前進光照射では、朝は短波長光、夕方はオレンジ色のメガネで短波長光を制限し、遅行光照射では、朝はオレンジ色のメガネで短波長光を制限した。2種類のクロノタイプ間では、光曝露に対する反応に有意な差は認められなかった。DLMOは、両群ともベースライン週と比較して、遅延光曝露後に有意に遅延し、促進光曝露後に有意に進行した。光誘発性の概日位相変化は、両クロノタイプについて以前に発表されたPRCによって予測されたものと一致していた。ベースライン週の被験者の光曝露は、光曝露週の被験者よりも有意に少なかったことが指摘された。
被験者間研究では、(Ho Mien et al 2014)は、24人の被験者を、露出の種類によって8人の3つのグループに分け、連続的な赤色光(631 nm、13 log photons/cm2/s)間欠的な赤色光(1分間のオン/オフ)および白色光(2500 lx)の影響を、時間の手がかりから解放された実験施設で実験した。実験は、26時間の一定のルーチン期間(覚醒)と8時間の睡眠期間から始まった。光照射は6時間持続した。光照射の前後に唾液性メラトニンを評価した。その結果、連続的な赤色光と断続的な赤色光は、ほぼ1時間の平均的な位相遅延で、同様のサーカディアンリセット反応を誘発することがわかった。赤色光はメラトニンの分泌を抑制することはなかったが、瞳孔収縮の延長を誘発した。しかし、個人差が認められた。赤色光を照射された2人の被験者の概日反応は、明るい白色光を照射された被験者のものと同程度の大きさであり、コーン光受容体が個人によっては概日位相のリセットに寄与している可能性が示唆された。これは、視覚光受容体が光誘起メラトニン抑制に部分的に関与している可能性を示唆しており、興味深い知見である。しかし、サンプルサイズが光曝露の種類ごとに8個と小さかったことは認められる。
(Sander et al 2015)は、29人の高齢者(>65 a)を対象に、無作為化クロスオーバーデザインで、自宅で午前中と昼間(08:00~13:00 h)に青色富化光(280ルクス)または青色抑制光(240ルクス)のいずれかの光を3週間にわたって2回照射する光実験を行った。午後1時以降は、すべての症例で光量を約140ルクスに低下させた。曝露期間は、晩秋(10月~11月)のコペンハーゲン地域で、曝露の間に1週間を置いて実施した。唾液は夕方の各光暴露期間の後に採取した(45分ごとに7サンプル)。唾液中のメラトニンプロファイルは、朝/昼間の青色光照射と青色光照射の間で有意な変化を示さなかった。被験者の測定された青色光暴露の唯一の有意差は、8:00〜9:00の実験からの青色富化光が、昼光からの寄与によるものであった。被験者のメラトニン濃度には大きな個体差が見られた:濃度は、検出可能な夜間の上昇がなく、すべての時間帯で<2 pg/mLから、深夜と早晩の間に約50 pg/mLへの明確な増加に幅を持っていた。
(Geerdink et al 2016)は、自宅でのプラセボ対照試験で、ブルーライトとアンバーライト治療のサーカディアン・フェーズシフトの影響を調査した。仕事の日に「社会的時差ぼけ」に悩む約42人の被験者を無作為に割り付け、両方とも30分間のパルスで与えられた同様の光学的照度を持つ高強度ブルーライトまたはアンバーライト照射(プラセボ)のいずれかを受けさせた。プロトコルは30日間持続した。睡眠制限のない14日のベースライン、睡眠促進スキームで朝に30分間の青色光またはアンバー光パルスのいずれかで連続した9日間の治療日、および睡眠制限のない7つの治療後の日。唾液性メラトニンは、ベースライン期間の開始時、中間時、終了時、光照射期間の後、治療後期間の後に採取した。光曝露は連続的に記録した。朝の青色光照射は、朝の琥珀色光(48±47分)に比べて、光治療期間中のメラトニンリズムの位相前進が有意に大きかった(84±51分(SD))。治療後の日中の起床時間は、被験者のベースラインと比較して、ブルーライト群ではわずかに早く(-21±33分)アンバーライト群ではわずかに遅く(+12±33分)なっていた。このことから、家庭での朝のブルーライトは、サーカディアン位相を進めるために利用できることが示唆された。
