
Contents
Are environmental pollution and biodiversity levels associated to the spread and mortality of COVID-19? A four-month global analysis
オンラインで公開2020年12月21日
www.ncbi.nlm.nih.gov/pmc/articles/PMC7752029/
Daniel Fernández,a,∗ Iago Giné-Vázquez,b,c Ivy Liu,d Recai Yucel,e Marta Nai Ruscone,f Marianthi Morena,g Víctor Gerardo García,h,m Josep Maria Haro,b,c,i William Pan,j,k and Stefanos Tyrovolasb,c,l.
要旨
2020年3月12日、WHOはCOVID-19をパンデミックと宣言した。COVID-19の蔓延と死亡率の進化に及ぼす環境要因や生態系要因、さらには生物多様性の集団的影響は、特に生態系の範囲が広い地域では経験的に不明のままである。本研究の目的は、これらの要因がCOVID-19の蔓延と死亡率にどのように影響しているかを国別に評価することである。本研究では 2020年1月21日から5月18日までのWHOの日次症例報告と他の公的に入手可能な指標をマージした世界的なデータベースを作成した。生物多様性、気温、降水量の影響を特定するために時空間モデルを適用し、環境変数の影響を特定するために一般化線形混合モデルを適用した。さらに、COVID-19の拡散と大気質要因との関連を特徴づけるために、カウント時系列を用いた。すべての解析は、世界160カ国の社会人口統計学、国別所得水準、政府政策介入の交絡因子で調整された。その結果、COVID-19感染と国や都市レベルでの生物多様性指数、大気質、汚染要素(PM10,PM2.5,O3)などのいくつかの要因との間に統計的に意味のある関連があることが明らかになった。特に、生物多様性の喪失、高レベルの大気汚染物質、大気質の低下は、COVID-19感染の拡大と死亡率との間に有意な関係があることが明らかになった。本研究で得られた知見は、COVID-19の感染拡大と死亡率との関係について、今後の研究のための実証的な基盤を提供するものである。本研究で測定された関係は、政府がCOVID-19感染症の新たな発生に対応し、将来の危機を予防するための代替戦略として、環境・健康政策を立案する際に有用であると考えられる。
キーワード COVID-19,世界的、死亡率、感染、生物多様性、大気の質
グラフィカル・アブストラクト
1. はじめに
1918年のインフルエンザの大パンデミック(Taubenberger and Morens, 2006)2014年のエボラ(Rewar and Mirdha, 2014)ウイルス病、コウモリの白鼻症(Blehert er al 2013)トドマツの木の真菌症、世界的に両生類を死滅させたパンデミック・キトリ菌症(フィッシャー et al 2009;フィッシャー et al 2012)などが、頻度を増して恐ろしい結末を迎えている。パンデミックイベントやパンデミック性疾患の主な原因の一つは、人間の個体群と病原体を運ぶ家畜や野生動物との密接な相互作用です(Woolhouse and Gowtage-Sequeria, 2005)。ほとんどの病原体は、狩猟、野生種の消費、野生動物の取引、その他の野生動物との接触を通じて、野生動物の貯蔵庫から人間の集団に伝播する。さらに、地球の気候や天候の変化は、地球の生態系に影響を与え続けており、その中には感染性疾患の病原体やベクターとして作用する宿主を持つ環境コミュニティが含まれている(Yeh K et al 2020)。したがって、感染性病原体の出現の激化は、気候変動、生物多様性の喪失、生息地の劣化、人間と野生生物の相互作用(HWI)の速度増加にも起因すると考えられる(Schmeller et al 2020)。
生物多様性の豊かさと均等性の減少は、人間、動物、植物への感染症の伝播の緩衝材としての役割を果たす生態系の重要な部分の消失を引き起こす(Peixoto and Abramson, 2006; Pongsiri er al)。 その意味で、生物多様性の喪失に伴って病気の伝播が増加することがいくつかの研究で示唆されている(Keesing et al 2010; Wood et al 2014; Lacroix et al 2014; Johnson et al 2015)。さらに、生物多様性条約(CBD)が発表した国連の生物多様性の展望報告書(GBO-5)の第 5 版では、気候変動や長期的な食料安全保障に対処する際の生物多様性の重要性が指摘されている。GBO-5 は、将来のパンデミックを防ぐためには、生物多様性を保護するための行動が不可欠であると結論づけている。さらに、報告書は、生物多様性の損失は、感染症の出現と再出現の速度を速めることにもつながる可能性があると指摘している。
