
Changes in functional connectivity and GABA levels with long-term motor learning
2015年8月01日
要約
新しい運動技能の学習は、一次運動野(M1)内の局所抑制回路を変化させ(Floyer-Lea et al 2006)長距離の機能的接続性を変化させる(Albert et al 2009)。このような効果が長期的なトレーニングで起こるかどうかは、あまり確立されていない。さらに、学習に関連した機能的接続性の変化と局所的な抑制との関係、および練習によるそれらの変調については、これまで検証されていない。
ここでは、安静時機能的磁気共鳴画像法(rs-fMRI)を用いて機能的接続性を評価し、MR分光法を用いて一次運動野(M1)のGABAを定量化した。参加者は1日30分(高強度群)または15分(低強度群)の練習を行った。我々は、異なるトレーニングレジームが脳の接続性と局所抑制の異なる変化に反映され、学習によって誘発されたGABAの変化と機能的接続性の間に相関関係が見られるのではないかという仮説を立てた。
両群とも練習によってパフォーマンスは有意に向上し、低強度群と高強度群の間でパフォーマンスの結果に差があることを示す証拠は見られなかった。低強度群では運動ネットワークの機能的接続性の増加とGABAの減少がみられたが、高強度群では機能的接続性の減少とGABAの有意な変化はみられなかった。機能的接続性の変化はパフォーマンスの結果と相関していた。機能的接続性の学習関連の変化はGABAの変化と相関していた。
この結果は、練習スケジュール間でパフォーマンスの結果が同等であっても、異なるトレーニングレジームが脳の変化の異なるパターンと関連していることを示唆している。さらに、安静時のネットワーク強度の学習関連の変化は、一部ではGABA作動性可塑性プロセスを反映していることが示された。
キーワード
可塑性、機能的接続性、GABA、運動学習
序論
数回のトレーニングセッションを経て獲得した要求の厳しい新しい運動技能の学習は、脳の構造的および機能的可塑性を誘導する(例:Dayan and Cohen, 2011; Doyon et al 2009; Sampaio-Baptista et al 2014; Scholz et al 2009)。実施される練習の量は、長期的な運動学習に関連する構造変化を調節し(Sampaio-Baptista et al 2014)タスクパフォーマンスのために募集された機能ネットワークに影響を与える(Doyon and Ungerleider 2002)。最近では、安静時fMRIを用いて、運動学習が脳の安静時機能連結性を変化させることが示されている。例えば、11分間の運動訓練を行うと、頭頂前野と小脳の安静時ネットワーク(RSN)の強度が増加した(Albert et al 2009)。別の研究では、運動RSNの強度は学習段階によって異なることが示唆されている(Ma er al)。 しかし、異なる練習スケジュールが脳の機能的接続性に与える影響については、これまで検証されなかった。また、これらの変化が神経生理学的メカニズムの観点から何を反映しているのかは明らかではない。
これまでの研究では、RSNはGABA作動性ネットワーク内の活動によって部分的に駆動される可能性が示唆されている(Kapogiannis et al 2013;Stagg et al 2014)。ある研究では、後頭葉皮質のGABA濃度とデフォルトモードネットワークの強さとの間に負の相関があることが示されている(Kapogiannis er al)。 これらの知見は、運動RSN内の接続性がM1内のβおよびガンマ周波数振動の変動に関連していることを示唆するシミュレーションおよびMEG研究から得られた収束した証拠と一致している。
学習に関連したシナプス変化の調節におけるGABAの役割は、動物モデルにおいてよく記述されている(Donato et al 2013;Frock et al 2000;TreppelおよびRaccine 2000)。同様に、ヒトにおける以前の分光学的研究では、感覚運動経験の短期的変化に応答してGABAレベルが減少することが示されている(Floyer-Lea et al 2006;Levy et al 2002)。しかし、ヒトにおける長期的な学習に伴うGABAの変調についての報告はない。
