
Contents
Novel Therapeutics for Epstein–Barr Virus
www.ncbi.nlm.nih.gov/labs/pmc/articles/PMC6429425/
オンライン2019 Mar 12掲載
概要
エプスタインバーウイルス(EBV)は、成人人口の95%が感染するヒトγヘルペスウイルスである。EBVの初感染は通常、小児期に起こり、一般的には無症状であるが、後年になって感染すると、35〜50%の症例で伝染性単核球症を引き起こすことがある。EBVは主にB細胞と上皮細胞に感染し、休眠状態の記憶B細胞に潜伏し、場合によっては上皮細胞にも潜伏する。EBVは発癌性ウイルスとして認識されているが、免疫力の高い宿主では、生体内での形質転換を防ぐ免疫反応によってEBVの再活性化が制御される。免疫抑制下では、原因にかかわらず、免疫系がEBVの複製を制御できなくなり、その結果、新生物が出現することがある。EBVに関連する主要な悪性腫瘍はB細胞リンパ腫と鼻咽頭癌であり、これは生体内でのウイルス感染の主要な標的となる細胞を反映している。
いくつかの抗ウイルス剤が試験管内試験でEBVの複製を阻害することが証明されたが、臨床での成功は限られており、今日までEBV感染症の治療に承認された抗ウイルス剤はない。ここでは、EBV感染症の治療のために臨床で評価されている抗ウイルス剤をレビューするとともに、現在研究中の抗EBV活性を持つ新規分子や、EBV関連疾患の治療のための新しい戦略について議論する。
キーワード エプスタインバーウイルス、抗ウイルス剤、ヌクレオシド類似体、ヌクレオチド類似体、細胞標的
1. はじめに
ヒトγ-ヘルペスウイルスであるエプスタイン-バーウイルス(EBVまたはヒトヘルペスウイルス4,HHV-4)は、最もよく感染するヘルペスウイルスの一つであり、成人のヒト人口の95%までが感染していると言われている。EBVの初感染は一般的に小児期に起こり、通常は無症状である。しかし、思春期や成人期初期にEBVに感染すると、35%から50%の割合で伝染性単核球症を発症することがある。伝染性単核球症の症状(疲労感、発熱、喉の炎症、首のリンパ節の腫れ、脾臓の肥大、肝臓の腫れ、発疹)は、通常1~2ヵ月で治まるが、伝染性単核球症が治った後の青年期の慢性疲労症状の発生率は、6ヵ月後、12ヵ月後、24ヵ月後に、それぞれ13%、7%、4%となっている[1]。
EBVは、他のヘルペスウイルスと同様に、一次感染後、生涯にわたって潜伏する。EBVの感染は免疫系によって制御されているため、生涯にわたってEBVを保有する人のほとんどは、ウイルス感染に悩まされることはない。しかし、EBVは、免疫抑制状態(先天的なもの、HIV感染に伴うもの、移植や自己免疫疾患における免疫調整剤の使用に伴うもの)では、重篤な急性疾患や、リンパ系や上皮細胞由来の様々な生命を脅かす悪性腫瘍を引き起こす可能性がある。EBVは、バーキットリンパ腫などの非ホジキンリンパ腫、ホジキンリンパ腫、中枢神経系リンパ腫、移植後リンパ増殖性障害(PTDD)エイズ関連リンパ腫、ナチュラルキラー(NK)リンパ腫、T細胞リンパ腫などのB細胞悪性腫瘍の発症に関連している[2,3](表1)。また、非角化性上咽頭癌の100%に見られ、消化管癌との関連も散発的に認められている[4,5]。
表1 免疫力のあるホストと免疫力のないホストにおけるEpstein-Barr(EBV)関連リンパ腫
| 免疫担当ホスト | 免疫不全の宿主 | ||||
|---|---|---|---|---|---|
| リンパ腫 | EBV協会 | レイテンシープログラム | リンパ腫 | EBV協会 | レイテンシープログラム |
| BL(風土病) | 100% | IまたはWp-制限付き | PTLD、B細胞 | > 90% | III |
| BL(散発的) | 15〜85% | 私 | BL(HIV) | 25〜35% | 私 |
| クラシックHL | 40% | II | HL(HIV) | > 80% | II |
| 慢性炎症に関連するDLBCL | 〜70% | II | PEL(原発性滲出液リンパ腫) | > 80% | 私 |
| 高齢者のEBV陽性DLBCL | 100% | II | 形質芽細胞リンパ腫 | 〜70% | IまたはII |
| リンパ腫様肉芽腫症 | 100% | II | 形質芽細胞リンパ腫、口腔型(HIV) | 100% | 私 |
| 血管免疫芽球性T細胞リンパ腫* | > 90% | II | 原発性CNSリンパ腫(HIV) | 100% | III |
| 節外性NK / T細胞リンパ腫、鼻型* | 100% | II | 一次免疫障害のあるNHL | > 90% | III |
| アグレッシブNK細胞白血病* | > 90% | II | 医原性免疫不全リンパ腫 | 40〜50% | III |
| PTLD、NK / T細胞* | > 70% | III | |||
[2,3,4,5]から引用している。* EBV関連のT細胞およびNK細胞リンパ腫。DLBCL:びまん性大細胞型B細胞リンパ腫。散発性バーキットリンパ腫(BL)では、EBVとの関連性の強さは地域によって異なり、そのため報告されている割合の幅も広い。
