

Contents
CYPフォーム
脳のシトクロムP450
シトクロムP450は酵素は、細菌、植物、哺乳動物などほとんどすべての生物に存在する酸化酵素。
一般的に薬物、有機化合物など生体の異物を代謝させて排出する解毒プロセスの第一段階を担当する解毒酵素として認識されている。
しかし、これらの酵素は多くの内因性の基質を代謝し、生理学的に重要な役割も果たしている。
脳内でもさまざまなCYPアイソフォームが発現しており、コレステロールの代謝サイクル、ドーパミン、セロトニン、モルヒネ、ホルモン、保護作用をもつ脂質中間体の生合成に関与する。これまでの研究から、これらの酵素群が脳の恒常性維持に不可欠であるという証拠が積み重なっている。
潜在的には、すべてのCYP酵素が神経変性疾患と関連する可能性があるが、研究で神経変性疾患と関連づけられているCYP酵素は、以下の通り。
- CYP46A1 脳内コレステロールの排出
- CYP17A1 プレグネノロン-DHEA代謝
- CYP19A1 アロマターゼ酵素の形成
- CYP2C19(阻害)アラキドン酸の代謝物抑制による保護効果
- CYP2D6 ドーパミン代謝(パーキンソン病)
- CYP2E1 MPTP毒素の解毒(パーキンソン病)
ラットの脳ではシトクロムP450の含有量は肝臓の3%であり、30倍低い活性を有する。
脳内のCYP発現は不均一であり、一部のCYP(CYP46A1、CYP2D6)は肝臓よりも脳内でより大きな割合で発現する。
CYP1A1、2B、2E1、3Aなどの一部のアイソフォームは主にニューロンで見られるが、CYP2D6などの他のアイソフォームはニューロンとグリア細胞の両方で見られる。
CYP1Aファミリー
CYP1Aファミリーは、発がん性物質、ホルモン、医薬品の代謝に関与する。
多環芳香族炭化水素(PAH)の発癌性生物活性、複素環式芳香族アミン/アミド、ポリ塩化ビフェニル(PCB)、他の環境毒素と関連することが知られている。
www.ncbi.nlm.nih.gov/pubmed/17431034/
www.ncbi.nlm.nih.gov/pubmed/19255595/
CYP1A酵素群は、フェーズⅡと協調されずに過剰活性されると発がん性をもつ環境因子の毒性効果を高める可能性がある。
www.ncbi.nlm.nih.gov/pubmed/19581216/
CYP1酵素(+CYP1B1)は、エストロゲン代謝産物の形成にも関与する。
www.ncbi.nlm.nih.gov/pubmed/16112414/
CYP1A1
誘導剤
レスベラトロール
前臨床試験 レスベラトロールによるCYP1A1の促進
www.ncbi.nlm.nih.gov/pubmed/20716633
クルクミン
www.ncbi.nlm.nih.gov/pubmed/24501322
www.ncbi.nlm.nih.gov/pubmed/17044766
大豆
www.ncbi.nlm.nih.gov/pubmed/25011215/
ニンニク
www.ncbi.nlm.nih.gov/pubmed/12575903
フィッシュオイル(高用量)
www.ncbi.nlm.nih.gov/pubmed/11498267
ローズマリー
www.ncbi.nlm.nih.gov/pubmed/11498267
アスタキサンチン、酵母、オキアミ
www.ncbi.nlm.nih.gov/pubmed/8851821
阻害剤
エラグ酸・ベリー類
www.hindawi.com/journals/jnme/2015/760689/
グレープフルーツジュース
www.ncbi.nlm.nih.gov/pubmed/27444380
トマト、人参に含まれる赤い色素
www.ncbi.nlm.nih.gov/pubmed/20400267
スルフォラファン(ブロッコリー)
www.ncbi.nlm.nih.gov/pubmed/23566952
緑茶抽出物
www.ncbi.nlm.nih.gov/pubmed/11064004
ローズマリー
www.ncbi.nlm.nih.gov/pmc/articles/PMC3324618/
CYP1A2
カフェインを代謝する酵素として有名。CYP1A2の発現はタバコの煙に含まれる多環芳香族炭化水素(PAH)によって誘導される。
他の生体異物基質にはアフラトキシンB1、パラセタモール(アセトアミノフェン)が含まれる。
CYP1A2の活性は、精巣腫瘍リスクの増加と関連する。
www.ncbi.nlm.nih.gov/pubmed/14976127/
誘導剤