21世紀には、SARS、豚インフルエンザ、MERS、エボラ、ジカ、鳥インフルエンザの6つの大規模な疫病が発生している。ここでは、その特徴を簡単に紹介する。
- 2003年に発生した重症急性呼吸器症候群(SARS)は、8000人以上の感染者をもたらし、死亡率は約10%で、その影響は地域経済や地域経済に限定されていた(LeDuc and Barry, 2004)。このパンデミックは2003年7月に突然終了したが、それ以降、SARSコロナウイルスのヒト感染例は検出されていない。
- 豚インフルエンザの原因となる2009年のH1N1インフルエンザウイルスは 2009年3月にメキシコで初めて発生し、その後4月に米国で大パンデミックした。2009年の豚インフルエンザは、世界的な移動や航空旅行の結果としてパンデミックとなり、推定症例死亡率は0.4%に達した(Al Hajjar and McIntosh, 2010)。
- 中東呼吸器症候群(MERS)は 2012年にサウジアラビアおよびヨルダンでヒトで初めて確認された(Memish et al 2020)。MERSは、感染したドロメダリーラクダからヒトに飛び移る人獣共通感染性病原体と考えられている(El-Kafrawy et al 2019;Gardner et al 2019)。出現後1年以内に封じ込められたSARSとは対照的に、MERSは中東地域で限られた循環を続けており、断続的な散発性のヒト感染例、感染コミュニティのクラスター、および院内発生を引き起こし、これらはすべて世界的に広がるリスクが高い(Zumla et al 2015)。
- エボラウイルスは1976年にザイール(現在のコンゴ民主共和国)で初めて検出された。ウイルスが最初に検出されて以来、サハラ以南のアフリカ、主にスーダン、ウガンダ、コンゴ民主共和国、およびガボンで20以上のエボラの既知のアウトブレイクが確認されている(Malvy et al 2019)。現在のところ、エボラに対するワクチンまたは効率的な抗ウイルス管理戦略は存在しない(Hasan et al 2019)。エボラウイルスは、約28,000人の確定症例および11,000人の死亡者を有する2013-2016年西アフリカエボラウイルスパンデミックによって実証されたように、国際的な旅行の容易さのために、実質的な伝染病およびパンデミックの可能性を有しているが(Garske et al 2017年)エボラウイルスの感染は、エボラウイルスの感染を防ぐためには、エボラウイルスの感染を防ぐことが重要であると考えられる。
- ジカ熱(2015-2016)は、1947年にウガンダのジカの森で発熱したアカゲザルから初めて分離された。ヒトでの最初の症例が報告された1954年以降、ジカウイルスはアフリカとアジアで限られた散発的な感染を引き起こしたに過ぎなかった。しかし 2015年にはブラジルからアメリカ大陸の29カ国に約44万~130万人が感染した大規模なアウトブレイクが発生した(Plourde and Bloch, 2016)。2016年11月、WHOはジカの大発生の終息を発表した。
- 鳥インフルエンザ(または鳥インフルエンザ)は1997年に香港で初めて報告され、感染者数はわずか18人、死亡者数は6人であった。しかし、鳥インフルエンザの700例以上が60カ国以上から報告されており(Alexander and Brown, 2009)その中には中国で発生した2016年のアウトブレイクも含まれている(Chatziprodromidou er al)。
それらのパンデミックのいくつかは、その社会的および環境的影響を予測するために研究されてきた(Crameri et al 2015;Kim et al 2018;Qiu et al 2018)。現在の新規コロナウイルス疾患(COVID-19)は、空間的範囲および社会的影響の点で、21世紀のそれらの6つの大規模な伝染病を矮小化しており(Ali and Alharbi, 2020)COVID-19は、広範で複雑な環境影響を有する唯一のパンデミックである(Lal et al 2020; Nakada and Urban, 2020)。
2020年3月12日、世界保健機関(WHO)はCOVID-19をパンデミックと宣言した(WHO, 2020)。2020年5月18日までに、210カ国以上でCOVID-19の確定症例が報告された(Cucinotta and Vanelli, 2020)。中国国外へのウイルス感染の拡大は、様々な欧州連合(EU)諸国、アメリカ合衆国(米国)ラテンアメリカ諸国(ブラジルやペルーなど)アフリカ諸国(南アフリカなど)で報告された(Davis, 2020)。