EBVのライフサイクルには、溶解期と潜伏期という2つの異なるフェーズがある(図1)。EBVは、CD21受容体を介してB細胞に優先的に感染するが、低い頻度で上皮細胞やTまたはNK系列の細胞にも感染する。ウイルスは上皮細胞で溶血性複製を行い、循環しているメモリーBリンパ球に生涯潜伏し、定期的に潜伏から再活性化する[4,6]。溶血性感染の際には、ウイルスの遺伝子発現の全レパートリーが行われ、子孫ウイルスが産生される。EBVの2つの重要な即時型溶解遺伝子(すなわち、BZLF1とBRLF1)は、ウイルスや特定の細胞のプロモーターを活性化するトランジベーターをコードしており、初期の遺伝子発現とゲノム複製に続いて後期の遺伝子発現という、ウイルス遺伝子発現の秩序立ったカスケードを引き起こしている。上皮細胞内での溶解複製で生成されたウイルスは、宿主内や宿主間でのウイルス粒子の散布を可能にする。
図1 EBVのライフサイクル、潜伏期と派生するリンパ腫。
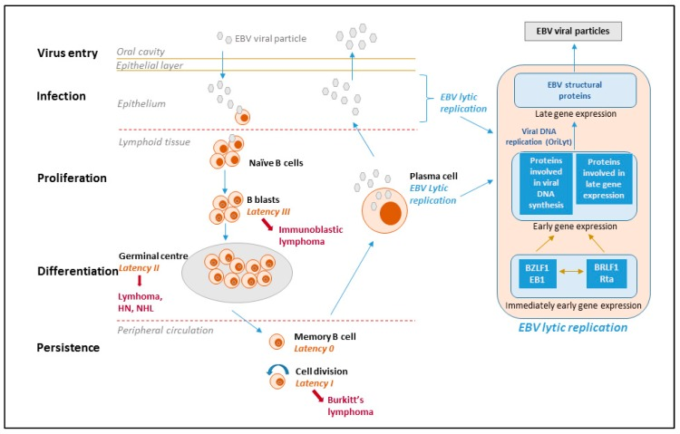
ウイルスのライフサイクルには、少なくとも5つの異なるステージ(ウイルスの侵入、感染、増殖、分化、持続)があり、そのうち4つのステージがEBV疾患に関連している。ウイルスは唾液を介して感染し、中咽頭粘膜のナイーブなB細胞に感染する。一次感染時、EBVに感染したナイーブB細胞は、潜伏遺伝子複合体(10種類のタンパク質)を全て発現する。潜伏遺伝子複合体(EBV核抗原(EBNA)潜伏膜タンパク質(LMP)の10種類のタンパク質)と、EBVにコードされた低分子RNA(EBER)やマイクロRNAを発現している。これはIII型潜伏と呼ばれ、この形態の潜伏は、休眠中のB細胞を活性化し、増殖や形質転換へと駆り立てる。しかし、これらの細胞は免疫原性が高く、EBV特異的T細胞によって速やかに排除される。ウイルスがB細胞内で生存できるのは、免疫原性タンパク質をダウンレギュレートしているからである。EBVは抗原駆動型のB細胞反応を模倣しており、抗原で刺激された芽球と同様に、EBVに感染したB細胞は毛包に入り、膨張して胚中心を形成し、3つのウイルスタンパク質のみを発現する(II型潜伏)。最終的には、1つのウイルスタンパク質(EBNA1,ウイルスゲノムが細胞ゲノムと一緒に分裂することを保証する)のみを発現してリンパ節を出る(I型潜伏)。EBVに感染した細胞が末梢血に入ると、タンパク質をコードするすべてのウイルス遺伝子が停止する。これを潜伏期0,またはウイルスタンパク質が発現していない潜伏期プログラムと呼ぶ。ウイルスが静止している休眠記憶細胞は、宿主の免疫系に攻撃されることはなく、長期的な存続の場となっていると考えられる。メモリーB細胞は、安定した細胞数を維持するために時折分裂し、ウイルスを保有する細胞が分裂すると、ウイルスのEBNA1タンパク質が発現して、ウイルスゲノムが細胞とともに複製されるようになる。また、記憶B細胞は、プラズマ細胞への終末分化を行い、抗体を分泌することがある。このような細胞にウイルスが含まれていると、EBVの溶解プログラムが活性化され、形質細胞から放出された感染性ウイルスが上皮細胞に感染し、そこでウイルスが複製され、大量に排出されて、他の宿主に感染することがある。潜伏タイプ0を除いて、各潜伏状態は特定のタイプのEBV関連悪性腫瘍に見られる。
潜伏期間中、ウイルスは、ウイルスゲノムの維持(核内のエピソームとして)や宿主の免疫系の回避に必要な限られた遺伝子のみを発現している。潜伏遺伝子の発現に基づいて、EBVの潜伏は異なるタイプに分類される(図2A)[7]。潜伏期0,I、II、IIIという命名法は、遺伝子発現のスナップショットを表していることに注目してほしい。EBVの潜伏期間は、潜伏0(健常者の循環メモリーB細胞で観察されるように、EBV抗原が発現していない状態)から潜伏III(PTLDで見られるように、すべての核タンパク質(EBNAs -1,-2, -3A, -3B, -3C, -LP)と2つの膜タンパク質(LMP1, LMP2)が2つの低分子RNA(EBER)とともに発現している状態)まである。腫瘍によっては、これらの潜伏パターンのいずれにも当てはまらない場合があり、さらに、免疫組織化学的には1つの生検内でも発現の不均一性が認められることがある。さらに、EBV関連の悪性腫瘍では、細胞内の遺伝子変化や共同感染が生じる(図2B)。EBVを原因とする腫瘍では潜伏プログラムが優勢であるが、溶解性ウイルスの複製も病原上重要である[8,9]。
図2 (a) EBV潜伏期の遺伝子発現パターン。
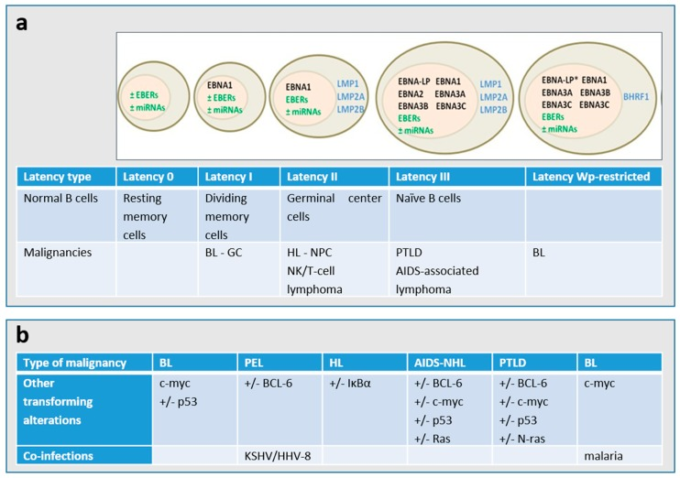
パンデミック性BLの大部分は潜伏I型を呈し、野生型の形質転換コンピテントEBVゲノムを持ち、EBNA1特異的潜伏プロモーターQpからのEpstein-Barr核抗原1(EBNA1)非コードEBER(Epstein-Barr virus-encoded small RNA)およびいくつかのマイクロRNA(miRNA)のみを発現している。BLパンデミック性腫瘍の約15%は、いわゆるWp-restricted BLと呼ばれ、EBNA2遺伝子が欠失したゲノムを持ち、Wp潜伏プロモーターからEBNA1,-3A、-3B、-3CおよびウイルスBcl2ホモログのBHRF1を発現している[2,6]。* Wp制限潜伏では、EBNA-LP遺伝子が部分的に欠失している。EBV関連悪性腫瘍における潜伏の主なタイプは、潜伏I遺伝子に加えて潜伏膜タンパク質LMP1,LMP2A、LMP2Bが発現している潜伏IIである。複数のEBNAタンパク質、LMP1,LMP2A、LMP2B、EBER、miRNAからなるEBV潜伏遺伝子複合体全体が発現しているのが潜伏III型である。(b) EBV関連悪性腫瘍の異なるタイプでは、細胞の遺伝子変化および/または共同感染が起こることが知られている。PEL:原発性滲出液リンパ腫、HL:ホジキンリンパ腫、BL:バーキットリンパ腫、NHL:非ホジキンリンパ腫、PTLD:移植後リンパ増殖性障害、NPC:鼻咽頭がん、GC:胃がん。
2. なぜ、EBV感染症の治療に承認された抗ウイルス剤がないのか?