ここでは、ネットワークレベルの機能的接続性と局所的な抑制の学習関連の変化と、異なる練習スケジュールによるそれらの変調との関係を検証する。我々は、運動ネットワークの強さとGABAレベルが異なる量のジャグリング練習によってどのように変調されるかをテストするために、1日あたりのジャグリング訓練の量を操作した。
我々は、局所的な抑制性緊張の低下によって回路が強化された場合、対応するRSNの強度が増加し、局所的なGABAが減少するという仮説を立てた。例えば効率性の向上によって回路の強度が純減している場合は、RSNの強度が低下すると予測する。さらに、運動学習に反応してGABA濃度が変化すると、運動静止状態ネットワークの強度の変化と負の相関があるという仮説を立てている(Kapogiannis et al 2013; Stagg et al 2014)。
方法
参加者と実験デザイン
参加者64名(平均年齢23.8歳、標準偏差3.5,女性31名)が、現地の倫理委員会の承認(Oxfordshire REC B 07/Q1605/65)に従って、研究への参加にインフォームド・コンセントを得た。
このうち、ナイーブな参加者44人が、1日30分間ジャグリングを練習する高強度トレーニンググループと、1日15分間練習する低強度グループの2つのグループのいずれかに無作為に割り付けられた。低強度グループの4人の参加者は研究から脱落した(高強度グループのn=22,低強度グループのn=18)。両群とも週5日、6週間のトレーニングを行い、ベースライン時、6週間のトレーニング後(ポスト1)その後の4週間後(ポスト2)に再度スキャンを行い、参加者はジャグリングを行わなかった。同じ参加者で取得した他の画像モダリティの結果は、以前に報告されている(Sampaio-Baptista et al 2014)。
さらに、20人の参加者の対照群は、6週間間隔で2回スキャンされたが、ジャグリングトレーニングを受けていない。
FMRIデータは、高強度群20名、低強度群16名、対照群20名の参加者で取得した。MRSデータは、高強度群20名(うち2名はfMRIデータを取得していないことに注意)低強度トレーニング群16名、対照群19名で取得したが、比吸収率(SAR)の限界を超えていたため、MRSデータは取得しなかった。
行動評価
行動評価は別の場所に記載されている(Sampaio-Baptista et al 2014)。簡単に言うと、訓練グループの参加者は、最初の訓練日にグループレッスンを受け、そこでは最も単純なジャグリングパターンである「3ボールカスケード」が教えられた。その後、被験者は29日間、毎日自宅で練習した。参加者は、自宅でのトレーニングの様子をウェブカメラで撮影し、毎日、安全なウェブサイトにトレーニングのビデオをアップロードすることが義務付けられた。トレーニング期間終了前に3ボールカスケードをマスターした参加者には、3ボールリバースカスケードのようなより高度なジャグリングパターンの練習を奨励した。トレーニング期間終了後、参加者は4週間ジャグリングを中止した。
一日の最終的なスコアは、参加者一人あたり29本のトレーニングビデオのそれぞれについて、実験者が0~10の尺度で評価したものである(0:2ボール、1:3ボールカスケードの1サイクル、2:2サイクル、3:3サイクル、4:5~10秒の持続的な3ボールカスケード)。4: 持続的な3ボールカスケードの5~10秒;5: 10~20秒;6: 20~30秒;7: > 30秒;8: > 60秒;9: > 60秒および60秒未満の場合は少なくとも1つの他のパターン;10: > 60秒および60秒以上の場合は少なくとも1つの他のパターン)(Sampaio-Baptista er al)。 , 2014; Scholz er al)。) 各参加者の毎日のスコアに対数曲線をあてはめ、曲線の傾き(学習率)を算出した。
MRI