アブラナ科野菜
(インドール3カルビノールのCYP1A2活性)
www.ncbi.nlm.nih.gov/pubmed/2342128
www.ncbi.nlm.nih.gov/pubmed/19843669
www.ncbi.nlm.nih.gov/pubmed/12147290
緑茶
www.ncbi.nlm.nih.gov/pubmed/24815822
www.ncbi.nlm.nih.gov/pubmed/7955108
活性の阻害
ケルセチン
ケール
ニンニク
にんにくオイルはマウスのCYP1A2を抑制する。
www.ncbi.nlm.nih.gov/pubmed/19875617
大豆(ダイゼイン)
www.ncbi.nlm.nih.gov/pubmed/12756512
グレープフルーツ、ナリンゲニン
300mlのグレープフルーツジュースとナリンゲニンはヒトのCYP1A2活性を阻害する
www.ncbi.nlm.nih.gov/pubmed/12756512
CYP1B1
多環式芳香族炭化水素や17βエストラジオールなどの発癌物質を代謝する。
ルイボス茶、セロリ(クリソエリオール)によるCYP1B1の選択的阻害
www.ncbi.nlm.nih.gov/pubmed/23448447
Cyp1B1発現はNF-κB活性を抑制することにより血管新生を促進する。
www.ncbi.nlm.nih.gov/pmc/articles/PMC3882387/
CYP2Aファミリー
CYP2ファミリーは、薬物、生体異物、ホルモン、ケトン類、グリセロール、脂肪酸などの他の内因性化合物の代謝に関与する。
www.ncbi.nlm.nih.gov/pubmed/12369887/
CYP2A6
ニコチン、クマリンの代謝に関わるCYP酵素
ケルセチン 、りんご、アプリコット、ブルーベリー、黄タマネギ、ケール、アルファルファもやし、インゲン、ブロッコリー、紅茶、チリパウダー 、ブロッコリー
誘導剤
クルクミン
www.ncbi.nlm.nih.gov/pubmed/20484172
阻害剤
アーモンド抽出物
www.ncbi.nlm.nih.gov/pubmed/12746108
シナモン精油(シンナムアルデヒド)
www.ncbi.nlm.nih.gov/pmc/articles/PMC4810772/
ホワイトグレープフルーツ
www.ncbi.nlm.nih.gov/pubmed/17112808
CYP2B6
薬剤の排出
www.ncbi.nlm.nih.gov/pubmed/23467454
誘導剤
阻害剤
甘草(グラブリジン)(強力)
herbpedia.wdfiles.com/local–files/attachments/herbal_bioactivation_mini_review.pdf
ボルネオール(マンゴージンジャーオイル)
CYP2C8
CYP2C8は、肝臓CYPの7%を占める。
CYP2C8ハプロタイプ構造にはかなりの人種的多様性がある
ロシグリタゾン、ピオグリタゾンはCYP2C8 により肝臓で代謝される。
www.ncbi.nlm.nih.gov/pubmed/22625877
CYP2C8は、スタチン製剤セリバスタチンの代謝に大きく関与し、フルバスタチンとシンバスタチン酸の代謝に控えめな役割を果たすと考えられている。
CYP2C8多型は、NSAIDの代謝に影響を与える可能性がある。
www.ncbi.nlm.nih.gov/pmc/articles/PMC2778050/
CYP2C9
薬の最大15から20パーセントの代謝クリアランスの原因であると推定されている。
CYP2C9によって代謝される薬剤
- イルベサルタン、
- ロサルタン アンジオテンシンIIブロッカー
- フェニトイン 抗てんかん薬
- シクロホスファミド アルキル化剤
- タモキシフェン 抗エストロゲン
- フルバスタチン スタチン
- セレコキシブ NSAID
- ジクロフェナク NSAID
- イブプロフェン NSAID
- ロルノキシカム NSAID
- メロキシカム NSAID
- ナプロキセン NSAID
- グリベンクラミド スルホニル尿素
- グリメピリド スルホニル尿素
- グリピジド スルホニル尿素
- トルブタミド スルホニル尿素
- ワルファリン 抗凝固剤
www.ncbi.nlm.nih.gov/pubmed/19152219/
www.ncbi.nlm.nih.gov/pmc/articles/PMC3201766/
CYP2C9はリファンピシンによって誘発される。
ワルファリン
誘導剤
葉酸