ヌクレオシド系(アシクロビル(ACV)ペンシクロビル(PCV)ガンシクロビル(GCV)およびその経口プロドラッグであるバラシクロビル(VACV)ファムシクロビル(FAM)バルガンシクロビル(VGCV)など)ヌクレオチド系(シドホビル(CDV)など)ピロリン酸系(フォスカビル(foscarir)など)の抗ウイルス剤がある。foscavir (foscarnet sodium, PFA))類縁体は、単純ヘルペスウイルス1型(HSV-1)および2型(HSV-2)水痘帯状疱疹ウイルス(VZV)および/またはヒトサイトメガロウイルス(HCMV)の治療薬として承認されている[10,11]。欧州の一部の国では,ブリブジン(BVDU)がHSV-1およびVZV関連疾患の治療薬として承認されている。これらの抗ウイルス剤の中には,試験管内試験でEBVの複製を阻害する効果があることが証明され,実験的に使用されたものもあるが[11,12,13],EBV感染症の治療薬としてFDA(米国食品医薬品局)やEMA(欧州医薬品庁)の承認を受けたものはなかった。
2005,GershburgとPaganoは、抗EBV薬が存在しない理由として、主に3つの説明を提案した[14]。まず、EBV関連感染症の治療薬の開発が成功していないのは、感染性単核球症の診断の難しさが、少なくとも部分的には原因となっている可能性がある。EBVは30歳になるとほとんどの人が感染するが、感染性単核球症を発症する人はごく少数である(通常は20代で感染した人)。伝染性単核球症の症状は、αヘルペスウイルスであるHSV(口唇ヘルペス)やVZV(水痘)による感染症とは対照的に、発症が微妙であり、潜伏期間も長い(4~6週間)ため、診断が遅くなる。
第二に、抗ウイルス剤は、EBVが高力価で放出される中咽頭で高濃度を達成する必要がある。アシクロビルを静脈内および経口投与したところ、中咽頭でのEBV排出量が有意に減少したことが示されたが、治療を中止してから3週間以内にウイルス放出が初期レベルに再開した[15,16]。伝染性単核球症の治療に抗ウイルス剤が使えない最も重要な理由は、この病気の症状や兆候がウイルスの複製の結果ではなく、血液中を循環し、さまざまな臓器の組織に浸潤しているEBV感染B細胞に対する免疫学的反応であるという事実に起因するのかもしれない。伝染性単核球症は、ウイルスに感染したBリンパ球に対する大量の細胞介在性免疫反応により、非定型リンパ球症を特徴とする。そのため、抗ウイルス剤と免疫調整剤(コルチコステロイドなど、医師が経験的に伝染性単核球症の治療に使用しているもの)を併用することが有効であると考えられる。しかし、多施設共同二重盲検プラセボ対照試験では、感染性単核球症の治療のためにプレドニゾロンをアシクロビルと一緒に投与したところ、臨床症状の期間やEBV特異的細胞性免疫の発達に影響を与えることなく、口腔咽頭のEBV複製が抑制された[16]。
3. リチン複製を標的とした抗EBV治療薬の医学的必要性
EBVの初感染は通常無症状であるが、一部の患者は感染性単核球症を発症し、その症状は発熱、咽頭痛、リンパ節腫脹などの軽度のものから、免疫力の低い宿主では致命的なものまである。さらに、感染性単核球症を伴う、あるいは伴わないEBVの一次感染は、合併症(自己免疫性溶血、扁桃腺肥大による気道閉塞、脾臓破裂、脳炎、重症肝炎、心筋炎など)を引き起こす可能性があるが、これらは主にウイルスに対する免疫病理学的反応の結果として起こる。その他、まれではあるが、重篤な合併症(無顆粒球症や再生不良性貧血など)が健康な患者に生じることもある。伝染性単核球症に関連する合併症の治療には、主にコルチコステロイドが用いられる。重度の伝染性単核球症の合併症の管理における抗ウイルス剤の役割は、ケースシリーズに基づいて議論の余地があるが、医師はEBV感染症の重篤な症状に抗ウイルス剤の使用を検討することができる。伝染性単核球症の病因は主に免疫を介したものであるにもかかわらず、EBV関連肝炎の重症度はウイルス負荷の高さと関連していることが示された[17,18]。そのため、特定の抗ウイルス剤を使用することで、感染性単核球症に見られるEBV関連の合併症の症状を緩和することが期待されている。
EBVの溶血性複製は、感染性単核球症に直接関連するだけでなく、慢性活動性EBV感染症(CAEBV)や口腔毛髪性白板症にも関連している。CAEBVはアメリカやヨーロッパではまれであるが、アジアや南米では頻繁に発生する。CAEBVは、EBV特異的抗体が著しく上昇し、ウイルス量が多いリンパ増殖性疾患である。CAEBVは、長期にわたる慢性または再発性の伝染性単核球症のような症状と、異常なパターンの抗EBV抗体によって特徴づけられる、しばしば致命的な疾患である。生命を脅かすCAEBVの合併症には、ウイルス関連の血球貪食症候群、白血病、T/NK細胞系のリンパ腫などがある[19]。すぐに初期の遺伝子の発現をアップレギュレートすることによって溶解サイクルを再活性化する遺伝子内EBV欠失は、ウイルスの産生およびリンパ腫形成の促進を回避することに関連していた[20]。CAEBVの治療には、免疫調整剤(インターフェロン-αやインターロイキン-2など)抗ウイルス剤(アシクロビル、ガンシクロビル、ビダラビンなど)化学療法剤、EBV特異的細胞傷害性Tリンパ球を用いた細胞療法、造血幹細胞移植などが用いられてきたが、その成果は限定的であった[21]。
移植後リンパ増殖性障害(PTLD)は、固形臓器移植後の重篤でしばしば致命的な合併症であり、EBVはこのグループの患者におけるPTLDの主要な危険因子であり、EBV未感染の患者が血清反応を起こした場合、30%から50%がPTLDと診断される[22,23]。PTLDは、同種造血幹細胞移植(HSCT)後の生命を脅かす疾患でもあり、薬剤による免疫モニタリング機能の低下が原因で、リンパ球が無秩序に増殖することで起こる。PTLDの発症率は、過去20年間で著しく増加している。これには、移植件数の増加、ドナーおよびレシピエントの年齢上昇、強力な新しい免疫抑制剤の使用、ハプロアイデンティティー造血幹細胞移植の導入、疾患に対する認識の向上、診断ツールの改善など、いくつかの理由が挙げられる[24]。高いウイルス量が持続することやウイルス量の増加は、PTDDのリスク増加と関連している[25,26]。PTLDでは、ウイルスが潜伏しているため、抗ウイルス療法は有用ではないと考えられてきた。しかし、抗ウイルス剤を予防的に投与することで、PTLDの発症率が低下した[27,28,29]。固形臓器移植を受けた人の血液中のEBV DNAレベルが高いと、PTLDの発症を予測することが示されているため、抗ウイルス剤はPTLDの予防に役立つかもしれない。早期発症のPTLD(移植後1年以内に発症)のほとんどの症例は、最近のEBV感染と関連している。移植後1年目以降に発症する遅発性リンパ腫は、EBVとの関連性が低いとされている。