スキャンはオックスフォード大学で12チャンネルのヘッドコイルを備えた3テスラSiemens Trioスキャナーを用いて行った。全脳fMRIは、目を開けて安静時に勾配エコーEPIシーケンスを用いて行った(TR = 2000 ms、TE = 28 ms、フリップ角 = 89°、視野 = 224 mm、ボクセル次元 = 3 × 3 × 3.5 mm、取得時間 = 6分4秒)。FMRIデータは、高強度群20名、低強度群16名、対照群20名で取得した。
MPRAGEシーケンス(TR = 20.4 ms、TE = 4.7 ms、フリップ角 = 8°、ボクセルサイズ = 1 × 1 × 1 × 1 mm3)を用いて、1セッションにつき1枚の軸方向T1強調解剖学的画像を取得した。
運動手の表現における代謝物濃度は、局所的に取得されたスピンエコー全強度(SPECIAL; TR = 3000 ms; TE = 8.5 ms; フリップ角 = 90°; ボクセルサイズ = 20 × 20 × 20 × 20 mm3; 総スキャン時間 = 9 分 48 秒)を使用して評価した(Mekle et al 2009)。データは、左前頭前ノブの上に手動で配置された20×20×20mmのボクセルから取得した(Yousry et al 1997)。MRSデータは、高強度群の参加者20名(このうち2名はfMRIデータを取得していないことに注意)低強度トレーニング参加者16名、対照群の参加者19名で取得したが、比吸収率(SAR)の限界を超えていたため、MRSデータを取得した。
MRIによる解析
MRIデータの解析はFSLツール(www.fmrib.ox.ac.uk/fsl)を用いて行った。安静時fMRIは、Multivariate Exploratory Linear Optimized Decomposition into Independent Components (MELODIC) (Beckmann et al 2005)を用いて解析した。MELODICはデータ駆動法であり,他の成分とは独立した時間的に相関のある脳領域を含む成分を同定する.標準的な前処理には、頭の動きの補正、脳の抽出、6mmの半値幅(FWHM)で全幅のガウスカーネルを用いた空間平滑化、150秒(0.007Hz)に相当するハイパス時間フィルタリングが含まれている。FMRIボリュームは、境界ベースのレジストレーション(BBR)(Greve and Fischl, 2009)を用いて個人の構造スキャンに登録し、次にFMRIBの非線形画像レジストレーションツール(FNIRT)(Andersson et al 2007)を用いて標準空間に登録した。
被験者ごとに180の時間点を含む前処理された機能データを被験者間で時間的に連結し、単一の4Dデータセットを作成した。次に、全参加者と全スキャンを含むこの4DデータセットをMELODICへの入力として使用した。MELODICは、以前に記述された「正規の」安静時ネットワークをグループレベルで同定するために使用された(Beckmann and Smith, 2004)。
次に、各RSNの強さの個々の被験者尺度を計算するために、関心のある各グループレベルのRSNに二重回帰アプローチを適用した(Filippini et al 2009)。二重回帰の手順は以下の通りであった。
1. 関心のある各グループ-ICAに関連付けられた空間マップを、各グループレベルのコンポーネントに関連付けられた各被験者の時間コースを見つけるために、個々の被験者データに回帰させた。
2.これらの個々の被験者のタイムコースは、次に、ボクセル値が関心のある特定のグループICAとの関連の強さを表す被験者固有の空間マップを識別するためのリグレグレッサーとして使用された。
3. 関心のある各成分のグループ平均ICA空間マップを関心領域として使用し、各参加者のネットワークの平均値を計算するために、ステップ2で作成した個々の被験者マップに適用した。この値はネットワークの「強さ」に対応しており、値が高いほどネットワーク内の領域の相関性が高い(Stagg et al 2014)。