COVID-19の憂慮すべき広がりを介入させ、封じ込めるために、いくつかの社会的遠距離措置が実施された(Pan et al 2020)。ワクチンや薬物治療法を開発するための様々な臨床試験が、ウイルスとの戦いに向けられている。ウイルスの拡散および死亡率に対する環境因子(すなわち、温度および湿度)の影響に関するいくつかの初期データが提示されている(Prata et al 2020;TobíasおよびMolina 2020;Triplett 2020;Wang et al 2020a;Wang et al 2020b)。しかし、これらの研究では、社会的距離と政府の移動制限政策の相互影響などの要因を交絡因子として含んでいるものはなかった。さらに、COVID-19の拡散における生物多様性の役割については、様々な研究者が指摘しているが(Corlett et al 2020;Grandcolas and Justine 2020;Lorentzen et al 2020;Outlook et al 2010年)現在まで、地域レベルや国際レベルでのその役割に関する情報はない。生態系フォーラムではまだ議論が続いているが、理論的には、生物多様性の向上は、未知のウイルスに対する人間の免疫システムを強化する保護因子として作用する(Maas et al 2006; Rook 2013)。
COVID-19の蔓延に対する生物多様性だけでなく、環境要因や生態系要因の総合的な影響は、特に生態系の範囲が広い地域では、経験的には不明のままである。我々の知る限りでは、世界レベルでのCOVID-19と生物多様性の関係についての情報はない。生態系と生物多様性の決定要因を考慮した COVID-19 の拡散と死亡率の経時的な推定は、これらの要因のどのレベルが拡散と死亡率を遅らせるのに有益で、どの程度の影響があるのかを特定するのに役立つかもしれない。我々の見解では、環境と生物多様性のパラメータの影響を他の要因と合わせて推定することは、将来の発生を封じ込めるための早期決定を行う際に、当局の指針となりうる洞察を与えることができる。したがって、本研究の目的は、生物多様性、環境、およびその他の生態系要因とCOVID-19の蔓延と世界レベルでの死亡率との関係を評価することである。
2. 研究方法
2.1. 研究デザイン
レトロスペクティブ、観察的、縦断的研究を実施した。218 カ国から COVID-19 の普及と死亡率、および関連する危険因子に関するデータを取得した。WHO(https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports/)が報告している最新の公開された人口レベルの情報(国別)に基づき 2020年1月21日から5月18日までのCOVID-19の1日の症例数と死亡数のデータセットを編纂した。本研究は、Parc Sanitari’s Sant Joan de Déu, Ethics Committee(PIC-67-20, Barcelona, Spain)の承認を得ており、1975年ヘルシンキ宣言の倫理ガイドラインに準拠している。
2.2. COVID-19国際データおよびその他のベースライン対策
WHOの日次状況報告書を用いて、218の国・地域ごとに 2020年1月21日から5月18日までの間に、1日の確定症例数、総確定症例数、1日の死亡確定症例数、死亡確定症例数の総量、最後に報告された症例からの経過時間のデータを集計した。クルーズ船で確認された症例は分析対象から除外した。中国全省の症例をまとめてグループ化した。COVID-19の症例は、特に香港、マカオ、台湾などの中国の行政区では、中国本土とは異なる政府の介入や政策措置が適用されているため、個別に分類した。WHOのデータベースに基づき、プエルトリコ、北マリアナ諸島、グアム、米国バージン諸島を米国とは別に分類した。
国ごとの各社会的遠距離介入の有効日をオンラインの政策データベース(世界保健機関 2020)から最初に抽出し、「低」、「中間」、「高」、「非常に高」の介入レベルを表すレベル0から3まで変化させた4レベルの政府政策介入スコアを作成した。レベル0(「低」)は、入国時の温度チェックや追加のメディカルスクリーニング(入国時)または入国時の旅行健康アンケートの記入、または被災地から輸入された疑いのある症例の検疫を各回ともに実施していない国、レベル1(「中級」)は、「低」の措置に加えて、特定の旅客に対するビザの停止や入国停止、またはCOVID-19の被災地との間のフロー停止を発表した国と定義されている。レベル2(”高”)とは、上記のすべての措置を講じた上で、大規模な集会やイベント、教育活動(学校や大学の閉鎖)の計画や中止、特定の地域の隔離などを行った国を指す。レベル3(「非常に高い」)とは、上記のすべてを適用し、国全体の隔離(完全または強制的なロックダウンと定義)を行った国、自宅待機命令を発表した国、または全国的な外出禁止令を適用した国(特定の政府命令で、国民が自宅待機を要求されたり、外出を許可されたりする期間を指すものと定義され、夜間または昼間の外出禁止令が考慮された)と定義される。上記の方法論の詳細は、Tyrolovas er al)。