EBVの溶血性感染がリンパ増殖性疾患に寄与することが示されているため、溶血性タンパク質の発現を抑制することを目的とした治療戦略は、EBV関連悪性腫瘍の初期段階の制御に役立つはずである[8,30]。EBVのIEタンパク質であるBZLF1とBRLF1は、初期のリンパ増殖性疾患を促進する球状感染細胞におけるIL-6の分泌に寄与している[31]。IL-6は、免疫機能の維持、造血細胞の分化促進、および炎症の永続化に重要な役割を果たすことが知られているサイトカインであるが、様々な血液および上皮性悪性腫瘍の重要な因子でもある。IL-6は、パラクラインおよびオートクライン機構を介して作用し、細胞の生存を促進するとともに、signal transducer and activator of transcription 3 (STAT3)を誘導する。したがって、上皮細胞とリンパ系細胞の両方に感染できるウイルス(EBVなど)が、IL-6の発現を誘導するメカニズムを持っていることは驚くべきことではない。また、細胞に感染した細胞は、細胞性およびウイルス性のIL-10 [32]の発現を誘導し、B細胞がより効率的に成長することを可能にし、また、B細胞および上皮性悪性腫瘍の両方において血管新生に寄与するVEGF [33]の発現を誘導する。
4. 臨床で評価されているEBVに対する抗ウイルス剤
4.1. ヌクレオシド系抗生物質(アシクロビル、バラシクロビル、ガンシクロビル、バルガンシクロビル)
アシクロビルとガンシクロビルは試験管内試験でEBVを阻害することから[11,34,35]、これらの薬剤とその経口プロドラッグについて、免疫抑制中のEBV再活性化の抑制を評価した。EBV血清陰性の肺および心肺移植を受けた患者のPTLD発症率は、アシクロビル、バラシクロビル、ガンシクロビルによる抗ウイルス剤の予防により減少した[28]。このリンパ増殖性疾患の発生率を1996年以前(歴史的グループ)と1996年から 2000年の間(抗ウイルス剤の予防投与を受けたグループ)で分析し、長期の抗ウイルス剤の予防投与がEBVゼロネガティブなレシピエントのPTDD発症に与える影響を比較した。その結果、継続的に抗ウイルス剤の予防投与を受けたEBVゼロネガティブ患者ではPTLDが発症しなかったのに対し、歴史的群では4.2%の患者にPTLDが発症した。
EBV陽性のドナー(D+)からグラフトを受けたため、EBV一次感染のリスクがあるEBV未発症の小児腎移植患者(R-)を対象に、ガンシクロビルおよびバルガンシクロビルの予防投与がEBVウイルス量に及ぼす影響を評価した[27]。移植後1年間、ガンシクロビルまたはバルガンシクロビルによる抗ウイルス剤の予防投与は、EBV一次感染の発生率を有意に減少させた。抗ウイルス剤の予防投与は、EBVのウイルス量を有意に低下させる一方、免疫抑制療法の種類や強度は、EBVの一次感染の発生率やウイルス量のレベル/持続性に影響を与えなかった。
2000年1月から 2016年1月の間に移植された、成人の腎臓または腎膵臓のEBV血清陰性レシピエント73人の単調なレトロスペクティブコホートにおいて、HCMV疾患を予防するために使用される抗ウイルス剤の影響が調査された[36]。37人(50.7%、予防群)がバラシクロビルまたはバルガンシクロビルを3~6カ月間投与され、36人(49.3%、予防なし群)が抗ウイルス剤を投与されず、平均追跡期間は69カ月(予防群)91カ月(予防なし群)であった。予防投与は,EBV ウイルス量のモニタリングにより,100 日後の一次感染を遅らせた(43%対 84%).早期PTLDの発生率は両群間で差がなかったが、EBV関連新生物の発生率は、予防を行わなかった群(6例の新生物が報告されている)に比べて、治療を受けた患者(1例も観察されなかった)で有意に低かった。エビデンスレベルは弱いものの、抗ウイルス剤の予防投与は遅発性PTLDを予防できる可能性がある。
小児の肝移植患者のコホートでは、ガンシクロビルの静脈内投与による治療は、EBVウイルス血症の検出から8週間後および1年後にEBV負荷が減少した患者の割合に変化はなかった[37]。2002年から 2015年までノルウェーで行われたこのレトロスペクティブ・スタディでは、EBVウイルス血症の患者38人が含まれ、そのうち32人が中央値22(21~38)日の間、ガンシクロビルの静脈内投与を受けた。移植からウイルス血症までの期間が短いこと、移植時の年齢が若いこと、移植前にEBVのセロコンバージョンがないことは、慢性的なEBVウイルス血症の有意な予測因子であった。
バルガンシクロビルは,HCMV と EBV の再活性化を起こしやすいアレムツズマブ(すべての B リンパ球と T リンパ球に発現する CD52 に対するヒト化モノクローナル抗体)による免疫抑制を受けている患者 29 名のグループにおいて EBV の再活性化を抑制した[38]。血漿中のEBV DNA量は,29人の患者(258検体,中央値は1人あたり7検体)で定量化された。24人の患者では、定量可能なEBV DNAは検出されず、5人の患者(17%)ではEBVの再活性化が見られたが、4人の患者では自然に減少した。また、他の強力なT細胞抑制剤(フルダラビン)の投与を受けていた1名の患者が、EBV陽性のホジキンリンパ腫を発症した。
免疫抑制剤を減少させないバルガンシクロビルの有効性と安全性は、血液中に持続的なEBV DNAを示した肝移植を受けた子供たちのグループで評価された[29]。抗ウイルス剤の長期投与(中央値8か月)を受けた患者の20/42(47.6%)にウイルス量の検出不能が認められ、そのうち60%が治療に対する反応を維持した。しかし、本研究では対照群がないため、結果の解釈には慎重を期す必要がある。
EBVの経口排出に対するバルガンシクロビルの効果は、無作為化二重盲検プラセボ対照試験で評価された[39]。対象となったのは26名の男性で、参加者全員が男性と性交渉を持つ男性であることを自認し、16名(62%)がHIV-1に感染していた。彼らは,バルガンシクロビルまたは毎日のプラセボを8週間経口投与し,その後,2週間の「ウォッシュアウト期間」を経て,代替療法を8週間行った。バルガンシクロビルは、EBVが検出された日の割合を61.3%から17.8%に、検出されたウイルス量を0.77ログにそれぞれ有意に減少させたことから、EBV関連疾患に対するバルガンシクロビルの影響をさらに調査する必要があると考えられる。
4.2. ヌクレオチド類縁体
ヌクレオチドアナログのシドフォビル(CDV)は、幅広いスペクトルを持つ抗DNAウイルス剤である。この化合物は,AIDS患者の網膜炎の治療に承認されているが,DNAウイルスに起因するいくつかの感染症の治療にも非標識で使用されている[40]。抗ウイルス作用が認められているほか、抗増殖作用があることも知られている[41]。
抗ウイルス剤であるシドフォビルを用いた局所再発EBV関連鼻咽頭癌の治療が2名の患者で成功したことが報告されている[42]。さらに,ヌードマウスのEBV陽性上咽頭癌異種移植片の腫瘍組織にシドホビルを注射すると,腫瘍の成長が抑制された[43].リボヌクレオチド還元酵素阻害剤であるヒドロキシウレアおよびジドックス(3,4-ジヒドロキシベンゾヒドロキサム酸)との併用により,EBVに形質転換した上皮細胞およびEBV陽性の鼻咽頭癌異種移植片におけるシドホビルによるアポトーシスが増強された[44].シドフォビルはEBVのオンコプロテインを減少させ、EBV関連悪性腫瘍(バーキットリンパ腫、鼻咽頭癌)の放射線感受性を高めた[45]。