GABA濃度は、41個のシミュレートされた代謝物モデルスペクトルからなる基底セットを使用して、LCモデル(Provencher, 2001)を使用して自動的に計算された。すべての代謝物濃度は、総クレアチン(クレアチン+ホスホクレアチン)に対する比として与えられる。LCModel の適合の信頼性の指標である Cramer Rao Lower Bound(CRLB)が 15%を超える代謝物濃度は分析から除外した(対照群で 2 例(合計 n = 17)低強度群で 2 例(合計 n = 14)高強度群で 2 例(合計 n = 18))。
FMRIBの自動セグメンテーションツール(FAST)を使用して、高解像度T1強調解剖学的画像に基づいてボクセル内の灰白質(GM)および白質(WM)の相対量を計算した。GABA濃度は、ボクセル内のGM体積の割合[[[GM]/([GM]+[WM]+[脳脊髄液]で割った]]のために補正され、クレアチンは、ボクセル内の総脳組織体積の割合[[[GM]+[WM]で割った]/([GM]+[WM]+[脳脊髄液]で割った]のために補正された。(Stagg et al 2009)。
統計的分析
SPSSソフトウェア(バージョン21.00)を使用して、ジャグリングパフォーマンス、安静時ネットワーク強度、神経伝達物質濃度のデータを分析した。
統計的検定の前に、すべてのデータについてShapiro-Wilkで正規性を検定した。我々は、グループ間のジャグリング性能の経時変化(30日)について、Mixed-Design ANOVA(MD-ANOVA)を用いて検定した。真球性のMauchlyの検定が統計的に有意であった場合、Greenhouse-Geisser F-検定を使用し、それぞれの自由度を報告する。さらに、t-検定は、グループ間の学習率の違いを検定するために使用された。この実験からの行動結果は以前に報告されているが(Sampaio-Baptista et al 2014)ここでは、MRSまたは安静時fMRIを取得した特定の参加者の行動結果を報告していることに注意してほしい。
我々は、MD-ANOVAを使用して、グループ(高、低、コントロール)時間点(ベースラインとポスト1)とRSN(運動、デフォルトモード)と相互作用効果の間のRSN強度の違いについてテストした。ベースライン1とポスト1のタイムポイントだけがコントロールのために収集されたので、最初のANOVAに含めることができたことに注意してほしい。モーターRSNを関心のあるネットワークとみなし、デフォルトモードRSNをコントロールとして使用した(図1a,b)。有意な相互作用が発見された場合は、必要に応じて、ポストホック反復測定(RM)ANOVAまたはt検定を用いて追跡した。グループ間のポスト1 RSN強度の差は、ベースラインの差を考慮してANCOVAを用いて検定し、ベースラインの測定値を共変量とし、ポスト1 RSN強度を従属変数とした。
図1
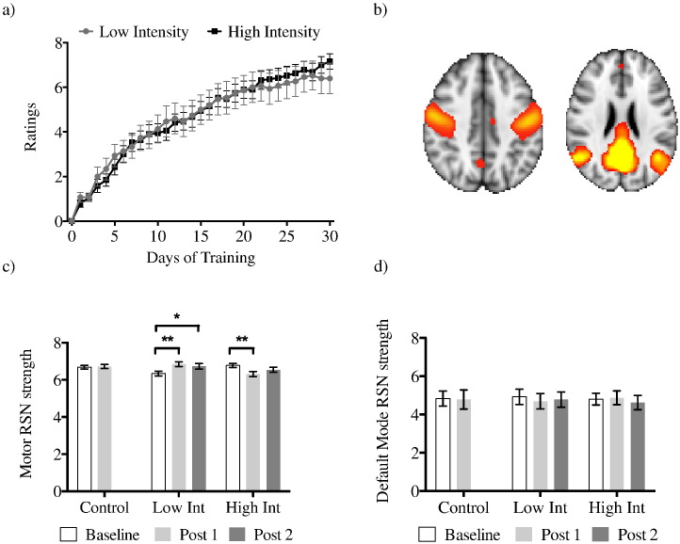
各グループの1日あたりの平均的なパフォーマンス評価を示した。
a) 各グループの1日あたりの平均パフォーマンス評価。日の有意な効果(F(3.933,129.795)=142.2, p = 0.00001)があるが、有意な相互作用効果(F(3.933,129.795)=0.999, p > 0.1)やグループ間の有意差(F(1,33)=0.005, p > 0.1)は見られない。デフォルトモードのネットワーク(右)。低強度群では、ベースラインとポスト1の間で有意な増加が見られ(**t(15)=3.283,p=0.005)、ベースラインとポスト2の間で増加傾向が見られた(*t(15)=2.347,p=0.033)。高強度群では、ベースラインとポスト1の間に運動RSN強度の有意な低下が見られた(*t(19)=2.787,p=0.012)。コントロール群では、ベースラインとポスト1の間に運動RSN強度に差はなかった(**t(19)=0.243, p > 0.1)。トレーニングはデフォルトモードRSNに有意な影響を与えなかった。棒グラフは標準誤差を示す。*補正なし, **ボンフェローニ補正に成功, p < 0.016.