www.ncbi.nlm.nih.gov/pubmed/20206792
セントジョーンズワート
www.ncbi.nlm.nih.gov/pubmed/15100173
PXRの活性化による潜在的なCYP2C9誘導剤
- ハマヨモギ
- アルテミシニン
- ビャクジュツ(オオバナオケラ)
- ベルベリン塩酸
- カフェストール
- 中国アンジェリカ
- オウレンキネンシス
- クルクミン
- イカリソウ
- オイゲノール
- クチナシの果実
- イチョウ
- ググルステロン
- 川芎(Ligusticum chuanxiong)
- リグスチリド
- ルテオリン
- キョウチクトウ
- ミルクシスル(オオアザミ)
- ジャノヒゲ
- シャクヤクlactiflora
- ツルドクダミ
- アカヤジオウの根(地黄)
- レスベラトロール
- 朝鮮五味子
- シザンテリン A
- キカラスウリ
- ターメリック
阻害剤
レスベラトロール(強力)
www.ncbi.nlm.nih.gov/pmc/articles/PMC2933312/
ベルベリン
www.ncbi.nlm.nih.gov/pmc/articles/PMC4898966/
甘草
herbpedia.wdfiles.com/local–files/attachments/herbal_bioactivation_mini_review.pdf
CYP2C19
プロトンポンプ阻害薬、抗てんかん薬など、生体異物の代謝に関与する。
臨床で使用される薬物の10%に作用する肝臓酵素。
エポキシゲナーゼ活性があり、脂質代謝に関わる。
アルツハイマー病への変異体保護効果
CYP2C19遺伝子多型のアルツハイマー病患者では、アミロイドβの負荷が減少しており、変異体が保護効果を示す。
推定されるメカニズムは、CYP2C19がエストラジオールの触媒作用に関与しており、遺伝子多型によりエストロゲンの異化作用が低下することでエストロゲンの有益な効果がアミロイドβ産生におよぶとするもの。
もうひとつは、アラキドン酸の代謝物であるエポキシエイコサトリエン酸(EET)は、血流、皮質血管新生の調整、抗炎症反応などに有益な効果があることが示されており、CYP2C19変異がアラキドン酸の他の経路によって代謝されることで、EETの産生が減少し保護効果が失われるというもの。
rs4388808
www.ncbi.nlm.nih.gov/pmc/articles/PMC5820598/
阻害剤
レスベラトロール
www.ncbi.nlm.nih.gov/pubmed/20186406
セントジョーンズワート
www.ncbi.nlm.nih.gov/pubmed/19371257
誘導剤
アスピリン
www.ncbi.nlm.nih.gov/pubmed/12621391
イチョウ
www.ncbi.nlm.nih.gov/pubmed/19371257
セージ
www.ncbi.nlm.nih.gov/pubmed/19371257
セントジョーンズワート
www.ncbi.nlm.nih.gov/pubmed/19371257
バレリアンルート
www.ncbi.nlm.nih.gov/pubmed/19371257
CYP2D6
CYP2D6は、脳の様々な領域の神経細胞、グリア細胞で発現しており、神経保護酵素として考えられている。
ドーパミン合成
CYP2Dを介した脳内でのチラミンからのドーパミン合成がin vitro、in vivoで示されておりCYP2D6多型は、パーキンソン病と肺がんに関連している可能性がある。
ドーパミンやセロトニンバランスに関与することで、ヒトの性格や行動特性に影響を与える。
www.ncbi.nlm.nih.gov/pubmed/19604084
www.ncbi.nlm.nih.gov/pubmed/19374522
セロトニン合成
5-MTから5-HTへのO-脱メチル化は、肝臓/脳ミクロソームにおけるヒトおよびラットcDNA発現CYP2Dアイソフォームについてin vitroで報告されている。
www.ncbi.nlm.nih.gov/pubmed/23098818
パーキンソン病の保護CYP
CYP2D6は年齢とともに脳内で増加するが、パーキンソン病患者では低下する。
CYP2D6は農薬の代謝に関与しており、CYP2D6の遺伝的多型による酵素活性の低下がパーキンソン病発症リスクを高めることはよく知られている。
www.ncbi.nlm.nih.gov/pubmed/28871472
CYP2D6
レスベラトロール(高用量)