NajjarとZeitzer(2016)は、39人の被験者を対象に16日間の実験研究を実施し、連続光(n = 8)または2.5~240秒の刺激間隔を持つ2-msの光フラッシュのシーケンス(n = 31)のいずれかに60分間曝露し、曝露期間の前後の唾液サンプルでメラトニンの位相シフトと抑制を評価した。被験者は16日間のプロトコルの最初の14日間は自宅で規則的な睡眠/覚醒リズムを維持し、15日目と16日目の間は夜の検査のために研究室に滞在した。間欠的な光は、連続的な光よりも概日リズムの変化を誘発するのに効果的であることがわかった。1時間の連続光はメラトニン分泌を51%±40%抑制し、-0.60±0.34時間シフトさせた(遅延)。ヒトのサーカディアンシステムは、画像形成(視覚)光受容とは対照的に、ミリ秒単位の時間スケールで光を検出する能力を持っていることが実証された。フラッシュの間隔の長さは、位相シフト(遅延)に影響を与えるが、メラトニン濃度には影響を与えないことがわかった。刺激間隔を最大にしてフラッシュ光を照射した場合、同じ強度で3800倍の持続時間で照射した場合と比較して、概日系の位相遅延が少なくとも2倍以上になることがわかった。点滅光は、刺激間隔に応じて概日リズムを-2.99~0.30時間変化させた。
(Cho et al 2016)は、23人の被験者を対象に、夜間の薄暗い光の影響を調査した。被験者を無作為に2つのグループに分け、1つは5 lx(n = 11)もう1つは10 lx(n = 12)を23時間から07時間の夜間に投与し、被験者は薄暗い光条件の間、仰臥位で寝ていた。被験者は研究期間の1週間前から規則的な睡眠覚醒サイクルを維持し、薬、コーヒー、アルコールの摂取を控えるように求められた。プロトコルは3日間、最初の2日間は光を照射しない状態(第0夜と第1夜)で、第3夜(第2夜)はいずれかの薄暗い光を照射した状態で行われた。レム睡眠%と潜伏時間を測定した。睡眠構造には両群間の差は見られなかったが、睡眠構造には第1夜と第2夜の間、つまり夜行性薄暗い光への曝露なしと曝露ありの間に有意な差が見られ、その中には両夜の間のレム睡眠%の有意な差も含まれていた。レム睡眠潜時は、夜間で有意な夜群間相互作用を示したが、夜間での差は認められなかった。夜間薄暗い光への暴露は、レム睡眠量の増加と有意に関連していた。この知見は興味深いもので、夜間の光のレベルが比較的薄暗く(5または10lxのいずれか)被験者は光曝露中に目を閉じて寝ていたことを示している。このことから、夜間の光への曝露を避けることで睡眠の質が向上する可能性があることが示唆された。
資格論文のまとめ
単色光照射によるメラトニン抑制の評価については、2つの先駆的な研究がある(Brainard et al 2001;Thapan et al 2001)。波長に応じた照度のしきい値を持つ十分な強度の光照射(Thapan er al 2001)は、メラトニン分泌を抑制し、循環メラトニンレベルを低下させた。また、メラトニンレベルは、照射停止後約15分間は最大に抑制されたままで、その後は約15分で回復した(Thapan et al 2001)。
これらの2つの前述の作品は、青色光と緑色光への暴露の間の実用的な比較を刺激した(Cajochen et al 2006; Gooley et al 2010; Sletten et al 2009)。青色光への暴露は緑色光よりもメラトニンのレベルを抑制した(Cajochen er al 2006)、この効果は時間の関数としての大きさで現れた(Gooley er al 2010)のに対し、青色光と緑色光の暴露はメラトニンの概日リズムで同程度の位相シフトを生じさせ、この効果は年齢に依存しない(Sletten er al 2009)。
夜間には、断続的な暴露のいずれかの赤色光(ホーミエン et al 2014)または青色光(Najjarとツァイトツァー2016)にメラトニンの分泌を阻害しなかったが、これらはメラトニンの概日リズムの位相をシフトした。驚くべきことに、朝の起床後、1時間の青色光(CrowleyとCarskadon 2010; Sharkey et al 2011)または青色濃縮光(Sander et al 2015)への暴露は、メラトニンの概日リズムの位相をシフトする効果はなかった。しかしながら、青色光への曝露は、睡眠促進スキーム(Geerdink et al 2016)または相反する光曝露のコントロール(Appleman et al 2013; Figueiro et al 2014)と一緒に投与された場合、予想通りメラトニンの概日リズムの位相シフトをもたらした。