4.3. ピロリン酸類似体
無機ピロリン酸アナログであるフォスカーネットは、ヘルペスウイルスのDNAポリメラーゼを直接阻害する。ピロリン酸結合部位を阻害し,デオキシヌクレオシド三リン酸からのピロリン酸の切断を阻止することができる。フォスカーネットは、ウイルスのDNAポリメラーゼを非競合的に阻害する薬剤で、成長するウイルスのDNAには組み込まれず、細胞の酵素よりもウイルスの酵素に対して100倍以上の活性を持つ。EBVを含むすべてのヒトヘルペスウイルスに対して活性を有するが、フォスカーネットはAIDS患者におけるHCMV網膜炎の治療や、免疫不全患者におけるアシクロビル耐性HSV感染症の治療に承認されている。また、UL97プロテインキナーゼ(PK)の変異を原因とするガンシクロビル耐性HCMV感染症の治療にも使用されている。その他のヘルペスウイルス感染症に対する安全性と有効性は、まだ確立されていない[10]。EBVの持続感染にフォスカーネットを使用して成功したという報告が時々ある。フォスカーネットと免疫グロブリンを併用することで、免疫抑制剤の強度を下げた後にPTDDの臨床的改善を示した肺移植患者のEBVの持続感染を制御することに成功した[46]。この患者では、細胞性免疫の回復によりPTLDは改善したが、EBV感染の制御には効果がなかったため、抗ウイルス剤と免疫グロブリンによる治療が必要であった。また、2人のAIDS患者において、フォスカーネットによる治療中にEBV関連のリンパ増殖性疾患が退縮したことが報告されている[47]。
5. 研究中の抗EBV化合物
5.1. EBVプロテインキナーゼBGLF4の阻害剤
マリバビル(MBV)は,HCMVとEBVの両方に有意な活性を示すが,他のヒトヘルペスウイルスには作用しない,経口ベンズイミダゾールL-リボシドの治験薬である[48,49]。マリバビルは、ウイルスのDNAポリメラーゼを標的とする抗HCMV薬と比較して、副作用が少なく、特異性が高い。ヌクレオシドやヌクレオチドアナログとは異なり、マリバビルの阻害効果は主にHCMVとEBVのPKを阻害することで生じる[50]。この化合物は,HCMVのUL97 PKを選択的に阻害し(キナーゼ活性の直接阻害により判定),UL97遺伝子の一点変異がHCMVにマリバビル耐性を付与する[51]。UL97は,HCMVの脱出に重要な役割を果たすセリン/スレオニン特異的なPKである。UL97は,HCMVに感染した細胞において,いくつかのウイルスタンパク質や細胞タンパク質のリン酸化に必要である[50,52]。ガンシクロビルとマリバビルで選択された試験管内試験および生体内試験のUL97変異は異なるものであり,交差耐性をもたらさないことが判明したが[53,54],ガンシクロビルとHCMVに有効なメチレンシクロプロパンヌクレオシドアナログであるシクロプロパビルとの間には部分的な交差耐性が認められた[51,55]。しかし、Chouら[56]は、ガンシクロビルを投与された被験者で発見されたか、シクロプロパビルを用いて試験管内試験で増殖した後にUL97キナーゼ変異が発見され、3つの薬剤すべてに対して中~高レベルの耐性を示したことを報告した。また、HCMVのUL27遺伝子の変異により、マリバビルに対する低レベルの耐性(2~3倍)が生じることもある。UL27の機能は不明であるが、マリバビルの直接の標的ではないようである。
同種幹細胞移植を受けた患者のHCMV疾患予防のためのマリバビル予防投与の有効性は、第3相:二重盲検、プラセボ対照、無作為化試験で評価された[57,58]。HCMV感染症の管理を目的としたマリバビルの臨床開発は、試験デザインに関するいくつかの批判があったものの、同種幹細胞移植を受けた患者における主要評価項目(HCMV疾患の予防)を満たすことができなかったため、現在保留されている[59]。
マリバビルは、EBVに対しても顕著な活性を示し、ウイルスのDNA複製の阻害とウイルスの転写の阻害というEBVに対するユニークな二重の効果を持っている[14,60]。HCMVとは対照的に、EBVに対するマリバビルの活性は、EBVのPKであるBGLF4を直接阻害することによるものではなかった。実際、マリバビルの投与は、EBVのDNAポリメラーゼの処理因子であるBMRF1のリン酸化を阻害することが示された[49]。EBVのRNAにほとんど影響を及ぼさないアシクロビルとは異なり、マリバビルは複数のRNAの発現を阻害する。さらに,マリバビルの転写産物の阻害プロファイルは,PKの発現と活性がノックアウトされたEBV変異株が産生するものと類似しているように見えた[61]。このことから,マリバビルはEBVのPKには直接影響を与えないものの,BGLF4の阻害を通じてEBVの転写産物レベルに大きく影響を与えていることが示唆された[62]。EBVのBGLF4には少なくとも20のウイルス標的があることを考えると、マリバビルは下流の標的にも間接的に影響を与える可能性がある。
5.2. EBVのDNAポリメラーゼの阻害剤
新規のl-ジオキソランチミジンアナログである1-[(2S,4S-2-(ヒドロキシメチル)-1,3-ジオキソラン-4-イル)5-ビニルピリミジン-2,4(1H,3H)-ジオン(HDVD)は,HSV-1,KSHV,EBV,およびマウスヘルペスウイルス68(MHV-68)に対して活性があることが証明された[35]。この化合物は,HSV-2,VZV,サイミール・ヘルペスウイルス(HVS)の複製を弱く阻害し,HCMVとアカゲザル・ラジノウイルス(RRV)に対しては抗ウイルス活性が認められなかった。また、HSV-1やVZVに対して強力な抗ウイルス活性を持つことで知られる関連化合物brivudinとは抗ウイルス活性スペクトルが異なる。しかし、brivudinと同様に、HDVD耐性ウイルスの特徴から、HSV-1,MHV-68,HVSのウイルスのチミジンキナーゼ(TK)がこの化合物の活性化に必要であることがわかった。BALB/cマウスのMHV-68感染の鼻腔内モデルにおいて、HDVDおよびブリブジンの経口投与を評価した。HDVDの投与は、ブリブジンの投与とは対照的に、未投与のコントロールと比較して、ウイルスDNA量の減少と肺での急性ウイルス複製時のウイルス遺伝子発現の減少をもたらした。HDVDのバリルエステルプロドラッグ(USS-02-71-44)は,HVDよりも脾臓における潜伏感染の予防に効果的であった[35]。様々なヌクレオシド誘導体の耐性のメカニズムに関する研究では、ピリミジンヌクレオシド誘導体はγ-ヘルペスウイルスのTKによってリン酸化され、プリンヌクレオシドはγ-ヘルペスウイルスのPKによって優先的に活性化されることが示された[13]。これは、ガンシクロビルやアシクロビルが溶解性ウイルスの産生を阻害するためには、TKではなくEBVにコードされたPKが必要であるという以前の知見と一致する[63]。