トレーニング群内のポストホックt検定では、3つのタイムポイント(ベースライン、ポスト1,ポスト2)すべてを考慮した。適切な場合にはボンフェローニ補正を行った。
また、ベースラインのRSNを共変量として用い、RSN強度の変化とパフォーマンスの変化の間に部分的な相関があるかどうかを調べた。正規性はShapiro-Wilkで検定し、データが有意にノンパラメトリックな場合はSpearman検定を使用し、そうでない場合はPearsonのRを使用した(p < 0.05,2-tail)。我々は、Fisherのr to z(2-tail)を用いて、相関の強さの違いを検定した。
MD-ANOVAを用いて、群間(低、高、対照)および時点間(ベースラインおよびポスト1)の差、およびGABA濃度の相互作用効果について検定した。グループ間のポスト1のGABAの差は、ベースラインの差を考慮するためにANCOVAを用いて検定し、ベースラインの尺度を共変量として、ポスト1のGABAを従属変数として用いた。トレーニンググループ内のポストホックt検定については、すべての3つのタイムポイント(ベースライン、ポスト1,ポスト2)を考慮した。適切な場合はボンフェローニ補正を行った。
最後に、GABA変化が運動RSN強度変化と負の相関があるかどうかを検定した(p < 0.05,1-tail、この関係の方向性は先験的に予測されていたので(Kapogiannis et al 2013; Stagg et al 2014))。
結果
参加者全員が6週間のトレーニングで3ボールカスケードを3回連続で行うことができた。高強度グループの5人と低強度グループの4人は3ボールカスケードを完全にマスターし、逆3ボールカスケードなどのより高度なパターンを習得した。まず、ジャグリングを30日間続けて、低強度グループと高強度グループのパフォーマンススコアの違いを調査した。両グループの平均ジャグリング・パフォーマンスは時間の経過とともに向上したが(日の主効果(F(3.933,129.795)=142.2, p = 0.00001))グループ間の差(グループの主効果(F(1,33)=0.005, p > 0.1))や日とグループ間の交互作用(F(3.933,129.795)=0.999, p > 0.1)は見られなかった(図1a)。2つの訓練群では、学習率(傾き)に差はなかった(t(34)=0.758,p > 0.1)。まとめると、毎日の練習はジャグリングのパフォーマンスを向上させたが、1日あたりの練習量はパフォーマンスの結果には有意な影響を与えなかった。
続いて、練習による行動の改善が安静時脳活動の変化と関連しているかどうかを検証した(図1b)。長期学習は、ベースラインとポスト1スキャンの間で、ネットワークに特異的な方法で安静時脳活動を変化させた。MD-ANOVAにより、ネットワークの有意な主効果(F(1,53)=921.158,p = 0.00001)時間とグループ間の交互作用効果の傾向(F(2,53)=3.101,p = 0.053)グループ×時間×ネットワークの交互作用(F(2,53)=9.182,p = 0.00037)が明らかになった。これらの結果をさらに調査するために、各RSNについてポストホックMD-ANOVAを別々に行った。
トレーニングはデフォルト・モードRSNには有意な影響を与えなかったのに対し、運動RSNについては、時間とグループ間の有意な交互作用(F(2,53)=9.176,p = 0.000379)が見られたが、時間(F(1,53)=0.105,p > 0.1)やグループ(F(2,53)=0.880,p > 0.1)の有意な主効果は見られなかった(図1c,d)。
次いで、ベースラインの測定値を共変量として使用して、ベースラインの差を考慮するためにANCOVAを使用した。グループ間でポスト1 RSNの強さを比較すると、グループの有意な効果が見られた(F(2,53) = 5.154,p = 0.009)。この効果は、低強度群ではRSN強度が増加し、高強度群では減少することがポストホック検定で確認された(図1c)。コントロール群では、ベースラインとポスト1の間に運動RSN強度に差はなかった(t(19)=0.243,p > 0.1)(図1c)。
