
Contents
pubmed.ncbi.nlm.nih.gov/32095358
Ecosystem antifragility: beyond integrity and resilience
2020年2月11日オンライン公開
要旨
生態系の回復力という概念を、情報理論的なアプローチから生態系の完全性との関連において検討する。生態系の回復力を高めるための生態系の特性、生態系の摂動に対する応答、複雑性という3つの主要な物語を特定し、このテーマに関する文献を要約する。また、理論的・定量的な発展や応用例など、独創的なアイデアも紹介する。主な貢献は、レジリエンスを再考する新しい方法であり、数学的に正式で、実世界のアプリケーションで発見的に評価することが容易な、生態系の反脆弱性である。生態系が環境の変動性から利益を得ている場合、生態系は反脆弱である。なぜなら、弾力的で堅牢なシステムは単に摂動に強いだけであるのに対し、反脆弱な構造はストレスに耐えるだけでなく、ストレスから恩恵を受けるからだ。
キーワード
反脆弱性、生態系の完全性、レジリエンス、複雑性
はじめに
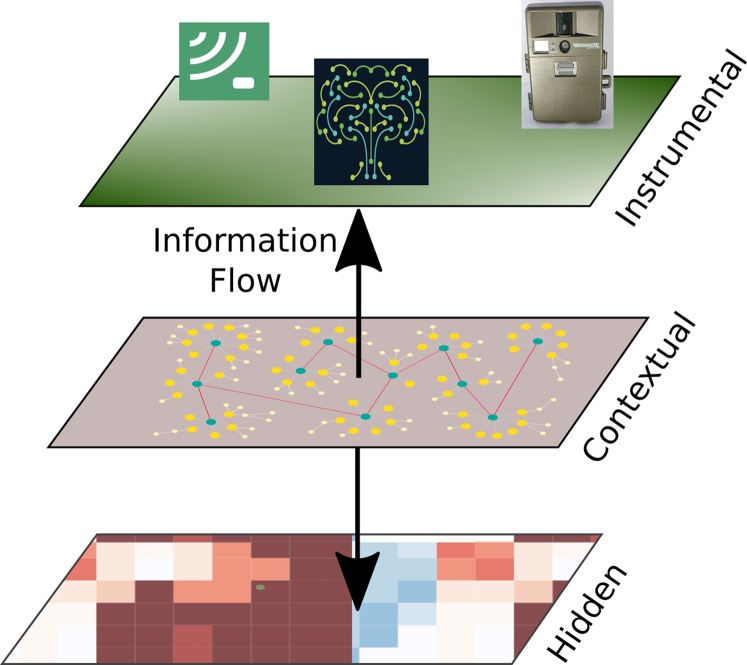
持続可能な開発には、自然生態系の構造と機能、すなわち完全性を維持することが必要であることは言うまでもない。以前の研究(Equihua et al., 2014)では、生態系の完全性を定量化するための運用フレームワーク(図1参照)と、生態系の介入方法を管理し、持続可能な道に沿った開発を促進するのに役立つ実行可能な基準が開発されている。

図1 エコシステムインテグリティ3層モデル。
生態系の完全性とは、生態系の構造的特徴、発生プロセス、および後天的構成において、特定の発現をもたらす生態系の構成における根本的な属性であると理解される。つまり、生態系の健全性は、熱力学的メカニズムに由来する自己組織化のプロセスから生じるものであり、局所的に存在する生物相やその配置にあるエネルギーや物質を通じて、「最適」な運用ポイントに到達するまでの間、固定的ではなく、むしろ物理条件の変動や生物相や環境に生じる変化に応じて変化する。生態系の健全性の3層モデル(3TEI)を示す。内部の層は観察者には見えないが、その状態は、構造(組成やその他の生物多様性の特徴を含む)と機能に関する測定値が得られる機器層または観察層で得られる情報によって推測することができ、もちろん生態系が発展する背景を考慮している。矢印の先端は、想定されるメカニズム的な影響の方向を示しているが、情報はどちらへも向かうことができる。
人類は、生物圏のダイナミクスを変化させる圧倒的な強制因子であると認識され始めている。この見解では、地球システムは、採用する概念的スタンスによって、人新世(Steffen, Crutzen & McNeill, 2007)、技術新世(López-Corona, Ramrez-Carrillo & Magallanes-Guijón, 2019)、あるいは資本新世(Haraway, 2015)と呼ぶことができる地学時代に入ったと解釈することができる。人間の意思決定がこのような推進力を持つため、生態系が人間の摂動から回復する(またはしない)方法を理解し、測定することに長い間関心が持たれてきた。持続可能性を運用するためには、生態系の完全性(自然資本資産の量を表す状態変数の構成要素:状態×拡張)に加えて、この生態系の能力の定義とそれを評価する指標が必要である。回復特性は、すでに生態系レジリエンスの概念に包含されている。ここでは、次の小節で述べるように、情報理論が生態系の完全性と回復力の両方を包含するのに適したフレームワークであることを提案する。
Equihuaら(2014)によると、多くの研究が生態系の完全性の適切かつ包括的な定義を見つけることを目的としているが、現在までに一般的な合意は得られていない。Equihuaと共同研究者は、複雑系アプローチを採用しており、生態系は、その構造(生物学的組成または生物多様性を含む)に制約を受けた自己組織化実体であり、熱力学的散逸系特性(Kay, 1991;Regier, 1995;Manuel-Navarrete, Kay & Dolderman, 2007;Michaelian, 2005)および進化過程(Levin, 2005)によって機能していると考えられている。
生態系レジリエンスの概念は、Holling (1973)によって初めて導入され、自然または人為的な誘因による環境ストレスの存在下で、自然構造が持続することを表現している。Hollingは、その代表的な著作において、システムダイナミクス分析に従っている。そこでは、レジリエンスを次のように定義している:「システムが擾乱を吸収し、変化を受けながら再編成し、本質的に同じ機能、構造、アイデンティティ、フィードバックを維持する能力」(Walker et al., 2004)と定義されている。しかし、一般に同義語として用いられるレジリエンスやスタビリティには、少なくとも163の異なる定義がある(少なくとも70の安定性/レジリエンスの概念に分類される)(Grimm & Calabrese, 2011)。例えば、Saint-Béatと共同研究者(Saint-Béat et al., 2015)は、レジリエンスと、しばしば互換的に用いられるその他の関連概念を以下のように要約している:
- レジリエンスとは、システムが攪乱後に平衡状態に戻る速度のことである(DeAngelis, 1980;Pimm & Pimm, 1991)。復帰時間が長いということは、レジリエンスが低いということと同じだ。コミュニティのレジリエンスは、最もレジリエンスの低い種(平衡状態への復帰が最も遅い種)に依存する。このレジリエンスの定義は、Holling (1996)が定義した「エンジニアリング・レジリエンス」に相当し、バランスあるいは安定した状態は一つしかないと仮定している(Gunderson, 2000)。
- 持続性とは、ある変数が異なる状態に変化する前に、同じ状態を維持する時間のことである(Pimm & Pimm, 1991)。持続性は、システムが時間と共にそれ自身を維持する能力の尺度である(Loreau et al., 2002)。
- 抵抗力とは、外的擾乱の存在下で、生態系がその初期状態を維持する能力として説明される。(Harrison, 1979)。生態系内の(量と強度の)小さな変化だけが、高い抵抗力に対応する。この概念は、Holling(1996)が定義した「生態系レジリエンス」に類似しており、様々な安定状態が存在することを示唆している。
- ロバストネスは、環境の安定性の耐久性に関係する。そして、ロバスト性は、生態系が異なる状態に変化する前に、どの程度の撹乱に耐えることができるかを示す尺度である。(Loreau et al., 2002)。食物網が堅牢であればあるほど、より安定していると言える。
統一的なフレームワークとしての情報理論
生態系の完全性は、熱力学的メカニズムに由来する自己組織化のプロセスから生じる。このプロセスは、その地域に存在する生物相と、彼らが自由に使えるエネルギーや物質を通じて、最適な運用ポイントに到達するまで続くが、このポイントは固定されておらず、むしろ物理条件の変動や生物相に生じる変化に応じて変化する(Equihua et al, 2014).