就寝前2時間の光への曝露は、その後のレム睡眠を変化させなかった(Chellappa et al 2013)。一方、8時間の光照射ではレム睡眠の割合が増加した(Cho er al 2016)が、睡眠の質が良いと判断される範囲(総睡眠時間の20~30%)にとどまった。
考察
現代社会や発展途上地域では、人間は、SPD、タイミング、持続時間、強度の点で自然光暴露パターンとは異なる人工光にさらされることが多くなってきている。自然光から人工光への光曝露が健康に与える潜在的な影響については、いくつかの科学的研究が明らかにしている。光が人間の概日リズムに与える影響に関する個々の研究とその知見を総合するために、系統的な文献レビューを実施した。このレビューは、人工的な光曝露による概日リズムの乱れについて理解を深めるとともに、照明製品の環境影響をLCA法で評価するための光害の人間の健康関連指標を開発するための基礎となる。
適格論文の批判的比較
文献検索の結果、光曝露の影響を受けたヒトの概日リズムに関する実験的研究をメラトニン分泌量またはレム睡眠パラメータのいずれかの観点から測定した128の論文が得られた。品質基準を満たした 15 本の論文を詳細に分析し、光が人間の概日リズムに与える影響についての科学的理解を深めた。その結果、夜間に光(460nm)に2時間曝露するとメラトニンが抑制され、光曝露のメラトニン抑制効果が最も短い波長(424nm)で最大になるが、メラトニン濃度は曝露停止後15分以内に急速に回復することがわかった。これは、光曝露の短期的または同時的な影響を示唆している。また、メラトニン分泌とメラトニン抑制は加齢とともに減少することがわかったが、光によるメラトニンのサーカディアンリズムの位相進行は加齢とともに損なわれていない。サーカディアンリズムは、朝と夕方の両方の光の照射(青く濃縮されているか、青く枯渇しているか)に反応し、朝型と夕方型の人(クロノタイプ)では有意な差がないことがわかった。
非限定論文に関する観察
SM 1の非限定論文に関しては、注目に値するパラメータは閉眼を用いた実験に関連している。光フラッシュを用いた最初の研究では、(Zeitzer et al 2014)は、参加者の小さなサンプルを暗闇または睡眠中に2ミリ秒の光フラッシュのシーケンスのいずれかに曝露し、就寝後2時間目から3時間目までの30秒ごとに与え、まぶたを閉じた。これは、睡眠自体に大きな変化を与えることなく、概日時計のタイミングをシフトさせた。同様に、FigueiroとRea(2012)は、1時間の間にまぶたを介して配信された光がメラトニンと位相シフトDLMOを抑制する方法を示した。どちらの研究も、光治療は目を閉じた状態でも、また、睡眠中であっても行われる可能性があることを示唆している(Zeitzer et al 2014))。しかし、光害の観点からは、睡眠導入後の光への曝露は、夕方の曝露よりも等しく有害である可能性があると結論づけることができる。
夕方の光曝露に対する個人の感受性については、(Canton et al 2009)では青色と褐色の虹彩色の影響の大きさに差は見られなかったが、Higuchi et al 2008)では瞳孔面積の個人間差がメラトニン抑制と正の相関を示すことが示されている。また、思春期初期や思春期前の段階では、思春期後期や思春期後の段階に比べて、低照度の150ルクスであっても、思春期後期や思春期後の段階では、思春期後期や思春期後の段階に比べて、思春期後期や思春期後の段階では、夕刻の光に有意に敏感であった(Crowley et al 2015))。
実験的な設定とは対照的に、SM 2は、概日リズムのために自然に発生し、個別に選択された条件に注意を引く。高緯度の暗い冬の季節は、他の季節と比較して、夜間の間に最も高い最大メラトニンレベルに関連付けられている(Adamsson et al 2016)。研究はまた、習慣的な就寝時間と光への曝露が概日リズムの位相シフトを決定する際の相互作用を強調している(例:Akacem er al 2016; Burgess and Eastman 2004)。他の行動因子には、朝の光への自発的な曝露が含まれる。例えば、朝の光を積極的に回避することは、夜勤労働に対する概日リズムのより良い適応と関連していた(Koller et al 1994)。
