
Contents
軸索原形質輸送(認知症・アルツハイマー病)
概要
軸索輸送とは
軸索輸送とは、神経細胞の軸索の中で様々なものを運ぶ機能のこと。
微小管がレールの役割を果たし、キネシンやダイニンなどの分子モーターによって物質が運ばれる。

神経細胞における役割
軸索輸送によって、神経細胞の軸索および神経末端にタンパク質、脂質、ミトコンドリアを供給し、ミスフォールドタンパク質を除去またはリサイクルして毒性凝集物の蓄積を防ぐ役割をもつ。
また、ニューロンの栄養シグナルやストレス障害への反応をにも関与し、細胞内の神経伝達物質に対して重要な役割を果たす。
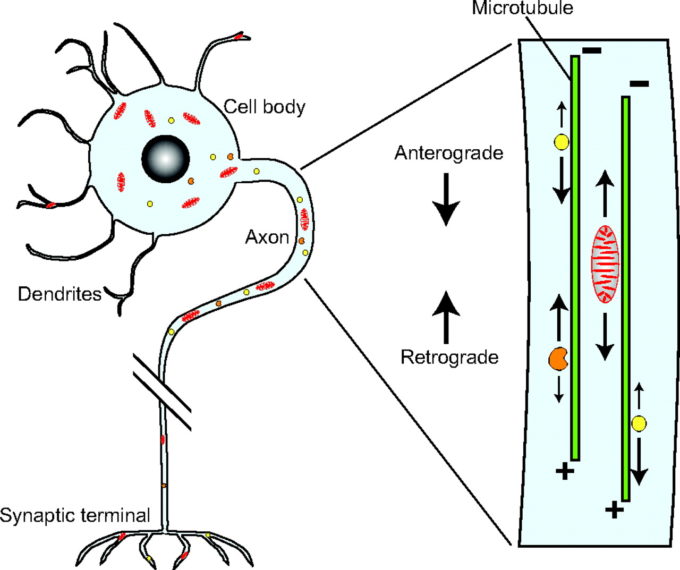
用語
順行性軸索輸送
移動方向:細胞体 → 軸索末端
モータータンパク質:キネシン(4nmのモーターヘッド、8nmのステップ長)
キネシンはアミロイド前駆体タンパク質(APP)として同定されている。
www.ncbi.nlm.nih.gov/pmc/articles/PMC1621108/
逆行性軸索輸送
移動方向:軸索末端 → 細胞体
モータータンパク質:ダイニン(10nmのモーターヘッド、8~32nmのステップ長)
逆行性輸送はダイニンだけでなくキネシンが加わった両者によって行われることがある。

www.cell.com/trends/biochemical-sciences/fulltext/S0968-0004(15)00143-7
速い軸索輸送
移動速度 50-400mm/日
貨物:ペプチド、糖脂質、小胞細胞
速い逆行性軸索輸送は10~20cm/日に到達することがあり、シナプス小胞などを利用して軸索末端の状態を体細胞に知らせる。
キネシン一歩間の距離を80cmとするなら、一日1~2万kmの移動に相当する!
中間体軸索輸送
移動速度 15~50mm/日
貨物:ミトコンドリア
遅い軸索輸送
低速輸送にはキネシンが関与する。
移動速度 8mm/日未満
貨物:チューブリン、アクチン、ニューロフィラメント
アルツハイマー病における軸索輸送障害
軸索輸送障害は原因なのか結果なのか
アルツハイマー病を含め多くの神経変性疾患は、軸索輸送の突然変異と関連していることが浮上している。
軸索輸送障害はアルツハイマー病病理が生じさせた結果であって原因ではない、と一般的に想定される傾向にある。
しかし、軸索輸送の障害が起因となって毒性タンパク質の蓄積に寄与する経路が複数存在し、軸索輸送障害によってアルツハイマー病が発症すると考えることが可能な証拠も十分に存在する。
軸索輸送障害が疾患の原因であるのか結果であるのかは現在議論の対象となっているが、軸索輸送の障害が生じるには複数の経路が存在しおそらく双方向的な関係にあるものと考えられている。
軸索輸送障害がアルツハイマー病の発症原因となる理由
・アルツハイマー病の初期段階で軸索輸送障害が関与している。
・軸索輸送の閉塞、減少は、シナプス小胞前駆体の枯渇および軸索障害となる物質の蓄積により、エネルギー産生の減少、シナプス機能不全、核周囲変性を引き起こす。
・軸索輸送障害によってアミロイドβ形成が引き起される。
・タウの過剰リン酸化および凝集は、軸索輸送障害によって誘発されるミトコンドリア呼吸鎖複合体Iの阻害から生じることができる。
軸索輸送を支える4つの基本成分と推定機序
細胞骨格(微小管)
過剰リン酸化タウ、アミロイドβオリゴマー → 微小管の障害 → 軸索輸送障害
モーター分子
ミトコンドリア障害 → Ca2+脱調節 → モーター分子の機能不全 → 軸索輸送障害
ATP(エネルギー)
ミトコンドリア障害 → ROS → エネルギー低下 → 軸索輸送障害
アダプタータンパク質
(貨物とモーターの結合に関わるタンパク質)
リン酸化タウ → アダプタータンパク質の結合能低下 → 軸索輸送障害

www.sciencedirect.com/science/article/pii/S0968000415001437
相乗的に生じる軸索輸送障害
アルツハイマー病病理においては、上記4つの軸索輸送基本成分が、それぞれ異なるシグナル伝達経路によってかき乱され軸索輸送障害を引き起こす。
これらの要因は重複しており、相乗的、循環的に軸索輸送障害に作用する。
content.iospress.com/articles/journal-of-alzheimers-disease/jad14108
軸索輸送障害因子
アルツハイマー病関連因子
病理学的な形態のAPP、タウ、PS1、アミロイドβは速い軸索輸送に影響をおよぼすことが示されている。
APP(アミロイド前駆体タンパク質)
APP過剰発現マウスにおけるNGF逆行性輸送の減少。エンドソームの膨張に起因する可能性がある。