この研究では、生態系の完全性と摂動に対する応答との相互作用を強調したい。生態系が摂動にどのように応答するかを理解するための標準的な方法は、生態系の回復力、すなわち生態系が摂動に耐えて目標状態に戻ることができる大きさであり、別の安定状態にシフトするのとは対照的である。
両者は関連する概念だが、同じではない。外部からの摂動が生態系の完全性の状態に影響を与えることは明らかであり、同様に完全性の状態は、摂動に対処するための生態系の能力を反映する。しかし、例えば、枯渇などの劣化プロセスを経た生態系であっても、構造や組成の点では高い完全性を示しているが、機能群に1種しか含まれていない場合がある。この場合、機能はまだ代表的であるため、生態系は完全性を維持しているが、その重要な種に影響を与える摂動が発生すると、非常にもろくなることが予想される。このように、完全性とレジリエンスは同じではない。前者は生態系の静的なイメージであり、後者は生態系のダイナミクスに大きく関係する。
ウラノヴィッチは2003年の論文で、ニュートンの法則で完全に記述される古典力学やマクスウェルの方程式による電磁気学とは異なり、エコロジーは登場から100年以上経った今日でも、あまりにも多様で対立しており、一つのまとまった理論にまとまることはできないと指摘した(Ulanowicz. 2003)。
しかし、情報理論に基づく指標は、自然と人間のシステム要素を橋渡しし、システムの異質な状態変数を理解することができると提案されている(Cabezas et al., 2005)。さらに、情報理論は、生物システムに関する私たちの知識において主要な役割を果たしており、生態系を情報システムそのものとみなすことは、新しいことではない(Nielsen, 2000;Straskraba, 1995)。例えば、Brooks & Wiley (1984),Wicken (1987),Michaelian (2012)などの複数の著者が、進化過程におけるエントロピー、熱力学、情報の重要性を指摘している。
同様に、Fath, Cabezas & Pawlowski (2003)が説明するように、生態系は発展するにつれて、平衡状態から遠ざかっていく。その結果、遺伝情報の容量が大きい生物が生まれるという仮説が提起されている(Jørgensen & Nielsen, 1998)。したがって、このような情報の利用は、個々の生物の内部構成と生態系全体の構造的な組織とを結びつけることになる。さらに、すべての生命システムは、環境から情報を取得し、環境の変化に対応するためのメカニズムを生み出す必要がある。そのため、生物システムは、センサーから得られるデータをもとに、常に内部表現で環境をモデル化している。この文脈では、ハードウェア(センサー)の性能が高ければ高いほど有利だが、関連する情報を抽出、要約、統合するこれらの表現がうまく構築されているかどうかが、決定的な競争優位性をもたらし、最終的に生存と絶滅を分けることができる。実際、この機能は非常に重要であり、Hidalgoら(2014)は、生態系はこの計算能力が最大になる臨界状態にある傾向があると提案している。
以上のことから、生態系の完全性と生態系が摂動にどのように応答するかは、生態系の状態を評価するために必要な2つの別の次元であり、情報理論はその両方を理解するための一般的な統一フレームワークを提供できると考えている。
このことを念頭に置いて、私たちは「生態系の完全性」「生態系の回復力」「情報理論」をキーワードとしたレビューを実現した。最初の2つは生態系ダイナミクスの異なる次元であり、非常に連関しているが些細なことではなく、情報理論はそれぞれの個別性と相互作用を理解するのにふさわしい一般的枠組みである。このレビューの結果は補足資料にまとめられているが、ここでは、発見された主なナラティブについて議論することに重点を置き、最も重要なことは、生態系の回復力に代わる新しい概念の提案である。この概念は、より一般的で、情報理論の枠組みによってよりよくサポートされているからだ。
文学の語り
情報理論のレンズの下でのレジリエンスと生態系の完全性との関係において、ナラティブは、生態系がよりレジリエントであり、したがって完全性を維持することを可能にする生態系の特徴を(情報理論のツールを用いて)特定しようとすることから、新しい数学的概念に加えて、時系列やネットワーク分析を用いたより技術的なアプローチに移行したと考える(図2参照)。相互作用の重要性と複雑系アプローチの重要性が強調されている。最後に、この分野は、生態学に再び戻り、生態系のレジリエンスの特徴や特性を洗練させるという輪を完成させる。

図2 選択した論文におけるコンセプトとナラティブの要約。
よりレジリエンスを高めることができる生態系の特性
「はじめに」で述べたように、生態系の内部特性がその完全性と回復力の両方を決定する可能性があることは直感的なことである。この小節では、この考え方が主な文献でどのように表現されているかを要約し、両概念と情報理論とのつながりを明確にすることを試みる。
まず、Aronson & Le Floc’h (1996)による改訂論文を考えてみよう。そこでは、著者らは、Noble & Slatyer (1980)の概念を修正した以前の研究(Aronson et al., 1993)に言及しており、その中で、再発する外乱に対する種の反応を決定する必須特性のいくつかのカテゴリーを説明している。
Aronson論文では、生態系完全性モデル(3TEI)(Equihua et al., 2014)で使用される属性と同じ種類の、生態系の構造と機能に関連し、それを表す特徴や属性を捉える試みとして、表1にまとめたVEAs(Vital ecosystem attributes)を定義している。生態系の回復力の観点から、これらのVEAは、生態系がより回復力のあるときに最適な値になると解釈することができる。興味深い点は、3TEIモデルとは異なり、VEAsは大規模なフィールドワーク(多くの場合、コストがかかり、拡張性がない)を必要とするため、VEAsは生態系の状態の傾向を全国的に評価するための良い選択肢ではない可能性が高いことである。
表1 Aronson et al. (1993)による生態系の重要な属性
| 構造 | 機能 |
|---|---|
| 多年草の種の豊富さ | バイオマス生産性 |
| 年間種数豊かさ | 土壌有機物 |
| 植物の総面積 | 最大利用可能水量 |
| 地上部ファイトマス | レインオフ効力係数 |
| βの多様性 | 雨天時の使用効果 |
| 生命体スペクトル | 水利用可能期間の長さ |
| キーストーン種 | 窒素の利用効率 |
| 微生物バイオマス | 微小共生生物の有効性 |
| 土壌生物相の多様性 | 周期的なインデックス |
AronsonとLe Floc’h (1996)は、VEAsの考えから、景観スケールで使用するための16の定量化可能な主要特性を特定し、景観の観点から生態系の修復やリハビリの結果を評価するための新しいVLAs (vital landscape attributes)を使用している。最も興味深いのは、VEAが生態系の完全性に関連するように、VLA(表2参照)が、私たちがすでに取り組んでいる景観の完全性(3TEI)モデルに関連する可能性があるということである。
表2 Aronson & Le Floc’h (1996)が提唱するVital landscape attributes
| 重要な景観属性(VLAs) |
|---|
| 地形の種類、数、範囲 |
| 生態系の数 |
| 土地ユニットの種類、数、範囲 |
| かつての人間の用途の多様性、長さ、強度 |
| 現在の人間の用途の多様性 |
| 土地利用タイプの数および比率 |
| エコチューンズゾーンの数・種類 |
| コリドーの数・種類 |
| 選択された重要な生物群(機能群)の多様性 |
| エコチューンを定期的に横断する生物の範囲と様式 |
| 生態系内外の水、栄養、エネルギーの流れや交換を表す循環指標 |
| 水と栄養の移動のパターンとテンポ |
| 景観の人為的変容のレベル |
| 外乱の広がり |
| 生物学的侵略の件数と重要性 |
| 合法・違法を問わず、さまざまな劣化の原因の性質と強さ |
これとは異なる考え方として、Gustavson, Lonergan & Ruitenbeek (2002)とその共同研究者による総説では、情報理論の観点から生態系の回復力の代理として機能する一般的な指標を開発している。著者らは、レジリエンスを記述し評価する試みがなされてきたが、レジリエンスの特性と生態系ダイナミクスの間の全体的な予測や理論的な関連性はまだ確立されていないと報告している。同様に、安定性と構造の間に起こりうる相互作用についても、多くのことが議論されてきた。一般に、レジリエンスの特性と生態系の仕組みとの間の予測可能な相互作用は、直感的には明らかではなく、存在しないかもしれない。
この目的のために、彼らはUlanowiczの上昇理論(Ulanowicz, 1986)に注目した。この理論は、生態系のネットワークの枠組みを通る情報の流れの大きさの尺度である。この理論を使うには、すべての種の相互作用の性質と大きさを比較的完全に記述する必要があるという制約がある。
ascendancyは、コンポーネントaとbの間の平均相互情報(Ulanowicz, 1986)で定義される、
【原文参照】
As=K∑i∑jp(ai,bj)ログ[p⟨bj|ai⟩p⟨bj⟩] (1)
p⟨bj|a i⟩aiが発生した場合のbjの確率、p(bj)はbjが発生する確率である。
上昇力の上限は成長能力であり、能力と上昇力の区別は俯瞰系と呼ばれ、経路の多重性を表すため、最終的には生態系の複雑性と結びつくことができる(Zorach & Ulanowicz, 2003).
複雑な構造は、生態系の健全性と回復力にとって重要である。なぜなら、後で定義するように、複雑さに関連するプロセスの多様性(すなわち、シャノン情報で測定)は、システムの生存に重要な役割を果たすからである(Ulanowicz et al., 2009)。特に、生態系の長期的な健全性と回復力を高めるためには、特定の高密度なネットワーク構造が有利である。このような仕組みは、十分に効果的であり、十分に多様である。
最終的に、「上昇力」は、生態系がその組織と規模の組み合わせによって混乱に打ち勝つ能力を一つの指標として捉え、持続可能性を達成するためには、生態系が妨害される前の「上昇力」のレベルに戻ることができることが望ましいとされた(Reynolds, 2002).