SM 3に記載されているが、選択された出版物の中には含まれていない睡眠脳波を用いた研究については、夜間の光曝露は、就寝後のフラッシュ光の連続的な曝露である場合を除き、翌晩の睡眠脳波の構造を変化させることが報告されている(SM 3参照)(Zeitzer er al)。 しかし、その影響は小さいようである。例えば、実験的に誘発された約1時間半の位相シフトは、健康な個体において、その後の睡眠傾向およびアーキテクチャに及ぼす影響はわずかであった(Carrier and Dumont 1995)。誘発された変化は、多くの場合、一晩中の睡眠の変化したコースと関連している。例えば、夜間に緑色と対照的に青色の光を浴びると、レム睡眠時間が短縮され、睡眠の最初の数時間は遅波睡眠が抑制され、早朝に向けてリバウンドする(Munch er al 2005)。注目すべきは、個人のメラトニンレベルも光に対して非常に異なる反応を示すことがわかったことである。
概日リズムに関連した強度とスペクトル特性に関する表示
サーカディアンの位相や夜間睡眠の望ましくない変化を避けるためには、最長波長(631nm)や間欠的な光曝露でさえも概日リズムのリセット反応を誘発するため、夜間や夜間、朝の光曝露を制御する必要がある。サーカディアン反応は、夜間の光量が低い家庭環境で目を閉じて寝ているときにも見られ、光量が低くても人間の概日リズムに関係していることが示された。
研究デザイン
レビューされた研究は、研究デザインや光曝露の詳細を提供する方法論が非常に多様であった。実験研究の一部には、暗い光、薄暗い光、またはその他のコントロールレベルの光にさらされた対照群が含まれており、他の研究では、同じ被験者が光と対照の両方の条件にさらされた反復測定実験が行われてた。研究はまた、様々な方法で事前の光曝露を指示し、制御した。事前の光曝露は、せいぜい実験の数週間前から携帯型光測定装置を用いて制御および/または測定されたが、いくつかの研究では事前の光曝露を全く測定していないという欠点があった。短期間の光曝露を特別に測定することに加えて、自然の季節的または行動パターンに起因する光環境への被験者の適応が、光曝露に対する感度に影響を与える可能性がある(Wehr 1991)。
メラトニンの測定
研究では、血液、唾液、尿サンプルからメラトニンレベルを評価した。青木ら(1998)は、唾液の採取は被験者にとって侵襲性が低いため、血漿(血液)よりも唾液の採取を好んだが、唾液メラトニンは概日相評価のために有効であることが確認されている(Voultsiosら(1997))。メラトニンの一次尿中代謝物である6-スルファトキシメラトニンは、より長い時間にわたってメラトニン産生を推定するために使用することができるが、より正確で頻繁なメラトニンレベルの評価のためには、唾液採取が推奨される。DLMOは血液と唾液のサンプルからしか推定できない。
多くの研究は、被験者間のメラトニンパターンの個人差を発見し(例えば、Ho Mien er al 2014; Kozaki er al 2016; Sander er al 2015)被験者は個々のメラトニンレベルと光曝露に対する反応を持っていることを示唆している。メラトニンレベルの違いに加えて、樋口 et al 2008)は瞳孔の大きさに大きな個体間差があることを発見した。レンズの特性が加齢によって変化することから、年齢が光に対する感度に影響を与えることは論理的に考えられる(例:Benloucif er al 2006; Münch er al 2011; Sletten er al 2009)。
一般的な方法論の限界