2つのチオヌクレオシド誘導体,KAY-2-41およびKAH-39-149は,生体内試験(MHV-68マウスモデル)では有効な抗ウイルス効果を示し,試験管内試験では強力かつ選択的な抗EBV活性を示した[34]。また,これらの化合物はHSVおよびVZVに対しても活性を示した。KAY-2-41およびKAH-39-149耐性のHSVおよびMHV-68は,ウイルスのTKに変異を持っていたが,これらの変異は,他のTK依存性の薬剤と比較して,KAY-2-41およびKAH-39-149に対して低レベルの耐性しか与えなかった。HeLa TK欠損細胞を用いた抗ウイルスアッセイでは、HSV TK欠損変異株に対してKAY-2-41とKAH-39-149の活性が認められなかった。さらに,酵素アッセイでは,HSV-1およびVZVのTK,細胞のTK1およびTK2が,KAY-2-41およびKAH-39-149を認識してリン酸化することが示され,これらの化合物がウイルスと宿主の両方のTKに依存して抗ウイルス活性を発揮することが明らかになった。KAH-39-149は、γヘルペスウイルス感染モデルマウスにおいて、KAY-2-41よりも優れていることが証明され、EBVに対する抗ウイルス治療薬としての可能性が強調された。
様々なメチレンシクロプロパンヌクレオシド(MCPN)類似体は,細胞培養や動物モデルにおいて,いくつかのヘルペスウイルスに対して有効であることが証明された[64,65,66,67,68]。MCPN類縁体の最初のシリーズは,デオキシリボヌルセオシドの5′水酸基を模倣して,シクロプロパン環上に1つのヒドロキシメチル基を有していた。このシリーズの化合物は、アシクロビルと同様に、3′水酸基に対応するものがないため、DNAに取り込まれると、さらに鎖の伸長を阻害する。第2世代のMCPN類縁体は、ジヒドロキシメチル誘導体であり、ガンシクロビルと同様に、DNAポリメラーゼによって認識される第2の水酸基を持ち、さらなる鎖の伸長を可能にする。このクラスの最も活性の高い化合物であるシクロプロパビルは,HCMV,マウスCMV,EBV,HHV-6,HHV-8に対して良好な抗ウイルス活性を示し,そのプロドラッグである6-デオキシシクロプロパビルは,経口投与で良好な活性を示している[69]。CMVに対するシクロプロパビルの作用機序は,DNA合成の阻害とUL97 PKの両方が関与する複雑なものである[70]。(S)-シングアノールのような1つのヒドロキシメチル基を持つ最初のシリーズの類似体は,B型肝炎とHIVを含むより広い抗ウイルス活性スペクトルを示す。さらに最近では,6-エーテルやチオエーテル部位を持つモノヒドロキシメチルやジヒドロキシメチル類似体も,EBVを含むいくつかのヘルペスウイルスに対して活性を示すことが報告されている。これらの類縁体の中には,HSV-1,HSV-2,VZVを阻害するシクロプロパビルよりも広い活性スペクトルを示すものもあった[66]。これらの化合物の活性は,HCMVのUL97 PKに依存していたが,HSVのTK活性とは比較的独立していた。これらのデータは,これらの類似化合物の作用機序がシクロプロパビルのそれとは異なることを示しており,最終的には広域抗ヘルペスウイルス剤として使用できることを示唆している。
N-メタノカルバシミジンは,コンフォメーションロックされたヌクレオシドアナログであり,αヘルペスウイルス,γヘルペスウイルス,オルソポックスウイルスに対して活性を示した[71]。この化合物の抗ウイルス活性は,ウイルスのTKによる活性化に依存している。この化合物は,ウイルスのTKによって活性化されると,ウイルスのDNA合成を阻害する。この化合物は,オルソポックスウイルスおよびHSV感染の動物モデルにおいても有効であることが証明されており[72,73],安全性を評価するために健康なボランティアを対象とした第1相試験が進行中である[10]。
シドフォビル誘導体を含むいくつかの非環状ヌクレオシドホスホネート(ANP)は,EBVおよび他のγヘルペスウイルスの複製を高い効力と選択性で阻害した[12]。注目すべきは,ANPの環状プロドラッグは,EBV strain P3HR-1に対しては活性が低下したが,EBV strain Akataに対しては活性が低下しなかったことである。シドフォビルとその環状体(環状シドフォビル)を用いた代謝研究により、これらの違いは、EBV再活性化後のP3HR-1細胞における薬物代謝の変化、すなわち、環状CMPホスホジエステラーゼによる環状シドフォビルの加水分解の低下に起因することが明らかになった[12]。P3HR-1細胞におけるサイクリックAMPレベルの変化は、このサイクリックヌクレオチドがホスホジエステラーゼを競合的に阻害していることを示唆している。シドフォビルとその5-アザ誘導体(HPMP-5azaC)は,マウスモデルにおいて,マウスγヘルペスウイルスの複製と拡散を非常に効果的に阻害することが明らかになった[12]。
ブリンシドフォビル(CMX-001)は,経口投与可能なシドフォビルの製剤である。このシドホビルのアルコキシアルキルエステルプロドラッグは,試験管内試験ではシドホビルと同等の広域抗ウイルス活性を有するが,細胞内のシドホビル二リン酸レベルが高いため,シドホビルと比較して最大で1000倍の活性を有する[74]。ブリンシドフォビルは,その高い抗ウイルス活性に加え,シドフォビルとは対照的に,近位腎尿細管に存在するヒト有機アニオントランスポーター1酵素の基質ではないため,腎毒性を示さない。brincidofovirの前臨床データは有望であるにもかかわらず、同種造血幹細胞移植患者のHCMV感染予防を目的とした第3相試験では、残念な結果となり、市場への導入が遅れている。SUPPRESSと名付けられたこの試験では、ブリンシドフォビル群でプラセボ群と比較してHCMV疾患が増加したことが報告されたが、これはブリンシドフォビルの胃腸疾患が副腎皮質ステロイドで治療された移植片対宿主病と誤診されたことに関連している可能性がある[75]。これらの結果を受けて、HCMVの臨床試験は中止されたが、アデノウイルスの臨床試験は継続しており、IV製剤の開発も進められている。EBV関連疾患を対象とした本剤の評価試験は予見されていない[10]。
5.3. EBV核抗原1(EBNA1)の阻害剤
EBVにコードされた核抗原1 (EBNA1) は、EBVゲノムの維持、複製、および分離に機能する汎用性の高いタンパク質であり、EBV関連悪性腫瘍を治療するための魅力的な治療ターゲットとなっている。このタンパク質は、潜伏期0を除くすべてのEBV潜伏型で発現している。潜伏感染細胞におけるEBVエピソームゲノムの複製と持続は、主にEBVにコードされた核抗原1(EBNA1)が同族のEBVオリプ要素に結合することに依存している。