www.ncbi.nlm.nih.gov/pubmed/20716633/
ガーデンクレス(コショウソウ)
胡椒のような辛味があるアブラナ科野菜
www.ncbi.nlm.nih.gov/pubmed/24711855/
ケール
CYP2D6への抑制効果
www.ncbi.nlm.nih.gov/pubmed/22975634
CYP2E1
発がん物質の活性化
CYP2E1は、ハロタン、イソフルラン、クロルゾキサゾン、エタノールなどの神経系薬を代謝し、発がん性ニトロソアミンおよびアフラトキシンB1を生物活性させる。また基質に関係なくフリーラジカルを生成する。
www.ncbi.nlm.nih.gov/pubmed/12369887/
CYP2E1多型は冠動脈疾患、胃癌リスクと関連する。
www.ncbi.nlm.nih.gov/pubmed/20406102/
www.ncbi.nlm.nih.gov/pubmed/12115538/
CYP2E1によって誘発される酸化ストレスは、GLUT4発現の抑制を介してインスリン作用の障害を引き起こすことも示されている。
www.ncbi.nlm.nih.gov/pubmed/24500986/
パーキンソン病
CYP2E1は、MPTP毒性の増強と関連しており、パーキンソン病リスクを高める可能性がある。
www.ncbi.nlm.nih.gov/pubmed/17017526
パーキンソン病患者の脳ではCYP2E1のDNAメチル化が低下を示す。
www.ncbi.nlm.nih.gov/pubmed/22238121
誘導剤
中鎖トリグリセリド(MCTオイル)
www.ncbi.nlm.nih.gov/pubmed/17681033
ニコチン
www.ncbi.nlm.nih.gov/pmc/articles/PMC4314297/
絶食
www.ncbi.nlm.nih.gov/pmc/articles/PMC4297929/
オメガ3多価不飽和脂肪酸
アルコール
www.ncbi.nlm.nih.gov/pmc/articles/PMC4314297/
NAFLD
www.ncbi.nlm.nih.gov/pmc/articles/PMC4757513/
阻害剤
レスベラトロール
www.ncbi.nlm.nih.gov/pubmed/26680654
ケルセチン
www.ncbi.nlm.nih.gov/pubmed/23583009
www.ncbi.nlm.nih.gov/pubmed/25317811
クレソン
www.ncbi.nlm.nih.gov/pubmed/9728894
にんにく
www.ncbi.nlm.nih.gov/pubmed/22138249
www.ncbi.nlm.nih.gov/pubmed/12470597
www.hindawi.com/journals/jnme/2015/760689/
N-アセチルシステイン
エラグ酸
www.hindawi.com/journals/bmri/2013/358945/
緑茶・紅茶
www.ncbi.nlm.nih.gov/pubmed/24815822
クリシン
クルクミン
www.ncbi.nlm.nih.gov/pubmed/24063989
ピペリン
www.ncbi.nlm.nih.gov/pubmed/27670974
たんぽぽ抽出物
www.ncbi.nlm.nih.gov/pubmed/20812277
丹参
www.ncbi.nlm.nih.gov/pubmed/25645193
CYP3
CYP3Aアイソフォームの発現は組織特異的。
誘導剤
ルイボス茶、ニンニク、魚油
阻害剤
緑茶、紅茶、ケルセチン
CYP3A4
病院で処方される薬剤の50%以上を除去する重要なCYP酵素。
www.ncbi.nlm.nih.gov/pubmed/24312617/
カフェイン、テストステロン、プロゲステロン、アンドロステンジオンはCYP3A4酵素系の基質であり、多くの食品やサプリメントはこの酵素活性を低下させる。
このCYP3A4酵素活性は幅広く個人差があるため、高用量であったり複数の薬物投与、食品摂取、ハーブの相互作用はリスクをもたらしうる。
誘導剤
セントジョーンズワート
www.ncbi.nlm.nih.gov/pmc/articles/PMC1885170/
www.ncbi.nlm.nih.gov/pubmed/23778343
カプサイシン
www.ncbi.nlm.nih.gov/pubmed/25678312
www.ncbi.nlm.nih.gov/pubmed/22648626
ビタミンD