この時点で、いくつかの点を明らかにしておくと便利である。生態系には固有のアトラクターがあるわけではなく、さまざまな開始条件に対して生態系が進化する傾向がある状態のサブセットである。便利な比喩として、卵の容器にボールを入れ、その中にアトラクターを入れるというものがある。ボールは、ある時点まで同じ流域に戻り、そこから新しい流域に落ちていく。これまで述べてきたように、システムが特定のアトラクターに戻る能力は、レジリエンス(回復力)であり、生態系の完全性と関連している。完全性のレベルが高ければ、レジリエンスも高いということは予想されることである。しかし、一方では、この関係はおそらく非直線的であり、生態系はレジリエンスに反映される前に重要な完全性の喪失を経験する可能性があることを意味する。一方、生態系は比較的高いレベルの完全性を持っているかもしれないが、例えば、特定の機能グループに固有の種が存在し、摂動によってこの種が排除されると、システムは一挙に回復力を失うかもしれない。レジリエンスの弱い概念は、いわゆる工学的レジリエンスと呼ばれるもので、システムが特定のアトラクターに戻るのに必要な時間で測定されるものである。この作品では、レジリエンスについて語るとき、単に工学的なレジリエンスではなく、生態学的なレジリエンスを意味する。
第3回目の論文改訂で、Saint-Béatら(2015)の主な目的は、組成、ダイナミクス、最終的には生態系の持続性、強度、または回復力を評価することができるという観点から、明確な生態系が地球変動にどのように反応するかを知ることである。
著者らは、生態系ネットワーク評価(ENA)が、定量的・定性的な要素を組み合わせて局所的な安定性を記述するための有効なアプローチであることを示した。しかし、局所的な安定性と大域的な安定性を組み合わせた実際の事例を記述することは、依然として不完全な作業であることを警告している。
そして、ENA から得られたレジリエンスに関連する3 つの結果、すなわち、(a)生態系の構造と機能における種の多様性の役割、(b)栄養リンクの数と相互作用の強さ、(c)循環能力、雑食性の広がり、上昇志向の観点から見た生態系の安定性に焦点をあてている。
高い生物多様性(外来種を考慮しない)は、環境撹乱に反応して生態系が大きく変化する脅威を最小化することに寄与すると提唱された(McNaughton, 1977)。草地区画における種の侵入に関する実験では、地域の生物多様性が様々な侵入者の定着と成功を減少させることが示されている(Kennedy et al., 2002)。同様に、草地の多様性を操作した研究によると、多様性が高まると種間競争が改善され、侵略の危険性が低下することが示されている(Naeem et al., 2000;Hector et al., 2001)。同様に、種の多様性も沿岸生態系の回復力を高めることに貢献することが示されている(Worm et al., 2006)。同様に、種の多様性を維持することは、生態系の完全性を維持し、人間社会にとって重要なサービスを提供するために極めて重要であると考えられる(Chapin Iii et al., 2000)。
しかし、著者らは、生態系は複雑であるため、多様性のような単一の変数を考慮するだけでは、生態系の安定性を十分に評価することはできないと警告している。したがって、食物網の構造と生態系の機能におけるその役割を十分に理解するためには、ENAの枠組みで全体的な指標を構築することがより良いアプローチであることを示唆している。
もし、十分に詳細な栄養ネットワークを構築することができれば(一般的には非常に難しいが)、標準的なネットワーク分析ツールを使って、例えば、ネットワークのトポロジー(すなわち、コネクタンス)を理解することができるだろう。その意味で、著者らは文献から得た証拠を要約し、リンクの増加は種の分布の変動による影響を分散させ、安定性を高めることを示した(MacArthur, 1955)。したがって、連結性が高まれば、生態系の回復力も向上する(DeAngelis, 1980)。したがって、連結性は食物網の頑健性、ひいては生態系の安定性を測る尺度として有用であると思われる。
接続性に加えて、相互作用の強さの多様性の重要性を示している。Ulanowicz (1983)の考えに従い、彼らは生態系の安定には弱い相互作用と強い相互作用がバランスよく存在することが必要だと主張する。さらに、捕食者と被食者の主要な相互作用が、高い多様性の中で弱い相互作用と組み合わされる場合にのみ、食物網は安定する。このように、生態系の中で変動し適応する能力があるため、弱い相互作用の種は食物網、ひいては生態系を安定させる力として機能する。
弱い相互作用の種は、生態系内で他の種との関係が緩やかであり、密接に依存していない種を指す。これらの種は、生態系内で様々な役割を果たすことができ、特定の関係に縛られることなく柔軟に適応する。以下に、弱い相互作用の種のいくつかの例を挙げる。
- 一般主義者(汎食性動物): これらの種は、多くの異なる種類の餌を摂取できるため、食物網内での相互作用が比較的弱いである。彼らは、多様な食物源に適応する能力があり、特定の餌に依存しないため、生態系の安定性に寄与する。
- オムニバランスの捕食者: これらの捕食者は、複数の被食者種との相互作用を持ち、それぞれの被食者に対する依存度が低いため、弱い相互作用を持っている。そのため、被食者の個体数の変動に柔軟に対応でき、食物網の安定性に貢献する。
- 競争関係の弱い種: これらの種は、資源の利用方法や生態的ニッチが異なるため、競争関係が弱く、生態系の安定性に寄与する。例えば、異なる植物が異なる土壌タイプや光環境を利用することで、競争を避けて共存することができる。
- 相互作用の弱い共生関係: いくつかの共生関係では、相互作用が比較的弱く、互いに独立した生活ができる場合がある。例えば、植物と菌類の間の共生関係では、両者が互いに利益を得ることができるが、どちらかが欠けても生存できることがある。このような関係は、生態系内での適応力を高め、安定性に寄与する。
これらの例からわかるように、弱い相互作用の種は、生態系内での柔軟性と適応力を高め、生態系の安定性に寄与する。弱い相互作用の種は、他の種との関係に縛られず、変化に対して適応できるため、生態系全体の回復力を向上させる。これは、生態系が外部のストレスや環境変化に対処できるようにすることが重要であるため、特に重要である。環境の変化が激しい現代社会において、弱い相互作用の種を含む多様で健全な生態系の維持は、生物多様性の保全や環境の持続可能性にとって重要な課題となっている。(by GPT-4)
安定性をより深く理解するために、Saint-Béatら(2015)は、サイクリング、雑食の存在、上昇の効果を検証している。
例えば、雑食性の存在は、生態系に栄養の柔軟性を与え、生態系の健全性と回復力を反映する明確な有益な特徴である。研究者らは、雑食性種が環境撹乱に対処するための優れた緩衝材となり、撹乱後の栄養経路を迅速に移動させることで生態系の迅速な反応を可能にすると主張する。例えば、攪乱が低栄養レベルに影響を与えた場合、それに直結する雑食性の種は迅速に反応する。それに比べ、特定の捕食者は、攪乱が自分のレベルに達するまで待たなければならないので、反応時間が長くなる。
Gustavson, Lonergan & Ruitenbeek(2002)の総説と同様に、Saint-Béatら(2015)は、生態系の機能を評価するための重要な指標として「上昇性」を用いることができることを示している。著者らは、「上昇性」の機能を理解するためには、2種類の安定性を区別する必要があることを示す。内的安定性が高いシステムとは、強く組織化された構造を可能にする十分な内的制約を持つシステムであり、高い上昇性(高い相互情報量)に相当する。一般的にこの条件下では、生態系は内部の摂動に対しては何らかの形で保護されているが、外部の摂動に対しては脆弱なままである。一方、上昇性が低いと冗長性につながるため、生態系は外的擾乱に対してより強くなる。興味深いことに、上昇性が高すぎると、ストレスの特徴として認識され、生態系の回復力が低下する可能性がある。
まとめると、臨界の枠組みの下での生態系の動的応答(Ramírez-Carrillo et al., 2018)において、健全な生態系とは、頑健性と適応性のバランスが取れたところにあると考えられる。ネットワークトポロジーの場合、生態系が安定するためには、強い相互作用と弱い相互作用のバランスがよく発達する必要がある。出世物語の場合、安定した生態系は出世と俯瞰のバランスがよく発達している必要があり、生態系に抵抗力と回復力を与えているようだ。
このことから、これら3種類のバランスはすべて、生命システムのより一般的な進化戦略である「反脆弱性」の特殊なケースである可能性があると考えられる。
生態系の撹乱に対する応答
Cabezas et al. (2005)は、生態系の完全性や回復力そのものではなく、持続可能性に主眼を置いた論文を発表している。しかし、彼らは、人間の構成要素(文化、経済、法律など)の構造と運用が、自然の構成要素の構造と運用(生態系の栄養結合、生物多様性、生物地球化学サイクルなど)の持続性を高めるものでなければならず、その逆もまた然りであることを強調している。「持続性」と「自然の構成要素の運用」という考え方にこそ、その関連性がある。彼らの観点では、持続可能性(Ramírez-Carrillo et al., 2018)を達成するためのシン・クア・ノン条件は、フィッシャー情報(Cabezas & Fath, 2002a)を用いて概念化した生態系の安定であり、その見解はいくつかの論文(Eason & Cabezas, 2012,Karunanithi et al. 2008;Mayer et al., 2007,Mayer, Pawlowski & Cabezas, 2006;Zellner et al., 2008;Ahmad et al., 2016;Karunanithi et al., 2011;Gonzalez-Mejia et al., 2012)、主にFisher情報に基づく情報理論アプローチ(Mayer、Pawlowski & Cabezas, 2006)により、いくつかの論文でさらに発展させている。
フィッシャー情報とは、統計学と情報理論の分野で用いられる概念で、未知のパラメータを推定する際の情報量を表す指標である。具体的には、観測データをもとに未知のパラメータをどれだけ正確に推定できるかを示すもので、フィッシャー情報が高いほど、推定の精度が高くなる。
フィッシャー情報は、サンプルデータのばらつき(分散)と密接に関係しており、データのばらつきが小さいほどフィッシャー情報は高くなる。逆に、データのばらつきが大きいと、フィッシャー情報は低くなる。
レジリエンス(回復力)とフィッシャー情報の関係については、システムが機能や構造を根本的に変化させる前に耐えることができる摂動(外部からの影響)の強度、頻度、期間によって定義されるとされている。レジリエンスの高いシステムでは、フィッシャー情報が同じ値かそれ以上に戻るという仮説が成り立つとされている(Cabezas et al, 2005)。これは、レジリエンスの高いシステムが、摂動による影響を受けた後でも、情報の精度を維持または回復できる能力を持っていることを示している。(by GPT-4)
安定性や回復力を理解するために、Binder (2000)やMayer et al (2007)に従って、ある状態変数の実際の値θを推定する中心問題を考えてみよう。この推定は、あるランダムノイズxの存在下での不完全な観測y= θ +xからの推論過程からもたらされる。この種のθの測定推論過程は、その結果が推定量θ ^焼け石に水θ^(y )の場合、平均二乗誤差がCramer-Raoの不等式に従う。
e2I≥1 (2)
ここで、Iはシステムのフィッシャー情報であり、次のように計算される。
I=∫dyP0(y|θ)[dP0(y|θ)dθ]2 (3)
ここで、P0(y|θ)は、問題の状態変数の真の値θが与えられた場合に、特定の値yを測定する確率密度関数である。
そして、情報が増加するにつれて誤差が減少するので、フィッシャー情報は、測定-推論プロセスからの推定θの品質として理解することができる。次に、システムが、スマート測定値yに関連する位相ベクトルs=(x1,…,xi,…,xm)を定義するm個の状態変数xiを有する位相空間によって特徴付けられると、以下のように証明することができる。
I(s)=1T∫0Ts”2s′4dt (4)
ここで、Tはシステムの1サイクルに必要な期間、s′(t)は接線速度、s″(t)は位相空間におけるシステムの経路に接するスカラー加速度である。両者は、状態変数xiの観点から次のように計算される。
s′(t)=∑im(dxidt)2———- (5)
s”(t)=1s′(t)∑im(dxidtd2xidt2) (6)
接線速度と加速度を計算するためのシンプルでロバストなアプローチとして、3点差分スキームを使用する。
dxidt=αxi(t+Δta)-(α2-1)xi(t)-xi(t-αΔta)α(α+1)Δta (7)
d2xidt=αxi(t+Δta)-(α+1)xi(t)-xi(t-αΔta)α(α+1)Δt2a/2 (8)
ここで、xi(t)は中心データ点、xi(t-Δta)は中心xiに続く次の点、xi(t-Δtp)はその前の点である。等間隔の点の場合、Δta=Δtp、α=Δtp/Δtaは前後の時間空間の比率である。
Frieden (2007)やCabezas & Fath (2002b)が示唆するテーゼは、フィッシャー情報のシフトが、ダイナミックなシステムにおけるレジームの変化を示唆するというものである:
- フィッシャー情報は、測定値のばらつきの関数である。ばらつきが小さいとフィッシャー情報が高くなり、フィッシャー情報が小さいとばらつきが大きくなる。
- 安定した体制にあるシステムは、フィッシャー情報が一定である傾向がある。その後、組織の損失により、変動が大きくなり、フィッシャー情報が減少する。
- 自己組織化システムは、そのばらつきを減らし、フィッシャー情報を得ることができる。
- 「レジリエンスが、システムが機能や構造を根本的に変化させる前に耐えることができる摂動の強度、頻度、期間によって定義されるとすれば、よりレジリエンスの高いシステムでは、フィッシャー情報は同じ値かそれ以上に戻るという仮説が成り立つ」(Cabezas et al, 2005).