次に、ベースラインRSN強度との偏相関を共変量として用いて、トレーニングに関連した運動RSN強度の変化がパフォーマンスレベルと関連しているかどうかを検証した。その結果、高強度群では運動RSN強度の低下と学習率との間に有意な負の相関が認められ(Pearson r = -0.549,p = 0.01)(図2b)低強度群では運動RSN強度の上昇と学習率との間に正の相関が認められ(Pearson r = 0.513,p = 0.04)(図2a)これら2つの相関を直接比較すると、高強度群では運動RSN強度の低下と学習率との間に有意な負の相関が認められた。これら2つの相関関係を直接比較するためにフィッシャーの検定を用いたところ、2つの相関関係の差が有意であることがわかった(z = -3.21,p = 0.0013)RSN強度の変化とパフォーマンスの関係が練習群間で異なることが示唆された。
図2
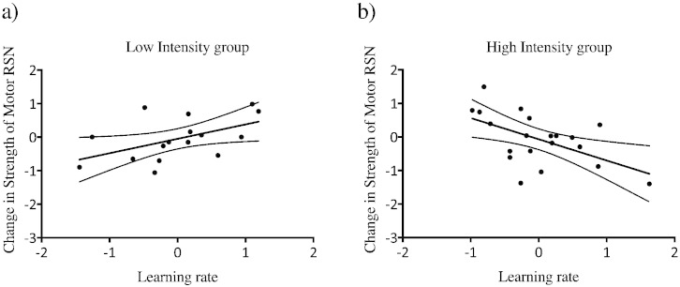
a) 低強度群では運動RSN強度の増加と学習率に正の相関がある(*p = 0.04) b) 高強度群では運動RSN強度の減少と学習率に有意な負の相関がある(**p = 0.01)。偏相関の残差をプロットした。2つの相関は互いに有意に異なる(z = – 3.21,p = 0.0013)。*未補正、**ボンフェローニ補正、p < 0.025。
次に、一次運動野内のGABAレベルに対するトレーニングの効果を調査した(図3a)。時間と群間の相互作用効果(F(2,46) = 4.655,p = 0.014)があったが、時間(F(1,46) = 2.583,p > 0.1)および群(F(1,46) = 0.597,p > 0.1)の主効果はなかった。ベースラインでの差を考慮するためにANCOVAを使用した。グループ間でポスト1 GABAを比較すると、グループの有意な効果が見つかった(F(2,46) = 3.942,p = 0.043)。ポストホックt検定では、これらの効果は、低強度群内での学習に伴うGABAの減少によって駆動されることが示された(図3b)。コントロール群では、ベースラインとポスト1の間に差はなかった(t(16)=0.695,p > 0.1)(図3b)。
図3
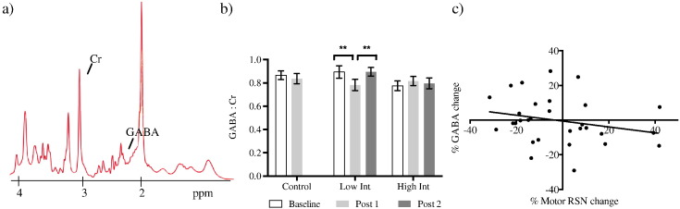
a)代表的なMRSスペクトル。 b)各群の時間を通してのGABA:クレアチン。低強度群のみベースラインとポスト1(**t(13)=3.899,p = 0.002)ポスト1とポスト2(**t(13)=3.075,p = 0.009)の間に有意な差がある。**ボンフェローニ補正p < 0.016。 c)GABA濃度の変化は、学習後の運動RSNの変化と負の相関がある(スピアマンr = – 0.326,p = 0.039)。エラーバーはSEMを表す。
すべてのジャグラーに渡って、GABAの変化は運動RSN強度の変化と負の相関があることがわかった(Spearman r = – 0.326, p = 0.039; 1-tail)GABAが減少すると運動RSN強度は増加した(図3c)。GABA変化と運動RSN変化との間には、各群に分けて有意な相関は認められなかった。
考察
本研究では、複雑な運動課題の長期学習が局所的な抑制緊張と安静時の脳の接続性に及ぼす変化を調べた。低強度の練習を行った被験者では運動ネットワークの接続性が増加し、GABAが減少したのに対し、高強度のトレーニングを行った被験者では運動RSN内の接続性が減少し、GABAには有意な変化は見られなかった。さらに、パフォーマンスの結果と運動RSN強度の変化には有意な関係があったが、その方向性はグループ間で異なる:高強度グループではパフォーマンスが向上した場合には減少が大きく、低強度グループではパフォーマンスが向上した場合には機能的接続性の増加が大きくなる傾向があった。