www.ncbi.nlm.nih.gov/pmc/articles/PMC3549031/
クルクミン
タバコ 多環芳香族炭化水素(PAH)
www.ncbi.nlm.nih.gov/pubmed/23845848
アフラトキシンB1
www.ncbi.nlm.nih.gov/pubmed/21641981
阻害剤
グレープフルーツジュース
www.ncbi.nlm.nih.gov/pubmed/15449971
www.ncbi.nlm.nih.gov/pubmed/15449971
レスベラトロール
ケール
www.ncbi.nlm.nih.gov/pubmed/22975634
ベルベリン
www.ncbi.nlm.nih.gov/pubmed/21870106
ケルセチン
www.ncbi.nlm.nih.gov/pubmed/25684704
ピペリン
www.ncbi.nlm.nih.gov/pmc/articles/PMC3208085/
www.ncbi.nlm.nih.gov/pubmed/16454693
アピゲニン
www.ncbi.nlm.nih.gov/pmc/articles/PMC4477257/
ミルクシスル
www.ncbi.nlm.nih.gov/pubmed/23674609
EGCG
www.ncbi.nlm.nih.gov/pubmed/19170155
コショウソウ
www.ncbi.nlm.nih.gov/pmc/articles/PMC3966352/
ブロッコリー(スルフォラファン)
www.ncbi.nlm.nih.gov/pubmed/17028159
ブラックコホシュ
オリーブオイル(オレウロペイン)
www.ncbi.nlm.nih.gov/pubmed/16454693
クランベリー
エキナセア
高麗人参(ジンセノサイドRd)
www.ncbi.nlm.nih.gov/pubmed/15133536
CYP3A5
その他のCYP450と同様、ほとんどが前立腺と肝臓で発現している。肝臓シトクロムの30%を担っている。遺伝的変異があり民族によって異なる。
www.ncbi.nlm.nih.gov/pmc/articles/PMC4164225/
誘導剤
セイヨウオトギリソウ
www.ncbi.nlm.nih.gov/pmc/articles/PMC4727115/
阻害剤
CYP4
CYP4ファミリーは、薬物代謝において役割を果たすと考えられているが、あまり詳細には解明されていない。
クロフィブラート、シプロ(脂質低下薬)、NSAIDは、プロスタグランジン、フタル酸エステルなどの毒物と関連する肝臓外のシトクロムファミリーであると理解されている。
www.ncbi.nlm.nih.gov/pubmed/9068972
CYP4A1
CYP4A1の遺伝的欠損は、20HETE形成の障害、血液脳関門の破壊、血管認知障害の発症に寄与する可能性がある。
www.ncbi.nlm.nih.gov/pubmed/25540098
誘導剤
緑茶抽出物
CYP4B1
CYP4B1アイソフォームは、MCTオイル(中鎖トリグリセリド)の代謝、および肺毒性および発がん性化合物の生物活性化に関与する。
www.ncbi.nlm.nih.gov/pubmed/16877261/
このサブグループの多型および過剰発現は、膀胱がん、大腸炎と関連している可能性がある。大腸炎とCYP4B1活性の関連性を調べ、カフェイン含有食品に見られるカフェイン酸によるCYP4B1活性の促進が大腸炎疾活動の低下、抗炎症作用と相関することを発見された。
www.ncbi.nlm.nih.gov/pubmed/19307459/
CYP7~27
内因性の物質の代謝はCYP7~27ファミリーが含まれ、ステロイドや、脂溶性ビタミンに対して高い基質特性を示す。
CYP7A1
コレステロール代謝に重要なCYP7A1遺伝子によってコードされる酵素。コレステロール7α-ヒドロキシラーゼとも呼ばれ、胆汁酸合成、コレステロールレベルの調節に関わる。(胆汁酸合成の律速段階)
胆汁酸のホメオスタシスは、核内受容体 ファルネソイドX受容体 / farnesoid X receptor (FXR)によって厳密に規制されている。
通常の条件下では、胆汁酸による核内受容体 ファルネソイドX受容体 (FXR)の活性化によって、CYP7A1の発現は抑制され胆汁酸産生は低下する。
コレステロールレベルが高い場合、CYP7A1は肝臓X受容体(LXR)によってアップレギュレートされ、胆汁酸産生を増加させ幹細胞コレステロールレベルを低下させようとする。