この最後の点は、生態系の回復力を正式に情報的に定義しているだけでなく、フィッシャー情報によってそれを測定する具体的な方法を提供しているため、非常に興味深いものであることがわかった。このアイデアを検証するために、私たちは”US-Me1. “のNDVIデータを分析した:このサイトは 2002年のEyerly山火事で深刻な被害を受けた中間齢のポンデローサ松林として記録されているオレゴン州のAmerifluxサイト(Guy, Kosugi & Sulzman, 2007)である。約100の研究ステーションからなるAmeriFluxネットワークは、アメリカ大陸の大規模な陸域炭素循環合成のための主要な研究グループおよび情報供給者であり、微気象学、気象学、生物学の情報のデータベースを確立している。
NDVIのデータは、空間的、時間的、バンド/レイヤーのパラメータを使用して地理空間データセットをサブセットできるアプリケーション(AppEEARS:Extracting and Exploring Analysis Ready Samples)を使用してダウンロードした(https://lpdaacsvc.cr.usgs.gov/appeears/)。特に、1990年01月01日から2018年01月01日までのMOD13A3.006 1km2分解能のNDVIの月次データを使用した。
2002年の山火事に焦点を当てると、図3に示すように、山火事擾乱によって、システムはNDVIとそれに対応するフィッシャー情報の両方が大きく変化していることがわかる。フィッシャー情報は約18カ月後に以前の値に戻るが、NDVI値は戻らないことがわかった。

図3 赤はアメフラシのUS-Me1サイトの座標に対応する1km2ピクセルの正規化NDVI時系列(月次サンプリング)である。
青字は、Cabezas and collaboratorsアルゴリズム(https://github.com/csunlab/fisher-information)を用いたFisherの情報の対応値。
一方、フィッシャー情報の回復は、Filotas and co-workers(Filotas et al., 2014)が生態系レジリエンスを「生態系が元の機能と構造を維持する能力を失う前に吸収できる変化の量」と理解する方法と関連しているかもしれない。攪乱の後、著者らは、回復力のあるシステムは、その初期の構造、機能、フィードバック、言い換えれば、その完全性を回復する能力を持つと主張している。
一方、生態系の回復力を確保するためには、フィッシャー情報の基準は必要だが十分ではないようだ。例えば、Schmellerら(2018)が提案した、Aronsonのvital ecologicalattributes(Aronson & Le Floc’h, 1996)の現代版とも言えるような必須変数が、攪乱後に以前の値に戻ることを期待すべきである。Dutrieux (2016)は、キャノピーバイオマスの情報を提供する波長770~900nmの近赤外(NIR)に相当するTMとETM+のB4バンドと、キャノピー水分量の情報を提供する波長1550~1750nmの短波長赤外(SWIR1)に相当するB5バンドの信号を一つの指標に統合した:
NDMI=nir-sw ir1nir+sw ir1 (9)
裸地や薄い林冠の場合はNDMI値が低く、厚く完全に発達した林冠の場合は値が大きくなると予想される。(Wilson & Sader, 2002)。
筆者は以下のようなハーモニックモデルを作成し、外乱前後の値を比較した:
yt=αi+∑13γisin(2πjtf+δj)+εt (10)
ここで、ある時刻tにおける従属変数yは、切片αi、季節性を表す異なる周波数の調和成分の和、誤差εtの和として表される。このモデルでは、jは調和次数に対応し、1つは年周期、γjとδjはそれぞれ調和次数jの振幅と位相に対応し、fは時系列の既知の周波数(すなわち、年あたりの観測数)である。
その後、各スペクトルバンドと各時系列観測について、対応するマッチングモデルを用いて新しい値を推定し、以下の式でユークリッド距離を算出することができる:
Dt=∑i=1k(y^それが-yそれが)2———— (11)
そして、アマゾン流域の再生林から採取した3,596のLandsat時系列データのスペクトル回復時間に適用し、スペクトル特性の回復時間(スペクトル回復力)と呼ぶ推定値を作成した。その結果、攪乱された森林がスペクトル特性を回復するには、平均で約7.8年、大きなばらつき(SD = 5.3)があることがわかった。
今、私たちは、(a)状態(必須/重要)変数とそのフィッシャー情報の両方の初期値と最終値の間の距離、(b)これらの回復が起こるべき時間スケールの閾値を決定する方法という新しい問題を持っている。原理的には、(a)は生態学的完全性の測定から決定できると考えているが、現時点では未解決の研究課題であるため、ここでは扱わない。(b)については、この回復時間は現在の生態系の状態を示すものであり、グローバルな安定性の尺度としてではないこと、すなわち、別のアトラクターが存在する場合であることを忘れてはならない。
別の考え方では、Sidleと共同研究者(Sidle et al., 2013)は、生態系がどのような状況で回復力、ティッピングポイント、またはエピソード的なリセットを示すかを確認することに焦点を当てている。彼らは、生態系の回復力は生態学的な観点から生まれたが、最新の議論では、動的システムの特性が撹乱後に元の状態に戻らない可能性があることが認められている地球物理学的な文脈が取り入れられていると指摘している(Dakos et al., s18、Gough et al., h17、Langdon et al., n16、Ravindran. 2016、Moor. 2018、Stefen et al., n18)。ティッピングポイントとして知られるこの現象は、一般に、慢性的な(典型的には人為的な、しかし時には自然な)変化が生態系を閾値まで押し上げ、恒久的な形であってもプロセスや機能の崩壊を引き起こす場合に発生する。生態系のリセットは、エピソード的な自然災害が、ほとんどあるいは全く警告なしに閾値を破り、その結果、環境特性や生態系の機能が長期的に変化したときに起こる。特に興味深いのは、地球生物圏をシステム全体として捉え、現在の惑星危機の下で起こりうる軌道を研究したSteffenら(2018)の研究である。特に、彼らは、自己強化型フィードバックが、最終的に地球の生物圏システムを惑星の閾値に押し上げ、それを超えると、完新世の温度体制(パリ協定で定められた産業革命前の条件)付近での気候安定化を妨げる可能性があるリスクを探っている。最悪の場合、人類の排出量を減らしても、地球は「温室効果ガス」経路の継続的な温暖化軌道に追い込まれる可能性がある。
レビューした他の論文と同様に、Sidle et al.(2013)は、Equihua et al.(2014)で言及した「システムが回復力があると見なされる場合、それは一般的に特定の境界内に、おそらく最適な運用ポイントに近い状態で残っていると認識される」と述べている。そこで著者らは、「境界のある生態系」の下の変数はどれにすべきか、生態系をレジリエントとみなすための値の範囲をどう決めるか、という問題を改めて設定している。さらに言えば、生態系の摂動が起きてから、レジリエンス変数がその境界限界に戻るまでにどれくらいの時間をかけるべきか。原理的には、生態系の自然な特性時間スケールと同じオーダーであるべきだと考えている。しかし、繰り返しになるが、生態系の任意の状態変数に対する特性時間スケールの測定は、未解決の問題なのである。主な問題は、ほとんどの場合、問題の変数の力学的モデルを持たず、時系列のみであることである。Abe et al. (2005)では、Wigner関数を用いて、システムが最適な表現に到達する特別な時間スケールが存在するかどうかを探っている。観察された複数の時系列について、彼らは様々な異なる時間領域をカバーするエントロピー値を対比した。その結果、自然特性時間では、エントロピーが最小になる可能性が高く、時間-周波数空間における不確実性が最小であることを示唆することがわかった。別の選択肢として、システムの記憶がゼロになる傾向のあるτ0時間を考えることができるかもしれない。これは、C(τ)自己相関関数が横軸を横切るτ時間絶対値で定義される(Fossion et al.)しかし、完全な動的考察において、生態系の回復力にとって、時間を戻すことはそれほど重要ではないことをもう一度覚えておいてほしい。
複雑性の視点
Filotas and co-workers review article(Filotas et al., 2014)は、複雑性についての驚くべき紹介をしている。著者らは、複雑性を8つの特徴に分解し、それらを森林の新しい物語に関連付けることで、生態系の回復力と完全性との興味深い関連性を示している。一般的に、システムが複雑であるのは、十分な数の構成要素があり、十分な相互作用がある場合、または観察者の時間スケールに匹敵する速度で変化する場合、そしてほとんどの場合、その両方であると言われている。システムとしての森林と森林管理は、複雑さの勾配において高い位置を占めていることは確かである。

著者は森林に焦点を当てているが、明らかに、彼らが説明することはあらゆる種類の生態系に当てはまる。しかし、森林は広範囲かつ集中的に管理されており、また人間のシステムと深く結びついているため、良いモデルであると言えるだろう。このアプローチは、森林科学者や森林管理者が、森林を社会生態学的統合システムとして概念化し、複雑な適応システムとして森林を管理する方法の具体例を示すのに役立つだろう。