低強度群で観察された運動RSN強度の増加は、短期の運動学習を1セッションだけ行った後に見られた増加の報告を反映している(Albert et al 2009)。低強度群と高強度群の間で観察された違いは、長期的な指の配列学習に関する先行研究と一致しており、学習の最初の2週間でM1とS1構造間の機能的接続性が増加し、その後、2週目から4週目にかけてこれらの領域間の接続性が減少することが示された(Ma er al)。 今回の研究では、トレーニング終了後4週間後にフォローアップスキャンを行ったところ、低強度群で検出された運動RSN強度の増加がこの時点でも継続していることがわかった。
今回の研究や先行研究で観察されたパターンの一つの解釈として、低強度群では以前に確立された機能的接続の強度を高めることに依存し、高強度群では効率性を高めることになるということが考えられる(Penhune and Steele, 2011; Toni et al 1998)。これらの結果は、重複する参加者のサンプルにおける最近の構造的知見と興味深い類似性を示している:ジャグリングの練習量が異なると、低強度群では前運動野およびDLPFCのGM量が減少し(パフォーマンスと相関していた)高強度群ではGM量が増加した(これもパフォーマンスと相関していた)(Sampaio-Baptista et al 2014)。構造的測定と機能的測定の両方が、重複する領域(前運動野)だけでなく、異なる脳領域(DLPFC)でも、このトレーニング課題における練習とパフォーマンスとの関係を示しており、運動領域の構造的および機能的変化のための共通の根本的なドライバーがあることを示唆している(Sampaio-Baptista et al 2014)。先行研究では、構造的な結果に基づいて、異なる量の練習が異なる細胞メカニズムを引き出すという仮説を立てた(Sampaio-Baptista et al 2014)。今回の研究では、練習量が低ければ(GM量の減少によって証明されるように)剪定が誘発され、その強度を増加させて機能的接続性を増加させることによって、以前に確立された機能的接続の大部分に依存する可能性があり、一方、練習量が多ければ新しい接続の形成(およびGM量の増加)が誘発され、回路効率の増加につながる可能性があり、機能的接続性の減少によって反映されるという更なる証拠を提供している。
以前に行われたバランシング課題を用いた研究では、参加者が週1回6週間練習した後のパフォーマンスと左内側頭頂皮質の安静時fMRI信号の変化に相関があることが報告されている(Taubert et al 2011)。これら2つの研究は、トレーニング課題や強度、安静時のfMRI測定法が異なる。例えば、Taubertらが行った全身バランス課題では、プロプリオレセプティブな処理が行われている可能性が高いため、内側頭頂皮質に効果が見られたのかもしれない。この研究で使用された機能的接続性の測定方法が異なるため、我々の結果と直接関連付けることは難しいが、どちらの研究も、長期トレーニングによって安静時の脳の接続性が構造の変化に関連する方法で変化するという考えを支持するものである(Sampaio-Baptista et al 2014; Taubert et al 2011)。
今回の研究で発見された解離についての別の考え方としては、両グループとも6週間の練習を行っているにもかかわらず、毎日の練習の強度が異なるということは、両グループの個人が実質的に学習の異なる段階にあることを意味している(Dayan and Cohen, 2011; Penhune and Steele, 2011)。しかし、この可能性を直接検証するためには、より多くのタイムポイントを用いて、6週間のトレーニングプロトコルの間に異なる段階での学習と脳の変化を問う研究が必要である。1つの予測としては、3週間後の30分群は6週間後の15分群と似たような(GABAと安静時のfMRI測定の観点から)結果が得られると考えられる。今後の研究では、より多くのスキャンタイムポイントを用いて、これらの変化の経時的な変化を調べ、トレーニングによってどれだけ早く変化が生じるかを評価することができるだろう。