www.ncbi.nlm.nih.gov/pubmed/16815330
PSEN1(プレセニリン-1)
プレセニリン1の変異は、GSK3βの活性増加と同時にKLCリン酸化レベルが増加、膜結合オルガネラ(MBO)に結合するキネシンIの量が減少、速い軸索輸送の欠損と一致する。
www.ncbi.nlm.nih.gov/pubmed/12805290
PS1 → GSK3β活性 → KLCリン酸化 →キネシン-1の貨物放出
MAPT(微小管関連膜タンパク質タウ)
タウの過剰リン酸化が微小管結合を低下させ、細胞骨格の完全性を失わせる。
微小管の不安定化は軸索輸送障害を引き起こす可能性がある。
www.nature.com/articles/4001483
www.ncbi.nlm.nih.gov/pubmed/18798283
www.ncbi.nlm.nih.gov/pmc/articles/PMC126299/
アミロイドβ
アミロイドβは、アクチン重合、凝集の促進を介して小胞の軸索輸送を、段階的かつ不可逆的に損なう。
www.ncbi.nlm.nih.gov/pubmed/14523099
アミロイドβは、NMDA受容体機構を介してミトコンドリアの軸索輸送を阻害する。
これにはGSK3β活性化を伴うが、NMDA受容体アンタゴニストまたはプロテインキナーゼ(PKA)-サイクリックAMP(cAMP)経路の刺激によって軽減されうる。
www.ncbi.nlm.nih.gov/pubmed/17035532
www.ncbi.nlm.nih.gov/pubmed/20610750
可溶性アミロイドβオリゴマー
軸索への可溶性オリゴマーアミロイドβの灌流は、内因性カゼインキナーゼ2の活性化およびKLCsのさらなるリン酸化により、小胞の双方向の輸送を阻害する。
アミロイドβ三量体がアルツハイマー病において軸索輸送障害を引き起こす可能性。
www.ncbi.nlm.nih.gov/pubmed/27629715
その他の軸索輸送障害因子・機序
ミトコンドリアの輸送障害
ミトコンドリアは神経細胞の細胞内で合成され、その生理学的機能を果たすために神経末端に運ばれる必要がある。
ミトコンドリアの輸送にはキネシン-1、KIF1Bαが主に関わる。
ミトコンドリアの軸索輸送はミトコンドリア機能不全などアルツハイマー病病変の影響を受けやすいと考えられている。
www.ncbi.nlm.nih.gov/pubmed/23061885
www.ncbi.nlm.nih.gov/pubmed/19605646
www.ncbi.nlm.nih.gov/pubmed/17166847
リソソームタンパク質の分解
ニューロンのリソソームタンパク質分解が、アルツハイマー病における軸索輸送障害の根底に存在し、ALS、前頭側頭型認知症など他の神経変性疾患においても重要な役割を果たしている。
www.nature.com/articles/nrn3380#
www.ncbi.nlm.nih.gov/pubmed/21613495
KLC1の枯渇
KLC1の枯渇による順行性軸索輸送の障害は、APP逆行性軸索輸送の増大およびアミロイドβの産生を促進する。
www.ncbi.nlm.nih.gov/pubmed/15731448
オートファジー
オートファゴソームの逆行軸索輸送障害が、アルツハイマー病病理のニューロンにおいてオートファジーストレスを増加させる。
www.ncbi.nlm.nih.gov/pmc/articles/PMC5235353/
細胞骨格組織の欠損
微小管へのモータータンパク質の付着による障害
キナーゼ活性の変化
モーターと貨物の結合の不安定化
biomedcircus.com/paper_03_42.html
RNA代謝と軸索輸送の相互作用
www.ncbi.nlm.nih.gov/pmc/articles/PMC5026263/
www.ncbi.nlm.nih.gov/pubmed/25511265
アルミニウム塩
www.sciencedirect.com/science/article/pii/0006899385913691
link.springer.com/article/10.1007/BF02815165
メチル水銀
www.ncbi.nlm.nih.gov/pubmed/4035699
水銀とEDTAの錯体
www.ncbi.nlm.nih.gov/pubmed/15364276
二硫化炭素などの工業溶媒
www.sciencedirect.com/science/article/pii/0006899387914715
感染症