森林とは何かという定義には、少なくとも800以上の異なるものがある。その中には、同じ国で異なる目的やスケールで同時に使用されているものもある(Lund, 2006)。これは、緯度、気候パターン、土壌特性、人間との相互作用などの要因によって、森林の種類が大きく異なることが一因である。また、誰が森林を定義するかによっても異なる。経済学者が森林学者や農家に対して、それぞれの関心事に応じて、まったく異なる方法で森林を説明することができる。最も広く使われている定義のひとつはFAO(1998)によるもので、森林を「面積が0.5 ha以上、樹冠率が10%以上で、農業やその他の特定の非森林利用が主でない土地の一部」と定義している。若い森林や気候的要因で樹木の成長が抑制されている地域では、樹冠率の要件を満たしながら、その場で少なくとも5mの高さに達することができる樹木であることが望ましい。一般に、森林の定義は2つの異なる視点に基づいている。一つは、最小面積被覆、最小樹高、最小樹冠サイズなどの定量的な被覆/密度変数に関連するものである。もう一つは、植林地や農業活動、森林以外の樹木が森林内にあるかなど、その地域の特徴的な空間的特徴に関連するものである(Kleinn, 1991,1992,2001;Lund, 2006;Vidal et al, 2008)。問題は、天然林が著しく劣化したり、プランテーションに取って代わられたりすると、CO2の吸収など不可欠な生態系サービスが失われる可能性があることだ。しかし、多くの定義では、技術的には森林に分類され続けるかもしれない。
このように、生態系の基本的な重要特性である複雑性が、ほとんどの森林の定義には本質的に欠けている。著者らは、複雑系科学が、(1) 不均質性(2) 階層性(3) 自己組織化(4) 開放性(5) 適応(6) 記憶(ホメオスタシス?この8つの特徴は、構成要素の性質にかかわらず複雑系に共通するもので、記事ではそれを「森の言葉」で例示している。複雑系アプローチは、生態系の回復力と適応力を向上させるための理論と応用アプローチの両方にインスピレーションを与えたという結論は、私たちの記事に最も関連するものである。森林は複雑系の代表例であるが(Perry, Oren & Hart, 1994)、森林生態学と森林管理のアプローチは、この分野ではあまり言及されない複雑性を取り入れ始めたばかりである。
生態系の構成要素が空間的にどのように分布しているかという不均質性は、いくつかの観点から重要である。例えば、資源の空間的な分布は、動物の採食に制約を与える。様々な動物の採食パターンは、レビーウォークで記述されるように、多くの時空間スケールを含むことが広く証明されている(Viswanathan et al., 1999;Ramos-Fernández et al., 2004;Miramontes, Boyer & Bartumeus, 2012)。この統計的な振る舞いは、人間の運動パターンにも見られ(Brown, Liebovitch & Glendon, 2007)、複雑な環境における探索戦略の面で進化的に有利であると考えられている(Bartumeus, 2007)。さらに、資源の分布パターンは、種子散布に関連する採食行動を誘発する。これらのパターンは、生態系のダイナミクスにフィードバックされ、時間の経過とともに資源の分布に影響を与える(Boyer & López-Corona, 2009)。この論文では、生態系に対する人間の影響が、空間的不均質性を変化させることで生態系の複雑性を低下させる可能性があることを指摘している。例えば、森林の被覆や居住生物相は、森林内や森林間の栽培方法の強化や標準化によって均質化されてきた。これらのパターンを変えることは、効果的に処理された物質やエネルギーを維持し、地域の生物相を受け入れる景観の能力に大きく影響する。この変化は、ひいてはその完全性を低下させ、レジリエンスの喪失をもたらす。さらに、人新世(Steffen, Crutzen & McNeill, 2007)または技術新世(López-Corona, Ramrez-Carrillo & Magallanes-Guijón, 2019)の文脈において、極端な場合、人間の影響は森林異質性を変更し、歴史的に同等でない新しい生態系パターン(ニッチ構築)と相互作用を生み出す可能性がある(Seastt, Hobbs & Suding, 2008)。
侵略種によって空間的不均質性が変化すると、捕食(Doherty et al., 2015;Rayner et al., 2007)、競争(Harris & Macdonald, 2007)、疾病伝播(Wyatt et al., 2008)、さらなる侵略種の定着促進(Simbloff, 2011)などを介して生物多様性を脅かすことができる。侵略的捕食者による在来種の減少や絶滅は、生態系全体やそれ以上に広がるカスケード効果を生み出すことが報告されている。(Courchamp, Chapuis & Pascal, 2003)。特に、人間が持ち込んだ種に起因する捕食効果は深刻なものとなりうる(Silva-Rodríguez & Sieving, 2011;Doherty et al., 2016)。ラット(Rattus rattus)、ネコ(Felis catus)、イヌ(Canis lupus familiaris)のいずれも、最近の研究により最悪の脅威種と認識されている(Hughes & Macdonald, 2013)。

世界の自然界では、犬が約200種を脅かしており、そのうちのいくつかはIUCNの脅威カテゴリーに含まれているほどだ。同様に、野良猫とアカギツネ(Vulpes vulpes)の捕食プロセスは、オーストラリアの掘削哺乳類種の3分の2を減少または絶滅させた原因として記録されている(Fleming et al., 2014;Woinarski, Burbidge & Harrison, 2015)。掘削哺乳類がいなくなったことで土壌への撹乱が減り、有機物がほとんど土壌に溶け込まず、種子の発芽率が低い貧困な景観になった(Fleming et al., 2014)。アリューシャン列島では、導入されたホッキョクギツネ(Alopex lagopus)による海鳥の捕食により、栄養の投入と土壌の肥沃度が低下し、最終的には植生が草原から矮小低木林へと移行している。

最近の研究(Dannemann, Boyer & Miramontes, 2018)では、レビーウォークと生態系の回復力の間に深い関係があることが示された。この研究では、Dannemanと共同研究者は、レヴィーウォークの本質的なまだ未解明のマルチスケール移動特性を明らかにし、それがどのように集団ダイナミクスの安定性に重要な役割を果たすかを明らかにした。ロトカ-ボルテラモデルを用いて、一般的に拡散するフォレージャーは、脆弱な断片化生息地で絶滅する傾向があり、個体がレヴィーフライトを行っていれば、その集団は劣化した状況に対して回復力を持ち、存在量を最大化することを予測している。
レヴィ・ウォーク(Levy Walk)とレヴィ・フライト(Levy Flight)は、確率過程の一種で、ランダムウォークやブラウン運動と同様に、ある粒子や個体が無作為に移動するものである。これらの過程は、自然界や物理学、生物学、経済学などのさまざまな分野で観察される現象をモデル化するために使用される。
レヴィ・ウォークとレヴィ・フライトの主な違いは、移動の距離と時間のスケーリングに関連している。
レヴィ・ウォークでは、移動距離の確率分布が、パレート分布(またはその他の重いテール分布)に従う。これは、移動距離が短いことが非常に一般的である一方で、極端に長い距離を移動することも可能であることを意味する。レヴィ・ウォークでは、移動にかかる時間も考慮されるため、長い距離を移動する際には、それに応じて時間も長くかかることが特徴である。
一方、レヴィ・フライトでは、移動距離の確率分布もパレート分布に従うが、移動にかかる時間は考慮されない。これは、粒子が瞬時に長い距離を移動できることを意味する。したがって、レヴィ・フライトは無限の速さのジャンプを許容する過程であり、現実世界の物理的な制約を無視した理想化されたモデルと考えることができる。
レヴィ・ウォークとレヴィ・フライトは、異なるタイプのランダムウォークとして研究されており、検索戦略や動物の探索行動、株価の変動などのさまざまな現象を説明するために使用される。(by GPT-4)

この解析とシミュレーションの結果は、マルチスケール採餌の範囲を個体レベルから集団レベルへと変化させ、保全生物学の幅広い応用に大きな価値を与えるものである。その結果、レヴィフライトは探索と利用のバランスをとり、集団の安定性と回復力に寄与することがわかった。このように、現代の森林管理は、自然の撹乱を模した戦略的な伐採によって、この複雑な特徴(不均質性)を促進し、枯木や生木、そのままの森林のパッチを含むいくつかの構造や生物をそのまま残し、樹種の混在を促進することによって、より適合性が高まっている。これらの方法は、自然生態系や管理された生態系において、生物多様性を利用して収量や回復力を高めるという比較的最近の戦略に似ている(Filotas et al., 2014)。これらの考え方を一般化すると、生態系の完全性と回復力を維持するために、管理システムは最低レベルの生態系の複雑性を維持することを考慮すべきことを示唆する重要な証拠があると考えられる。
もちろん、このことは複雑さをどのように測定するかという新しい課題を提起している。Gershenson & Heylighen (2003)に従って、シャノン情報を用いて複雑さを測定することができる。この情報理論の枠組みでは、新しい情報を得るためには、古い情報が変換される必要がある。また、複雑系の特徴である自己組織化(S)は、秩序の増大(エントロピーの減少)と相関している(Gershenson & Heylighen, 2003)。したがって、もし創発がエントロピーに類似した情報の増加を意味するのであれば、自己組織化は創発と反相関するような形で
S=1-I=1-E (12)
このようにして、次の(Gershenson & Heylighen, 2003;Fernández, Maldonado & Gershenson, 2014)複雑さは、次のように測定することができる。
C=4・E・S (13)
複雑系の視点に立つと、生態系は単純にトップダウンで管理できるようなシステムではない。相互作用が複数の階層的なレベルで行われることを明示的に考慮しなければならない。これは複雑系の一般的な特徴であり、構成要素は階層的に組織化され、異なるレベルの要素が相互作用して、システムを特徴づけるアーキテクチャを形成している。このように、複雑性は、あるスケールで起こる現象は、スケールを超えた相互作用を考慮しなければ理解できないことを主張する。しかし、それは同時に、環境政策、管理、介入をスケールの観点から見直す必要があることも意味している。この点で、Talebは”Principia Politica”(草稿はwww.academia.edu/38433249/Principia_Politica)で、政策をスケール依存と理解することを盛り込んでいる。したがって、生態系管理のための一枚岩の政策を目指すのではなく、異なるレベルの適用と関連した一連の政策を開発することを検討する必要がある。このようなアプローチは、壊滅的な隠れた影響のリスクを軽減するために必要である。