EBNA1の発現を低下させたり、その機能を阻害するための阻害剤の設計や同定には、ここ数年、かなりの努力が払われてきた。EBNA-1の顕著な特徴、その機能ドメインおよびEBNA-1阻害剤の開発における進歩については、最近、詳細にレビューされている[76]。例えば、Leeたち[77]は、EBNA1の低分子阻害剤(H20,H31)とその基礎となる阻害メカニズムを特徴づけた。H20は、in silicoのドッキング解析で予測されたように、EBNA1のDNA結合ドメイン(DBD)のポケットに収まったが、H20は、同族配列に対するEBNA1の結合には有意な影響を与えなかった。H20の限定的な構造関連性の研究により、疎水性化合物であるH31をEBNA1阻害剤として同定することができた。H31は、EBNA1依存のオリプの配列特異的なDNA結合活性を阻害したが、配列と非特異的な染色体の結合には影響を与えなかった。H31はEBNA1依存性の転写、複製、およびEBV oriPプラスミドの持続を抑制したが、これはEBNA1結合活性の阻害と一致する。重要なことは、H31はEBVのエピソームを徐々に消失させ、EBVに感染したリンパ芽細胞株やバーキットリンパ腫細胞の成長を選択的に遅延させたことである。このように、H31によるEBNA1依存性のDNA結合の阻害は、EBV感染細胞におけるEBNA1依存性の転写およびEBVエピソームの持続を減少させた。スクリーニング・アプローチにより、EBNA1-DNA結合、EBNA1-oriPトランザクティヴ化、EBNA1リンク領域を阻害する分子も同定された。また、EBNA1の二量体インターフェースからのトランケートされたペプチドに基づく阻害剤も報告されており、EBV関連がんの治療に対するEBNA1の「ドラッガビリティ」が確認されている[78]。
リガンド分子を収容する可能性のあるEBNA1結合ポケット(すなわち、「ドラッグ可能な」結合部位)の計算機による同定および構造的特徴は、以前に試験管内試験でEBNA1の阻害についてテストされた一連の化合物(PubChem AID-2381)に対するドッキングによって検証された[79]。ポケットの薬効評価は,薬効のある結合部位に属する多数のヒット化合物について,分子力学による一般化ボルン表面積計算による結合親和性予測と組み合わせて,誘導適合ドッキングおよび分子力学シミュレーションによって行われた。これらの研究により、EBNA1が創薬のターゲットとして確立され、AID-2381のアクティブなヒット化合物が個々の部位で相互作用するとEBNA1とDNAの結合を阻害するという計算上の証拠が得られた。Cullinan Oncology社は、Wistar研究所が発見した新規のEBNA1阻害剤であるVK-2019(EBNA1に結合し、EBNA1のDNA結合活性を阻害する)を開発している。現在、VK-2019の安全性と忍容性、薬物動態、薬力学および予備的有効性を評価するために、第1-2a相臨床試験(https://clinicaltrials.gov/ct2/show/NCT03682055)非盲検、用量増量および拡大、ヒト初回臨床試験が実施されている。
6. 細胞ターゲット
ウイルスの複製サイクルのステップを標的とした直接作用型抗ウイルス剤に代わる戦略として、ウイルスの複製に不可欠な細胞内タンパク質が、ウイルスの複製を特異的に阻害する新たな標的となる可能性がある。古典的な抗ウイルス剤は、少数のウイルスにしか作用せず、耐性発現がその特異性の特徴と考えられている。一方、ウイルスの複製に不可欠な細胞内タンパク質を標的とした抗ウイルス剤は、関連性のない様々なウイルスの複製に同じ細胞内タンパク質が関与している可能性があるため、より広い範囲のウイルスに有効であると考えられる。また、細胞事象を標的とした抗ウイルス剤は、ウイルスタンパク質を標的とした抗ウイルス剤に比べて、薬剤耐性ウイルスの変異株の選択速度が低いと考えられる。また、従来の抗ウイルス剤に耐性のあるウイルスの変異株に対しても、抗ウイルス剤は有効であると考えられる。しかし、細胞内のタンパク質を標的とする場合、細胞毒性や副作用の増加が大きな問題となる。
細胞性トポイソメラーゼIおよびII(Topo IおよびII)は、γ-ヘルペスウイルスの溶解性DNA複製に必須であるため[80,81]、ある種のTopo IおよびII阻害剤は、EBV感染に対する潜在的な抗ウイルス剤として考えられるかもしれない[82]。Topo II阻害剤は2つのカテゴリーに分類される。トポイソメラーゼ-DNA中間体を標的とするトポII毒薬と、酵素のターンオーバーを乱すトポII触媒阻害薬である[83]。トポイソメラーゼII毒薬には,抗腫瘍剤として使用されているエトポシドやドキソルビシンがあり,KSHVの複製やビリオンの産生を阻害することが示されたが,予想通り,かなりの毒性を示した[83]。一方,ノボビオシン,メルバロン,ルタマリンなどのTopo II触媒阻害剤は,ヒトγヘルペスウイルスに対して最小限の毒性で抗ウイルス活性を示した[83]。特に,(+)ルタマリンは,試験したTopo II阻害剤の中で最も高い選択性(SI > 63)を示し,細胞増殖にはほとんど影響を与えずにEBVのDNA複製とウイルス産生を阻害することができた[82]。したがって、ルタマリンは、EBV感染に関連するヒトの疾患の治療に使用できる可能性のある安全な薬剤であると考えられる。
Verdinexorは、SINE(Selective Inhibitors of Nuclear Export)化合物として知られる新しいクラスの新規小分子に属する。これらの化合物は、核輸出タンパク質であるXPO1に共有結合してこれを阻害し、XPO1依存性のタンパク質を細胞核内に封じ込める。Verdinexorは,免疫不全者が罹患する日和見ウイルスに対して,さまざまなレベルの有効性を示した[84]。Verdinexorは,Akata細胞におけるEBV複製の阻害に有効であり,50%有効濃度(EC50)は50nM,選択性指数は7であった。 Verdinexorの有効性は,ウイルスタンパク質SM(溶解複製中のEBV mRNAの核-細胞質間輸送に関与するアダプタータンパク質)がXPO1を介した核輸送に依存していることで説明できる。ベルディネクサーは、核外輸送を阻害することで、EBVのmRNAが細胞質に移動して翻訳されるのを防ぐ。
宿主の細胞性プロテインキナーゼおよびそれらが制御する下流の経路がヘルペスウイルス感染に重要な役割を果たしているという豊富な証拠があるため、いくつかの細胞性プロテインキナーゼ阻害剤が抗ヘルペスウイルス効果について試験されている[85]。移植患者のHCMV疾患の減少における哺乳類ラパマイシン標的(mTOR)阻害剤の成功は、ヘルペスウイルス関連疾患の治療における細胞性プロテインキナーゼ阻害剤の可能性についての更なる研究を促すかもしれない[86,87,88]。最近では,エベロリムスがDNA合成を遅らせ,感染の拡大を抑制し,サイトメガロウイルス感染を緩和することが示された[89]。
7. 薬用植物
薬用植物から単離された多くの化合物は,EBVの溶解複製を阻害することが知られている。その中でも、細菌感染、炎症、高血圧など様々な病気の治療に一般的に使用されているAndrographis paniculata(センシンレン)は、EBV IE遺伝子の転写とEBVビリオンの産生を阻害することが示された[90]。A. paniculataに含まれるジテルペノイドandrographolideは,抗炎症作用,抗血栓作用,抗がん作用,抗免疫賦活作用を有することから重要である。さらに,アンドログラフォリドはEBVだけでなく,HIV[91,92],インフルエンザウイルス,SARS[93],HSV-1[92,94]などの他のウイルスに対しても抗ウイルス活性を示した。アンドログラフォリドがEBVのBRLF1およびBZLF1遺伝子の転写をどのように阻害するかは現在のところ不明であるが,アンドログラフォリドがEBVのIE遺伝子の転写を活性化するシグナル伝達経路を阻害することはもっともである[90]。