異なる練習スケジュールのグループ間で有意なパフォーマンスの差がないことは注目すべきことである。しかし、我々の過去の研究では、今回の研究で使用した高強度プロトコルと同じ固定トレーニングプロトコルを使用した場合、最終的なパフォーマンスの結果には個人差が非常に大きいことが明らかになった。したがって、ここで採用されている異なる練習スケジュールの間でパフォーマンス成果の分布が重複していることがわかったのは、おそらく驚くに値しない。しかし、ジャグリングのスピードやより重要な動きの質を評価していないため、私たちの行動測定が粗雑すぎて微妙な違いを検出できない可能性もある。
私たちは、RSNの主要な分析を運動RSN、私たちの関心のあるRSN、およびデフォルトモードRSNに限定した。しかし、最近の他の報告では、トレーニングによる他のネットワークの変化が検出されているので(Albert et al 2009)すべてのネットワークについて上記と同様のアプローチで探索的分析を行ったが(データは示されていない)他のどのネットワークにも影響は見られなかった。
トレーニング後に低強度群で観察されたGABAの減少は、センサ運動経験の短期的な変化に反応してGABAレベルが減少することを示したヒトの過去のMRS研究と一致している(Floyer-Lea et al 2006;Levy et al 2002)が、長期学習後にこの効果を記述したのは今回の研究が初めてである。
ここで示されたGABAの所見は、示された機能的接続性の変化の神経生理学的根拠を提供するものである。運動RSN強度の変化と学習によるM1 GABA濃度の変化との間には、有意な(強さは控えめではあるが)相関関係があることがわかった。この知見は、運動学習に伴う安静時ネットワークの変化の根底にあると考えられる細胞メカニズムへの暫定的な洞察を提供し、(学習がない場合の)運動RSN内の活動が、GABA活性に関連することが知られているβおよびガンマ振動のパワーの変動に関連していることを示した先行研究(Brookes et al 2011;Cabral et al 2011)を拡張したものである(Hall et al 2011)。このように、我々の結果は、安静時のネットワーク強度は主要ネットワークノード内の振動活動の局所的なGABA作動性変調によって駆動される可能性があることを示唆する以前の研究(Kapogiannis et al 2013; Stagg et al 2014)を支持する。本研究では、RSN強度の変化が長期の運動訓練に応じたGABA作動性活動の変化と関連している可能性を示唆することで、これらの知見を追加している。
興味深いことに、低強度群のGABAレベルは、トレーニング終了後4週間のフォローアップスキャンではベースラインに戻っていた。この観察は、GABAの低下は主に学習が継続している間に見られるという考え方と一致している(Floyer-Lea et al 2006)。しかし、低強度群で観察されたGABA測定値のベースラインへの復帰は、この群で観察された機能的接続性に対する持続的な効果とは対照的であった。これらの差動効果は、これら2つの指標は学習が進行している間は関連しているが、長期的には異なることを示唆している。
これらの知見は、運動技能の長期的な獲得の基礎となるプロセスに光を当てるものであり、重要なことは、安静時fMRIが、脳卒中後の学習中に生じる神経生理学的変化や、ニューロリハビリテーションのような運動学習の一種であると解釈できる長期的な可塑性の例を調べるための感度の高いツールである可能性を示唆していることである。