自然と人間のサブシステムの結合を理解することは、管理に挑戦する全く新しい物語を提供する。生態系の管理は、意思決定者、科学者、管理者、関係する市民など、さまざまな主体による集団行動の結果である。複雑さ(生態系の完全性と回復力の鍵)は、創発と自己組織化(S)の間の動的バランスであるため(式(2)と(3)、いくつかの(そして正しいタイプの)自己組織化は育成される必要があるが、多すぎるのは良くないことである。多すぎる(間違ったタイプの)Sは、有害な結果をもたらす望ましくないフィードバックを持続させるかもしれない。例えば、ボルネオ島の違法伐採は、ステークホルダー階層のあらゆるレベルの相互作用に支えられた自己組織化現象と見ることができる(Puts & Chattaraj, 2013)。そのメカニズムはFilotasと共同研究者により説明されている。

フィードバックは、竪穴式製材業者が生計を立て、海賊版伐採業者が統治の失敗を利用することから始まる。これだけでは、悪徳な木材バイヤーや違法木材を洗浄する腐敗した政府関係者と組み合わさらない限り、これほど大きな影響を与えることはできない。メキシコでの経験から、腐敗は生態系の劣化という重要なプロセスにおける共通のリンクである可能性がある。最後に、不正に入手した商品に有利な出口を提供する、より高度なリンクとして、精通した国際トレーダーが存在するという。著者の説明によると、違法伐採が行われる場所では、木材市場が氾濫し、木材価格は下落し、立木は過小評価される。このような状況では、コミュニティの森林管理者は、長期的なリターンのために短期的な投資を伴うことが多い、持続可能な森林管理を実施する意欲を持てない。このような状況は、違法伐採を維持するための自己組織的なフィードバックにつながる。
複雑系の特徴である「固有の記述尺度を持たない」という性質は、最も普遍的なシステム全体の現象の1つである、ゆらぎ時系列の周波数空間上の1/f挙動に関連している。このいわゆるピンクノイズは、βスケーリング指数によって定義される1/fβ色ノイズまたはフラクタルノイズのファミリーの一つであり、臨界指紋と考えられている。揺らぎのダイナミクスを3つの典型的なノイズグループ、ホワイトノイズ(β〜0)、ピンクノイズ(β〜1)、ブラウンノイズ(β〜2)との類似性によって比較・分類することが一般的である(Bak, Tang & Wiesenfeld, 1988;Landa et al, 2011;Kleinen, Held & Petschel-Held, 2003;Peng et al., 1994)。臨界性の普遍性については現在も検討されており、秩序と無秩序の間を行き来する動的システムにおけるシステムは、堅牢性と柔軟性のバランスに達したときに最大レベルの計算能力に達するという「臨界性仮説」として知られている(Roli et al., 2018and references therein参照)。この臨界性の考え方は、最近、生態系の健全性と持続可能性を定義する情報理論的アプローチに用いられた(Ramírez-Carrillo et al., 2018)。この論文では、Ramírez-Carrilloら(2018)は、スケール不変性(パワースペクトルにおけるべき乗則として)と適応性と頑健性のバランスの混合として、生態系が臨界にある場合、健全であると考察する。
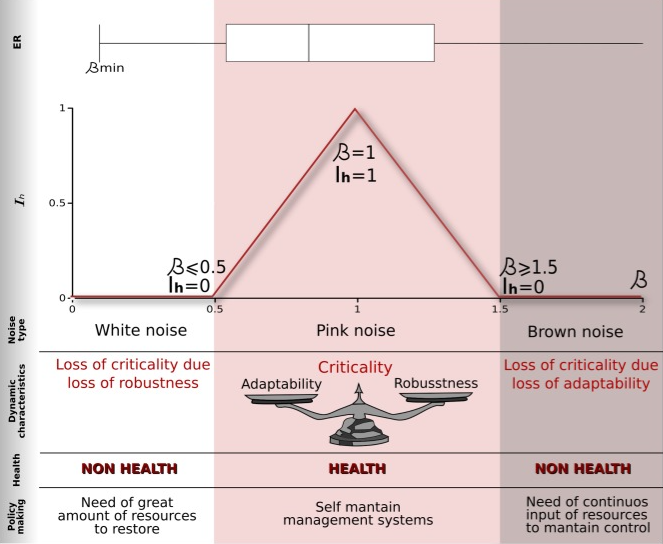
スケール不変性とピンクノイズの挙動で定義される臨界性の利用が、系統的な生態系の健全性を測る一つの方法である可能性を示した。非動的な変数(この場合は土地条件)を用いて分析を補完した。一般に、ピンクノイズ(β〜-1)の挙動を持つサイトは、ホワイトノイズ(β〜0)やブラウンノイズ(β〜-2)を持つサイトよりも、統計的に良好な土地状態であることがわかった。興味深いことに、土地の状態が良くてもシステムの健全性が低い場合や、その逆の場合もある。
私たちは、土地の状態の値が高く、生態系の健康の値が低いという最初のケースを、若いアスリートの心臓突然死のような現象を考慮した人間の健康との類似性という観点から解釈する[100]。
20代のオリンピック選手を考えてみよう。外見的な健康状態がこれほど良い人はなかなかいないと思うが、この選手は、全身的な健康状態に関係するこの症候群のために、トラック上で死んでしまうかもしれない。

一方、腕が折れている(外見上は明らかに不健康のシグナル)にもかかわらず、心臓、脳、一般的なシステム機能の点では完全に健康である人を考えてみよう。さらに、集中的に管理された生態系の第3のケースとして、この例えのように、生命維持装置を用いて何らかの形で人工的な健康状態を維持している集中治療中の患者を想定している。
これらのべき乗則は、Ramírez-Carrilloら(2018)、Mandelbrot(1982)、Newman(2005)、Sornette(2006)がまとめたように、地震統計、太陽フレア、疫病発生など多数の現象に現れている。また、生物学でも共通のテーマとなっている(Goldberger, 1992;Goldberger, Peng & Lipsitz, 2002b;Gisiger, 2001;West, 2010)。複数の研究者が、心臓活動などの生理的プロセスにおける動的臨界の証拠を報告し、それが健康な状態の主な特徴になり得ることを示唆した(Kiyono et al., 2005;Ivanov et al., 1996;Rivera et al., 2016a)。いくつかの研究(Goldberger, Peng & Lipsitz, 2002b;Rivera et al., 2016b)では、健康な心臓と1/f領域前後のスケール不変のノイズとの間に、医学的根拠に裏打ちされた関係性があることを示す有力な証拠を発見した。
この複雑性のアプローチは、明らかに生態系の完全性の物語を補完するものであり、EVLの値を「安全な」範囲内に維持することに加えて、重要な動的領域(スケール不変性と「1/f」揺らぎ)内に維持する場合、生態系は弾力性があると考えるべきである(Ramírez-Carrillo et al, 2018).
レジリエンスを超えて、アンチフラジリティ
生命システムは、単にランダムな突然変異とそれに続く選択によって環境の変動に反応するだけではなく、周囲の変動を発見し、逆境、変動、不確実性に対処できるような特性を内蔵していることがほとんどである。反脆弱性は、このような特性の一つであり、もしかしたら中核をなすものかもしれない(Danchin, Binder & Noria, 2011;Taleb, 2012)。
何かが壊れやすいというのはどういうことなのかを考えてみると、重要な特性は、環境の変動によってダメージを受けるということである。では、「壊れやすい」の反対語は何かと、身近な同僚に適当に聞いてみると、たいていの場合、「堅牢性」や「回復力」といった概念が出てくるだろう。しかし、よくよく考えてみると、そのどれもが「壊れやすい」の正反対ではないことは明らかだ。どちらも、環境の変動に鈍感なシステム、あるいは一瞬だけ影響を受け、すぐに初期状態に戻るシステムを表している。
フラジリティ(脆弱性)の正反対をタレブはアンチフラジリティ(反脆弱性)と定義し、外部からの摂動に答えるシステムの機能的能力を高める性質としている(Taleb, 2018)。言い換えれば、環境の変動性から恩恵を受け、撹乱された後によりよく機能する場合、システムは反脆弱であると言える。そうすると、反脆弱性は、堅牢性や回復力を超えていることになる。堅牢性・回復力のあるシステムは、ストレスに耐えて同じ状態を保つが、反脆弱性のある構造は、ストレスに耐えるだけでなく、ストレスから利益を得て学習・適応する。免疫系は、反脆弱性システムの重要な例となる。若い頃にさまざまな細菌にさらされると、免疫系は改善され、将来的に新しい病気を克服するためのさまざまな能力を獲得する(Pineda, Kim & Gershenson, 2018)。
ペイオフ空間における凸性としての反脆弱性の正式な定義は、Taleb & Douady (2013)とTaleb (2018)にある。2回連続微分可能な「応答」またはペイオフ関数f(x)を考えよう。すると、その関数の凸性は次の関係で定義される。∂ 2f ∂ x 2 ≥0となり、適切な条件のもとで簡略化できる。12 [f(x+Δx)+f(x-Δx)]≧f(x).すると、応答関数fは用量に対して非直線性を示すことになり、このことは、用量の増加に対して、より高い影響を与えることを意味する。Talebはこの結果を、以下のような線形結合に一般化した。∑αi=1,0≤αi≤1ように∑[αif(xi)]≧f[∑(α i x i )]。.再び議論を単純化すると、正しい条件のもとでは、f(nx) ≧nf(x)となる。このように、Xが関数fがよくふるまう支持を持つ確率変数で、fが凸である場合、Jensenの不等式が得られる。
E(f(x)≧f (E(x)) (14)
一般性を損なわず、密度φ(x)、サポートinを持つその連続分布が、φ(x/σ)、σ>0の位置スケール族分布に属するとすれば、次のようになる。Eσは、スケールσを指標とする確率分布の下での期待値を表す写像で、次のようになる:
∀σ2>σ1,Eσ2[f(x)]≥Eσ1[f(x)] (15)
このように、Talebは局所的な反脆弱性を「特定の区間において、式3のように分布の規模に応じて期待値が増加するか、同じ区間で用量反応が凸になる状況」と定義している。