単純ヘルペスウイルスによる順行性軸索輸送を利用したアルツハイマー病への寄与。
www.ncbi.nlm.nih.gov/pmc/articles/PMC3069030/
www.ncbi.nlm.nih.gov/pubmed/14677633
www.ncbi.nlm.nih.gov/pmc/articles/PMC5457904/
キネシン、ダイニンの遺伝子変異
www.ncbi.nlm.nih.gov/pmc/articles/PMC4269290/
コルヒチン
www.ncbi.nlm.nih.gov/pubmed/1731931
www.ncbi.nlm.nih.gov/pmc/articles/PMC1218294/
シナプス障害
シナプスの酸化障害
NMDAグルタミン酸受容体の酸化ストレス、
一酸化窒素NOの神経毒性によりシナプス変化を引き起こす。
NMDA受容体媒介のアポトーシスによるニューロン細胞死
シトクロムcの放出、カスパーゼ9の活性化、カスパーゼ3の活性によるミトコンドリア機能不全。
ミトコンドリア異常
ROS産生の増加、エネルギー貯蔵の低下、エネルギー代謝障害をもたらすシトクロムcオキシダーゼの欠乏、ミトコンドリア複合体の阻害
酸化ストレスによるタウのリン酸化
酸化ストレスによるPP1、PP2A阻害→ERK1/2活性の増強→tauの過剰リン酸化
www.ncbi.nlm.nih.gov/pmc/articles/PMC4470891/
一酸化窒素
一酸化窒素が、早期疾患段階ではシナプスの伝達および可塑性を高める。(神経保護と神経変性の役割)疾患が進行すると適切な機能として働かないかもしれない。
www.ncbi.nlm.nih.gov/pubmed/25926464
軸索再生阻害因子
Nogo-A(NgR)
MAG
RhoA
OMgp
Ephrin-B3
グリア瘢痕
炎症反応
cAMPの減少
bsd.neuroinf.jp/wiki/%E8%BB%B8%E7%B4%A2%E5%86%8D%E7%94%9F
標的候補・薬剤
HDAC阻害剤
HDAC6阻害剤
journals.plos.org/plosone/article?id=10.1371/journal.pone.0042983
www.ncbi.nlm.nih.gov/pubmed/22937007
ホスファチジルセリン
ホスファチジルセリンはHDACを阻害し、アセチル化チューブリンレベルを増加させ軸索の安定と成長を促進する。
www.ncbi.nlm.nih.gov/pmc/articles/PMC5436341/
微小管安定化剤
キサントフィル(ルテインなど)
www.ncbi.nlm.nih.gov/pubmed/11504633/
エポチロンD(EpoD)
www.biochemsoctrans.org/content/40/4/661
ダブネチド NAPVSIPQ(NAP)
www.nature.com/articles/mp201332
タキサン(パクリタキセル)
www.ncbi.nlm.nih.gov/pubmed/10074470/
その他
ディスコデルモライド、ディクチョスタチン、エリューテロビン、サルコドクチン、エレセラソイド、ラウリマライド、ペロルサイドA、シクロストレプチン、タカロニロリド、ザンパノライド、ダクチローリド、セラタミン、ジクマロール、ジャトロフェーンズ、ツベルシジン、フェニルピリミジン、ピリダジン、
www.ncbi.nlm.nih.gov/pmc/articles/PMC3493881/
Notchシグナル伝達標的
journals.sagepub.com/doi/10.1177/1756285613490051
ホスホトランスフェラーゼ活性
ROSの抑制
www.ncbi.nlm.nih.gov/pmc/articles/PMC5562667/
フルルビプロフェン
R-フルルビプロフェン(プロパン酸系の非ステロイド性抗炎症薬)がADマウスの軸索輸送を改善する。
www.ncbi.nlm.nih.gov/pubmed/21500269
ナノ抗酸化物質
ナノ抗酸化物質が、ADマウスの軸索輸送障害を改善する。(PEG-HCC?)
Ca2+の制御
jcs.biologists.org/content/125/9/2095
ATP産生
脂肪酸
脂肪酸(酢酸塩(C2)、カプロン酸塩(C6)、カプリル酸塩(C8)、カプリン酸塩(C10)、2-デセン酸塩(C10:1)、アラキドン酸塩(C20:4)の投与は、試験管モデルにおいて順行性軸索輸送、逆行性軸索輸送の両方で輸送される粒子の数が上昇する。
www.ncbi.nlm.nih.gov/pubmed/12711076