en.wikipedia.org/wiki/Cholesterol_7_alpha-hydroxylase
断食(PGC-1α活性)
www.ncbi.nlm.nih.gov/pmc/articles/PMC2397466/
ビタミンD
ビタミンD受容体の活性化はCYP7A1を増加させコレステロールを低下させる。
www.ncbi.nlm.nih.gov/pubmed/24365583
CYP8B1
主要な2つの胆汁酸ケノデオキシコール酸とコール酸のうちコール酸の合成にCYP8BAが必要となる。CYP8B1は、胆汁酸プールのケノデオキシコール酸とコール酸の比率を調節する。
www.ncbi.nlm.nih.gov/pmc/articles/PMC3676171/
CYP11A1
シトクロムP450(CYP11A1)はミトコンドリアの膜内の酵素であり、LXRアゴニストの形成を触媒できる。
www.ncbi.nlm.nih.gov/pubmed/21520051
CYP17A1
ステロイドホルモンの生合成ステップ
CYP17A1は、脳内のプレグネノロンをDHEAに変換する酵素。
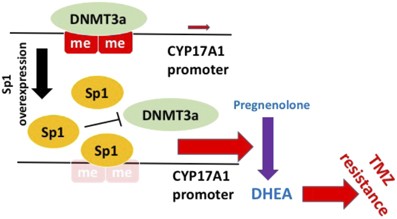
www.nature.com/articles/oncsis201731
rs743572
www.snpedia.com/index.php/Rs743572
www.ncbi.nlm.nih.gov/pubmed/20432167
www.ncbi.nlm.nih.gov/pubmed/22714708
CYP19A1
エストロゲンの形成(アロマターゼ酵素)
アンドロステンジオンは、17-ケトレダクターゼによってテストステロンに還元されるか、CYP19A1によってエストロンに芳香化される。
テストステロンまたはエストロンは、テストステロン還元(17-ケトレダクターゼ)またはエストロン芳香族化(CYP19A1)により、エストラジオール(17β-エストラジオール)に変換が可能。
エストロゲンと関連するCYP19A1遺伝子のrs3751592 A / G多型は中国漢民族のアルツハイマー病発症率と関連する。
www.ncbi.nlm.nih.gov/pmc/articles/PMC5008603/
誘導剤
フォルスコリン
www.ncbi.nlm.nih.gov/pubmed/14709151
CYP24A1
CYP24A1は、ヒト脳でのビタミンDの異化に関与している。CYP24A1の活性化は、不活性であるカルシトロン酸を形成する。
www.ncbi.nlm.nih.gov/pubmed/15544953/
www.ncbi.nlm.nih.gov/pmc/articles/PMC4553343/
en.wikipedia.org/wiki/Calcitroic_acid
CYP24A1には多くの多型が特定されているが、この多型が及ぼす影響については殆ど知られていない。高カルシウム血症を引き起こすことが予想されている。

www.ncbi.nlm.nih.gov/pmc/articles/PMC3927478/
CYP27A1
ヒトのCYP27A1遺伝子変異は脳腱黄色腫症(CTX)として知られる胆汁酸と関連する障害をもたらす。これらは胆汁酸の不足によるビタミンDの吸収不良の結果であると考えれている。
www.ncbi.nlm.nih.gov/pubmed/2019602/
紫外線に曝されたマウスは、ビタミンDの合成を高めCYP27B1の発現が4倍増加した。炎症サイトカインであるIL-1βを5倍、IL-17を減少させることから、紫外線による抗炎症効果が示唆された。
高用量のビタミンD投与でもIL-17の減少は認められたが、有意な差ではなかった。単独よりもビタミンD摂取と紫外線曝露の両方によるビタミンD合成によって、有益な効果を得ること示唆される。
www.ncbi.nlm.nih.gov/pmc/articles/PMC6116187/
CYP46A1
CYP51A1
CYP51は、ラノステロールからコレステロール合成への中間体を生成する。
家族性アルツハイマー病におけるコレステロールレベルの上昇の根底にはCYP51発現の増加が示されている。
www.ncbi.nlm.nih.gov/pubmed/19005074
CYP51の阻害剤(デブコナゾール)は、コレステロールレベルを低下させ、脂質ラフト関連APPの局在化を逆転させ大幅に低下することを示した。
journals.plos.org/plosone/article?id=10.1371/journal.pone.0210535