文献レビューの128件の研究から収集したデータの範囲を考えると、研究の中にはいくつかの制限がある。第一に、多くの研究でサンプルサイズが小さかった。128件中39件の研究では10件以下のサンプルサイズを使用していた。第二に、瞳孔の大きさが管理されているか、または測定されている研究は非常に少数であった。瞳孔拡張は瞳孔サイズをコントロールする方法である。典型的な生活環境における光曝露の影響に関心がある場合には、結果を誇張する可能性があるため、瞳孔を拡張すべきではない。第三に、多くの研究では光量がどこで測定されたかが示されていない。光量は、目が開いているときの目のレベル、つまり角膜で測定すべきである。第四に、一部の研究では、曝露中の眼の状態が明確に記載されていなかった。眼瞼はレッドパスフィルターとして機能し(Zeitzer er al 2014)波長依存的に光の約3%〜14%しか透過しない(Robinson er al)。 1991)。したがって、光の網膜露出は、目の状態(開いている、閉じている)に依存する。第五に、多くの研究は、正確な方法で被曝のスペクトル特性を示すことができなかった。そのためには、SPD、波長、または帯域幅のあるピーク波長の提示が必要である。全体として、年齢、眼の状態、瞳孔の大きさ、露光前のベースライン期間の長さとパターン、睡眠パターン(睡眠不足または習慣的な睡眠リズム)などの被験者の資質とともに、露光の強度、スペクトル、時間的特性を正確かつ明確に示すことが必要である。
今回のレビューの限界
今回のデータ収集では、レビューされた研究では多くの情報が非常に矛盾して報告されていたため、いくつかの制限が確認された。第一に、姿勢がメラトニンレベルに影響を与えることが示唆されているにもかかわらず、メラトニンサンプル収集中の被験者の姿勢に関する情報が収集されていないことである(Deacon and Arendt 1994)。第二に、実験のタイミングが収集されていない。これは、光の季節変動が血漿メラトニンパターンに影響を与えることが報告されているため、被験者の光レベルへの適応に影響を与える可能性がある(Arendt er al)。 1979; Touitou er al)。 1984)。第三に、女性被験者の月経周期と経口避妊薬の潜在的摂取に関する情報は収集されていない。経口避妊薬の使用は夜間のメラトニンレベルを増加させることがわかっており、月経期はメラトニンレベルに影響を与える可能性がある(Webley and Leidenberger 1986; Wright and Badia 1999)。第四に、ベースライン期間または対照期間の長さは収集されなかった。これは、事前の光曝露に関係している。
さらなる研究
この系統的な文献レビューの結果は、LCA手法における光害の人間の健康に関連する影響カテゴリーの開発に利用することが推奨される。従来の LCA 法で評価されたエネルギー関連フローや物質関連フローの環境影響と並行して、照明製品のより広い持続可能性のコンテクストを評価することが可能になる。光害のカテゴリーは、まず、光による概日リズムへの影響に焦点を当てるべきである。しかし、概日リズムの乱れは健康へのさらなる影響をもたらす可能性があるため、これらのさらなる健康影響を評価すべきである。最も重要なことは、夜間の光および夜勤労働は、がんリスクの増加と関連していることである(Straif er al)。 生理的メカニズムは不明のままであるが、メラトニンの抑制、概日リズムの乱れ、慢性的な睡眠不足など、いくつかのことが示唆されている(Haus and Smolensky 2013)。癌リスクの増加の可能性に加えて、メラトニン分泌の低下は、2型糖尿病の高いリスクと関連している(McMullan er al)。
本レビューに挙げた研究の限界を考えると、光による概日リズムの影響に関する今後の研究では、実験プロトコルの詳細と被験者を明確に示すべきである。多くの研究でメラトニンレベルに個人差が見られたため、サンプルサイズと反復測定計画には特に注意を払わなければならない。いくつかのレビューされた研究では、光の概日反応のスペクトル感度に起因する非視覚光受容体の役割が確認されたが、視覚光受容体(錐体)の潜在的な役割を無視することはできず、さらなる研究の必要性を示唆している。さらに、人間の健康に対する光の影響に関する科学的証拠をさらに明確にするために、他の光によって誘発される健康上の懸念に関するシステマティックレビューや、概日リズムの乱れを含む健康への悪影響に関するメタアナリシスが必要である。