Polygonum cuspidatum(イタドリ)の根から得られた酢酸エチルサブフラクションF3とその主要成分(すなわちemodin)は,EBVの溶解サイクルを阻害することが示された[95]。P. cuspidatumは,動脈硬化の治療に加え,癌,喘息,高血圧,咳などの治療にも用いられる漢方薬である。F3とエモジンは、EBVのIEタンパク質であるRta(R transactivator、BZLF1遺伝子の産物)Zta(Z EB replication activator、BZLF1の産物)EA-D(Early Diffuse Protein)の発現を用量依存的に減少させたことから、EBV複製サイクルの初期段階を阻害することが示唆された[95]。F3とエモジンは,BRLF1とBZLF1のmRNAの発現も阻害し,その結果,ウイルスの溶解性タンパク質の発現とEBVのDNA複製に影響を与えた。これまでの研究で、エモジンがp38,MAPK、ERK、JNKシグナルの活性化を阻害し、AP-1やATF1で活性化されるプロモーターの活性化に影響を与えることが示されていることから[96,97,98]、F3およびエモジンのEBV溶解サイクルに対する阻害には、シグナル伝達経路の活性化に対するエモジンの阻害が関与している可能性が示唆された[95]。緑茶の主要カテキンである(-)-エピガロカテキン-3-ガレート(EGCG)によるEBVの自発的溶解感染の抑制には,MEK/ERK1/2およびPI3-K/Aktシグナルの活性化の抑制も関与しているようである[99]。(-)-緑茶は、フラバノールまたはカテキンとして知られるポリフェノール化合物を多量に含むことが特徴であり、EGCGは、EBVの溶解サイクルの阻害など、緑茶の生物学的効果をもたらす主要な有効成分であると考えられている[100]。
最近、Saururus chinensis(ハンゲショウ)の根から単離されたリグナンの抗EBV溶解複製活性が報告された[101]。リグナンは,S. chinensis(チョウセンゴミシ)の主な有効成分であり,NF-kB [102]やHIVプロテアーゼ [103]の阻害活性や心血管作用など,幅広い生物活性を示すことが知られている[104]。S. chinensisのエタノール抽出物を分画して得られた19種の新規リグナンと9種の既知のリグナンのうち,Manassantin Bは,EC50が1.72 µMで,低毒性(CC50 > 200 µM),SI > 116という最も有望な阻害作用を示した。
Moronic acid(モロン酸)はトリテルペノイドのケト酸で,Rhus chinensis(ヌルデ)のガルスやブラジルのプロポリスに含まれている。モロン酸は,試験管内試験ではHIV [105],EBV [106],マウスではHSV-1に対して活性を示すことが明らかになっている[107]。Changらは,モロン酸がRta,ZtaおよびEA-Dの発現を阻害することを報告した[106]。さらに,モロン酸は,RtaがRta-response elementを含むプロモーターを活性化する能力を阻害したことから,モロン酸がRtaの機能を阻害していることが示された。一方、モロン酸はZtaのトランザクティベーション機能に影響を与えることがわかった。したがって、モロニック酸処理後にZtaとEA-Dが発現しないのは、Rtaのトランザクティベーション機能が阻害された結果、溶解サイクル中に生成されるEBV粒子の数が大幅に減少したためであると考えられる。一方,Thelypteris torresianaに含まれるフラボノイドの1つであるプロトアピゲノンは,ウイルスの溶解サイクルを妨げるZtaのトランザクティベーション機能を阻害し,Rtaプロモーターの機能には影響を及ぼさないことが報告されている[108]。
多様な天然物から単離された116化合物の実験室コレクションをスクリーニングした結果,アンジェリシンは,MHV-68,2つのヒトγヘルペスウイルスであるEBVおよびKSHVに対して抗ウイルス活性を示した[109]。アンジェリシンは,Psoralea corylifolia(オランダビユ)の種子やAngelica archangelica(セイヨウトウキ)の根に含まれるフロクマリンで,プソラレン(長波長の紫外線照射とともに様々な皮膚疾患の治療に用いられる光合成物質)の一種である。γ-ヘルペスウイルスに対するアンゲリシンの作用機序に関する詳細な分子メカニズムは不明であるが,アンゲリシンはRTAプロモーターの自動活性化を阻害し,その結果,複製性溶解サイクルの初期段階を阻害することが明らかになっている[109]。また,フロクマリンは,インフルエンザウイルス[110]やレトロウイルス[111]に対しても有効であることが証明されている。
8. 溶解導入療法における抗ウイルス剤の使用
溶解性複製が腫瘍細胞の死を促進することから、潜伏感染した腫瘍細胞でウイルスの溶解性複製を再活性化する戦略(溶解性誘導療法)の開発が注目されている。さらに、ウイルスを保有する腫瘍細胞は、溶解サイクル中に発現するウイルスキナーゼによって活性化される抗ウイルス剤(例えば、ガンシクロビル)によっても死滅する可能性がある。抗ウイルス剤と溶解サイクルの誘導剤の組み合わせは、EBVやKSHVが原因の腫瘍を治療するための有望な戦略として検討されている[112,113,114,115]。外部からの刺激による溶解活性化に特に抵抗を示す細胞株があり、また、溶解誘導刺激に反応する細胞株においても、溶解サイクルの活性化に反応しない細胞の亜集団が残っているため、溶解サイクルへの誘導効率を向上させるためのさらなる研究が必要である[112,116]。
9. 結論と今後の展望
抗EBV療法は、特に免疫力が低下している患者にとって、依然として大きなアンメットメディカルニーズである。他のヘルペスウイルスに承認されている抗ウイルス剤のうち、EBV関連疾患に対して評価されたものは、残念な結果となっている。いくつかの抗EBV薬の候補があるが、その有効性を示すには多くの課題が残されている。EBV関連疾患の治療戦略や、EBV疾患に関連した合併症を避けるために免疫抑制患者の予防に使用できる分子を開発するためには、さらなる研究が必要である。EBVワクチンは、EBV初感染による大きな負担を軽減し、ある種のヒトの悪性腫瘍の発生率を低下させるために大きな利益をもたらすはずであるが、EBVワクチンの開発は非常に遅れている。
EBVの増殖期と潜伏期の両方の感染に対処できる可能性のある新しい戦略は、CRISPR/Cas9ゲノム編集技術を用いて、ウイルスの適合性に必要なウイルスの遺伝的要素を標的にすることである。Lebbinkのグループは、複数のガイド付きRNA(gRNA)でEBVのゲノムを同時に標的にすることで、潜伏感染したEBV形質転換細胞からウイルスをほぼ完全に除去できることを実証した。このことは,新しいゲノム工学技術を用いて,病原性ヒトヘルペスウイルスを管理する治療法を開発するための新たな道を開くものである[117]。