アンチフラグリティの枠組みは、Talebによって金融リスク分析の文脈で開発されたが、その普遍的な数学的形式により、注目を集め、本来の範囲から遠く離れた場所でも適用されている。アンチフラグリティの概念は、分子生物学から都市計画まで応用されている(Pineda, Kim & Gershenson, 2018およびその中の参考文献を参照)。Pinedaと共同研究者(Pineda, Kim & Gershenson, 2018)は、システムの複雑さをペイオフ関数として定義することで、antifragilityの簡単な実装を提案した。これは、複雑さが批判的に、したがって堅牢性と適応性の間のトレードオフバランスと非常に関連しているので、私たちのレビューの文脈では非常に理にかなっている。
著者らは、フラジリティを次のように定義した。
∮=-ΔC|Δx|. (16)
ここで、ΔCは、次数の摂動によるシステムの複雑さの変化である|Δx|.複雑さは常に正規化されるため、∮の正の値は壊れやすいシステムを定義し、∮がゼロの場合、システムは堅牢/回復力があり、∮の負の値ではシステムは反脆弱であることを示す。
そして、Pineda, Kim & Gershenson (2018)は、それを遺伝的制御の働きのモデルのランダムブーリアンネットワーク(RBN)に適用した。彼らは、順序付きRBNが最も反脆弱であることを発見し、予想通り、CD4+ T細胞の分化と可塑性やシロイヌナズナの細胞周期など、生物学的によく研究された7つのネットワークが反脆弱であることを実証した。
中心極限理論から、正規分布は相互作用(確率的独立性)のない(単純な)系からしか生まれないことが分かっている。相互作用を考慮すると(確率的独立性がない)、対応する確率分布はファットテールを持つことになる。その意味で、複雑性はファットテール、ファットテールは脆弱性/反脆弱性と関連している(Taleb, 2012)。
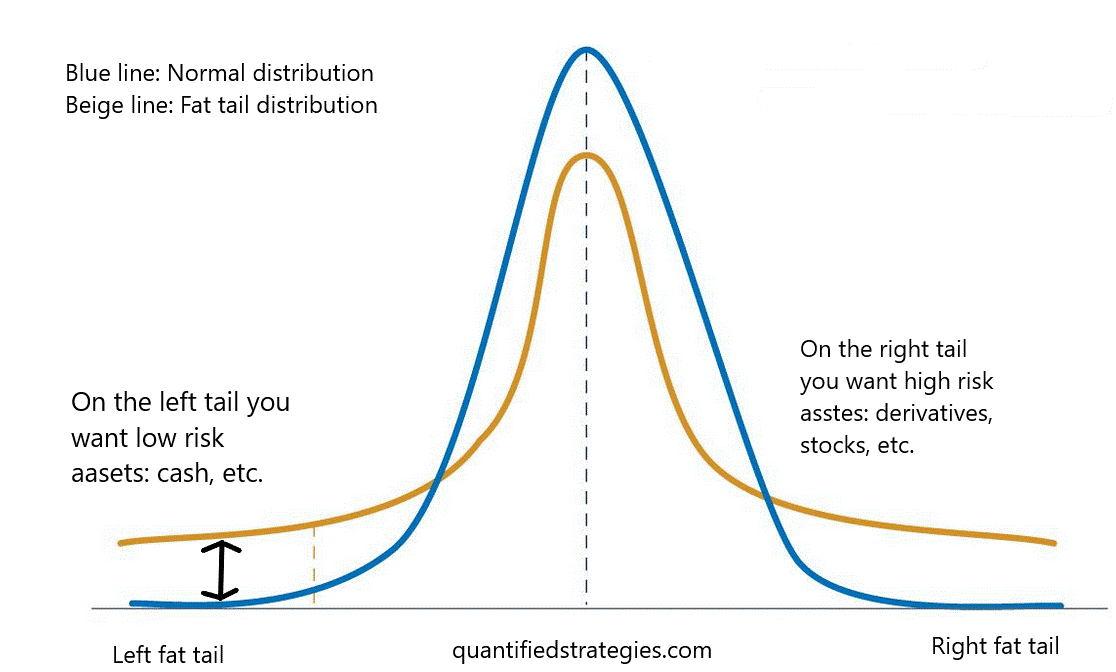
Talebの説明では、応答関数の正規分布は頑健なシステムを特徴づけるが、左の太い尾は壊れやすく、右の太い尾は反脆弱なシステムである。最も興味深いのは、Fossion, Rivera & Estanol (2018)が、ホメオスタシス(生理的回復力?)を、制御すべきもの(ホメオスタシスを維持するもの)と前者を制御する別のものの生理変数のペアと関連付けたことである。その主旨は、恒常性を保つ生理的変数(正常)を持つためには、他の変数(右太もも)を使って、環境から物質、エネルギー、情報、あるいはそれらの任意の組み合わせがランダムに注入されても、それを吸収する必要があるというものである。Fossion, Rivera & Estanol(2018)は、彼らの論文の図4において、(a)健康なコントロール(複数)、(b)最近診断された糖尿病患者(複数)、(c)長年の糖尿病患者(複数)についての心拍HRと血圧BPの変動解析の結果を示している。その結果、健常者では血圧は正常で、HRは右の脂肪尾状であることがわかった。糖尿病と診断されたばかりの患者では、BPは正常値を失い、左の尾を引くようになり、HRは正常値に戻る傾向がある。やがて、長年の糖尿病患者の場合、血圧は明らかに左の脂肪尾を持つようになり、HRは正常となる。これは、人間の健康における抗脆弱性の役割について、非常に説得力のある証拠である。
一般論として、Taleb (2012)は、反脆弱性を実現するための典型的な戦略として、いわゆるバーベル(またはバイモーダル)アプローチを提案している。アンチフラジリティを実現するための第一歩は、アップサイドを増やすのではなく、ダウンサイドを減らすことである。言い換えれば、確率は低いがペイオフが大きい不利な事象(すなわち、「ブラックスワン」事象)への曝露を減らし、自然のアンチフラジリティが自ら機能するようにすることである。
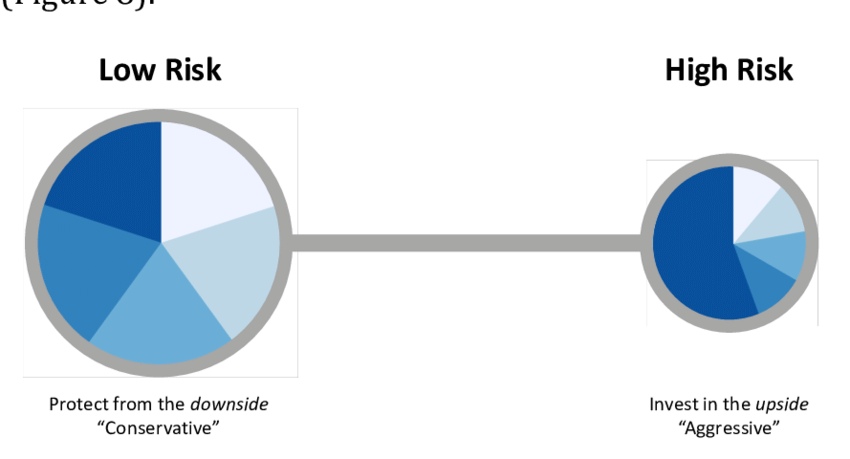
私たちは、Talebに続いて、その考え方が最も説明しやすい、低俗な金融の事例を紹介する(ただし、その多くは誤解されている)。金融におけるバーベル・アプローチは、自分の資源の90%を安全な商品(インフレから保護されていることが条件)、あるいは「価値リポジトリ数」と呼ばれるものに置き、10%を非常にリスクの高い、最大リスクの債券に置き、凸の方法で予測できない巨額の利益に自分をさらすことからなる。このような形で、ある種の二元最適化が行われ、安全な逆張りの堅牢性と、高リスクな逆張りの適応性が同時に利用されることになる。いわゆる「中位」の証券に100%投資している人(一元最適化)は、「完全なリスク破滅」の危険性がある。このバーベル戦略では、異常事態の危険性の評価における計算の非簡約性と脆弱性の問題を取り上げている。
反脆弱性を実現する基本的なメカニズムであるバーベル戦略は、ファット分布下での徹底したリスク管理戦略であり、自然界に広く存在することから、自然システムにおいて非常にユビキタスであることが期待できる。私たちは、このバーベルリスク戦略を、ネットワークトポロジーにおける強弱相互作用や出世と俯瞰の関係による「良いバランス」特性と、システムの揺らぎのダイナミズムにおける批判的(スケール不変と1/f型ノイズ)の指紋として識別される堅牢性と適応性のバランスとして特定する。この3つの「良いバランス」がすべて関連していることは明らかであり、本文の冒頭で述べたように、これら3種類のバランスはすべて、生命システムのより一般的な進化戦略である「抗脆弱性」の特殊事例であるという可能性が非常に高い。
考察
文献の分析から、査読付き論文からの引用ネットワークが浸透していないことがわかった。これは、研究分野が統一されておらず、学際的な研究を行う機会になっていると解釈している(補足資料参照)。
私たちは、(a)レジリエンスを高めるための生態系の特性、(b)摂動に対する生態系の反応、(c)複雑性の3つの主要な物語を発見した(図2参照)。このほか、参考にした文献から、生態系の回復力を示す11の指標を特定した(表3、表4の用語集を参照)。特に、フィッシャー情報は、確固たる形式的枠組みを持ち、実装が容易で、あらゆる種類のシステムに適用できるため、レジリエンスの非常に有望な代用品であると考えられる研究事例で、その適用方法を示した。
表3 文献レビューや補完的な論文に見られるレジリエンス対策
| キー | インジケーター | メジャー/プロキシ | 必要なもの | レジリエンス | ナラティブ |
|---|---|---|---|---|---|
| フィー | フィッシャー情報 | 安定性 | 時系列 | Cabezas et al. (2005)によれば、生態系が回復力を持つためには、擾乱の後、それ以前のFI値が回復している必要がある。 | 摂動 |
| ディヴ | 多様性 | オプション/リソーススペースの使用。 | プレゼンスフィールドデータ | 一般的に、多様性が高ければ高いほど、回復力も高くなる。しかし、構成や資源の利用の変化に関連する例外もある。 | 物件紹介 |
| コ | ネットワークコネクタンス | 安定性 | ネットワークを知り、つながりの強さを数値化できるようになったグスタフソンは、それに関する情報の不足に対処する方法を提案する。 | 接続数の増加により、種の分布のばらつきの影響が解消され、安定した種が強化される | 物件紹介 |
| オーム | 雑食性生物種の存在 | 異なるスケール間のコミュニケーション | 雑食性生物種が存在すること | 雑食動物の存在が安定性と回復力を高める | 物件紹介 |
| 数値制御 | ネットワークの重要性 | 堅牢性(強い相互作用)と適応性(弱い相互作用)のバランス。 | ネットワークを知り、つながりの強さを数値化できるようになったグスタフソンは、それに関する情報の不足に対処する方法を提案する。 | 生態系は、強いつながりと弱いつながりの数がバランスよく存在すると、より回復力が高まることが観察されている | 物件紹介 |
| エルブイシー | ロトカ・ヴォルテラ係数 | コミュニティ行列が与えられたとき、その実数部がすべて揃っていれば 固有値が負である場合、生態系は安定している。 |
コミュニティーマトリクス | より安定した生態系は、より弾力的である | 複雑さ |
| として | 昇天 | 平均相互情報量 | 相互作用のネットワーク(栄養ネットワーク)がある場合、ネットワークが2つのノード間のフローイベントを平均してどれだけうまく表現できるかを測定する。 | 生態系がその組織と規模の組み合わせにより、外乱に打ち勝つ能力を一つの指標としてとらえる。 | 物件紹介 |
| レヴィ | レビー便 | ピューマやジャガーなどの重要種の採食パターンのスケール係数 | リソースの空間的な複雑さの代用品である | レビー便の採餌パターンが関連し、生態系の回復力を高めることが明らかにされている | 複雑さ |
| フラック | フラクタル | 空間的な複雑さ | 高解像度衛星画像 | より複雑な生態系は、より弾力的であるべきである。 | 複雑さ |
| かちょうしゅうはすう | 反脆弱性(Antifragility | 生物(栄養)ネットワークの複雑さの変化、外乱に直面したときの変化 | 相互作用のネットワーク。例えば、ピューマやジャガーなどの主要な種とその獲物との共起のブールネットワークとすることができる。 | レジリエンスは、フラジリティとアンチフラジリティの中間的な状態であろう。 | 摂動 |
| H | ホメオスタシス | システムのホメオスタシス | 時系列 | レジリエンスに相当するもの | 複雑さ |
表4 この記事で使用されている一般的でない用語の用語集
| 期間 | 定義 |
|---|---|
| 反脆弱性(Antifragility | Antifragilityとは、外部の摂動に対応するシステムの機能的能力を高める特性である(Taleb, 2018)。つまり、あるシステムが環境の変動性から恩恵を受け、撹乱された後によりよく機能する場合、Antifragileであると言える |
| 昇天 | 生態系のネットワークの枠組みを通る情報の流れの大きさを表す指標である。 |
| 複雑さ | システムが複雑であるのは、十分な数の構成要素が十分に強い相互作用を示すか、観察者の時間スケールに匹敵する速度で変化するか、ほとんどの場合、その両方である。システムとしての森林と森林管理は、複雑さの勾配において高い位置を占めていることは確かである(Filotas et al., 2014)。それは、創発と自己組織化の産物として測られる |
| クリティカル | 臨界とは、システムが動的なスケール不変性(周波数空間におけるべき乗則)を持ち、堅牢性と適応性の「最適」なバランス(スケール係数が-1程度)にある領域である |
| エマージェンシー | 情報出現率Eは、情報の変換速度として定義することができる。シャノン情報として測定することができる |
| フィッシャー情報 | フィッシャー情報は、測定-推論プロセスの品質と理解することができる。位相空間における接線速度や加速度、ひいては安定性に関係する。 |
| ホメオスタシス | Fossionと共同研究者(Fossion, Rivera & Estanol, 2018)は、ホメオスタシス(生理的回復力か)を、制御すべきもの(ホメオスタシスを維持するもの)と前者を制御する別のものの生理変数の対に関連付けた。主な考え方は、ホメオスタシスな生理的変数(正常)を持つために、身体は他の変数(右太尾)を使って、環境から物質、エネルギー、情報、またはそれらの任意の組み合わせのランダムな注入を吸収しなければならないということである |
| 誠実さ | 生態系の構造、構成、機能の観点から、生態系の状態を示す指標である |
| レヴィフライト | 局所的な空間探索を特徴とするファットテイル型採餌パターン(正規分布)、非局所探索のための大きな「飛行」もあり |
| 永続性 | 持続性とは、ある変数が異なる状態に変化する前に、同じ状態を維持する時間のことである(Pimm & Pimm, 1991)。永続性とは、システムが時間と共にそれ自身を維持する能力の尺度である(Loreau et al., 2002) |
| ロバスト性 | ロバストネスとは、環境の安定性の耐久性に関係する。そして、ロバスト性は、生態系が異なる状態に変化する前に、どの程度の撹乱に耐えられるかを示す尺度である(Loreau et al., 2002)。食物網がより堅牢であればあるほど、より安定しているといえる |
| 自己組織化 | エマージェンシーの補数(1-E)であり、システムの組織力を高める能力を表す。 |
それにもかかわらず、この批判的な文献レビューから、レジリエンスを再解釈する新しい方法が浮かび上がった:アンチフラジリティ(Antifragility)である。Taleb (2012)、Taleb & Douady (2013)、Taleb (2018)が開発したこの斬新なフレームワークは、変動性に対するシステムの脂肪尾状の非線形応答に基づいている(図4参照)。簡単に言うと、ある変数に依存するペイオフ関数が凹型(非線形)であれば、そのシステムはそれに対して脆弱である。逆に、ペイオフが凸であれば、それはAntifragileであり、システムが本質的に変動に鈍感であれば、Robust/Resilientである。Talebの研究では、反脆弱性は「バーベル」と呼ばれる二峰性のリスク戦略と関連しており、私たちは、(i)ネットワークトポロジーにおける強い相互作用と弱い相互作用、(ii)適応性と堅牢性(Criticality)、(iii)出世とオーバーヘッド、の「良いバランス」として現れると考えている。

図4 摂動や変動に対して凸状に応答するシステムの特性である「反脆弱性」の観点から、システムの基本的な特性を説明する。
(A-C)はそれぞれ壊れやすいシステム、ロバスト/レジリエントシステム、アンチフラジャイルシステムの例、(D-F)は摂動に対するプロファイル応答の例、(J-L)は典型的な確率分布の例、(M-O)は複雑性変化に基づくメトリックで得られる特性値である。
実際の生態系反脆弱性は、(Ramírez-Carrillo et al., 2018)のように時系列順列エントロピーを用いて、Gershenson & Fernández (2012)の複雑性公式を用いて、ここに提示した複雑性に基づく指標を用いることができる。この直接的な指標に加えて、原稿の査読者の一人が指摘したように、生態系の状態変数分布における右肩下がりが、生態系の反脆弱性の一つの指標になる可能性があると考える。この直感は、Fossion, Rivera & Estanol (2018)による最近の研究から得られたもので、彼らは、人間のシステムにおいて(しかし私たちは一般的な結果であると考える)、血圧などの生物にとって重要な状態変数は、ガウス分布に従って非常に鋭い値の範囲に維持される必要があると提案している。これは、従来のホメオスタシスの考え方と一致するものであり、目新しいものではないが、これを実現するためには、生物が環境からエネルギー、物質、情報の揺らぎを吸収する必要があるとした点である。この吸収や制御のプロセスは、心拍数の変動によって行われる。このように、若さと健康は、恒常的な生理学的変数のガシアン性と、吸収(制御)的変数の右肩下がりの特徴を持っている。著者らは、糖尿病などの慢性疾患下では、その統計的挙動がどのように反転するかを示している:血圧(恒常性変数)は左歪度分布、心拍数(吸収変数)はガウス型になる(Fossion, Rivera & Estanol, 2018).いくつかの問題は、恒常性変数と吸収変数のペア(またはネットワーク)を特定すること、そして歪度が脆弱性以外の他のプロセスによって影響される可能性があるかどうかであろう。
私たちの主な関心は情報理論的アプローチにあったが、査読の過程で、情報理論を挟まずにIntegrityとResilienceが相互作用する論文をもっと取り込むために、検索基準の範囲を拡大することが提案された。そこで、「Resilience AND Integrity AND Ecosystem」を用いた新たな検索を行ったが、これらのキーワードがタイトルに含まれていることを条件とし、これらのキーワードに焦点を当てた論文であることを確認した。その結果、Müller, Burkhard & Kroll, 2009;Bratanova-Doncheva et al., 2014;Chipev et al. (2013)の3つの論文が新たに見つかり、先に確認したシナリオに該当することがわかった。
最初のものは、生態系のダイナミクスを理解するために、生態系が摂動にどのように反応するかを研究することの重要性について興味深い議論を展開している。著者らは、例えば、30年代に複雑性(当時は自己組織化として理解されていた)が安定性の中核的な前兆であることが確認された研究に基づき、このテーマの歴史的進化をレビューしている。50-60年代になると、著者たちは、安定性、抵抗力、堅牢性という3つの主要な構成要素を通じて、均衡の重要性を強調する(これらの用語の良い説明については、Graftonら(2019)の図1参照)。60~70年代の文献では、脆弱性、脆弱性、経路依存性、局所的不安定性に言及し始め、80~90年代には不平衡に至り、ホリングのレジリエンスの視点に置き換わったと言う。著者らは、レジリエンスと生態系のダイナミクスは、生態系の発展段階によって変化するという考えを提唱している。この考えは、生態系の発展段階は、破壊されていない連続体を通して構成要素の最適化(オリエンター)に支配されたダイナミクス状態から始まる。この文脈では、完全性はオリエンタルの発達を示すと主張するが、完全性の評価は、生態系が摂動にどのように応答するかについて情報を提供しないため、生態系のダイナミクスを完全に特徴付けるものではないと警告している。さらに、例えば、一般的に、相互依存関係が最大となるような高度な成熟度が達成されると、制約条件の変化に対する適応能力が低下する可能性があると結論づけることができる。これは、人間による生態系でも観察される特徴であり、高齢の個体は創発損失によって臨界から離れ、自己組織化が進みすぎる(Goldberger et al, 2002a)。
他の2つの論文は、完全性とレジリエンスを生態系サービスに関連付けるもので、生態系の特性に関する説明に該当するものである。生態系サービスは別の物語と考えることもできるが、それは実際には生態系ではなく、社会生態系を対象として定義されている。したがって、生態系サービスは、次のセクションで簡単に説明する社会システムレジリエンスの文脈の中で組み立てられた物語であるべきである。
結論
長期的には、生態系と人間システムの結合(気候変動など)を考慮すると、レジリエンスよりもアンチフラジリティの方がより望ましい機能であると考えられる。社会生態系を考えてみると、気候危機や土地の変化のような大きなストレス要因があっても、社会生態系は生き続けるだけでなく、繁栄し進化していくのが普通であることがわかる。実際、最近の研究(De Bruijn, Größler & Videira, 2019)では、設計基準として反脆弱性を用いることで、ブラックスワン(予測不可能で発生頻度は非常に低いがインパクトが非常に大きい事象)の影響を受けやすい環境において、研究対象のスキームは「単に」ロバストモデルよりも好ましい挙動を示していることが示されている。そうすると、社会生態系のガバナンスや計画、あるいは一般的な意思決定の観点からは、反脆弱性は、レジリエンスの願望よりも価値ある、達成すべき望ましい目標かもしれない(Blečić & Cecchini, 2018)。一方、ランダム性やストレス要因の抑制といった自然システムへの素朴な介入主義は、Taleb (2012)が広く論じているように、脆弱化プロセスにつながる可能性がある。Fernandez-Oto、Tzuk & Meron (2019)が、非線形フロント不安定性の存在下で、砂漠化からの自己回復プロセスに入る可能性がある乾燥地生態系で示したように、生命システムは繁栄するために適切な量のランダム性を必要とする。このことは、私たちに疑問を抱かせます:臨界性、反脆弱性、フィッシャー情報の関係はどのようなものなのか、また、反脆弱性の概念は、例えば、Cabezasら(2005)やRamírez-Carrilloら(2018)の持続可能性のフレームワークにはどのように入るのか。
資金調達
本研究は、CONACyT基金M0037-2018-07、番号296842、Cátedras CONACyTフェローシッププログラム(プロジェクト番号30)、Sistema Nacional de Investigadores SNI、番号62929による支援を受けた。資金提供者は、研究デザイン、データ収集と分析、発表の決定、原稿の作成には一切関与していない。