世界銀行の分類システムを用いて、218カ国をそれぞれの国の所得レベルを「高所得(HICs)」、「上所得(UMICs)」、「中所得(LMICs)」、「低所得(LICs)」に分類した。また、異なる地理的地域を定義するために、WHOの分類を以下のように使用した。ヨーロッパ地域、西太平洋地域、アメリカ大陸地域、アフリカ地域、東地中海地域、東南アジア地域とした。また、1平方キロメートル当たりの人口密度も世界銀行のデータ(Bank, 2016)から国別に評価した。
最後に、さらに2つの変数を用いた。”最初の事例からの日数」と「最後の事例からの日数」である。前者の変数は、各国で最初のCOVID-19症例が報告されてからの日数を測定する。この変数は、たとえ疾病の開始時期が異なっていたとしても、すべての国を比較することを可能にする。したがって、それはある地域への病気の到着を記述している。さらに、”最後の症例からの日数 “はWHOによって作成された変数で、ある国がCOVID-19の新しい症例を報告しなかった日数を測定する。したがって、これはある地域での病気の広がりの加速または減速を記述している。
2.3. 国別の生物多様性、環境、生態系の評価
生物多様性条約(https://www.cbd.int/gbo1/annex.shtml)で報告されている生物多様性指数(NBI)は、各国の遺伝的、種的、生態系レベルの変動を測る指標であり、4つの陸生脊椎動物クラスと維管束植物の豊かさと固有性の推定値に基づいている;脊椎動物と植物は等しくランク付けされている。NBIには国の大きさの調整が含まれており、値の範囲は0~1である。土地面積が 5000 平方キロ未満の国、海外領土、および依存関係にある国は除外した。
毎日の気温と降水量は、NASA の Goddard Earth Sciences Data and Information Services Center (GES DISC) からした。具体的には、Integrated Multi-satellite Retrievals for Global Precipitation Measurement(https://disc.gsfc.nasa.gov/datasets/GPM_3IMERGDE_06/summary)(IMERG)と呼ばれるアルゴリズム(Nasa ED, 2020a)からデータを取得した。precipitationCaの測定では、マイクロ波と赤外を組み合わせたスペクトルの降水量を測定している。さらに、MERRA-2 (a Modern-Era Retrospective analysis for Research and Applications version 2) (Nasa ED, 2020b) (disc.gsfc.nasa.gov/datasets/M2SDNXSLV_5.12.4/summary)から、各国の地上2mからの1日の最高気温、最低気温、平均気温を取得した。多重共線性を避けるために、我々のモデルでは最高気温のみを使用した。気温と降水量の測定値と症例と死亡率の確認との間の 14 日間を考慮して、14 日間のラグをとったことに注意する。このラグアプローチは、以前にも他の研究で同様に適用されている(Shi et al 2020;Runkle et al 2020)。
国ごとの生態系の活力と環境の健全性に関連する要因を表1に示し 2020年環境パフォーマンス指数(EPI)報告書(Index EP, 2018)から取得した(https://epi.yale.edu/downloads)。世界各地の持続可能性の状態をデータに基づいてまとめたものがEPIで提供されており、11の課題カテゴリーにまたがる32のパフォーマンス要因を使用している。データは、国際的な統治機関、非政府組織、学術研究センターなど、信頼できる第三者の情報源から入手している。信頼できるデータセットは、科学界の査読を受けた、あるいは国際的な権威に支持された、確立された収集方法に基づいている。
表1
以下のリストは、「2020年環境パフォーマンス指標フレームワーク」の環境・生態系の活力要因を示したものである。フレームワークでは、32の要因を11の課題カテゴリーと2つの政策目標に整理している。コードは各要因の変数を指定している。出典は以下の通り。Index EP, 2018)。
| 政策目標 | 問題のカテゴリ | 因子 | コード |
|---|---|---|---|
| 環境衛生 | 大気質 | 周囲の粒子状物質による汚染 | PMD |
| 固形燃料による家庭の空気汚染 | 持っていました | ||
| オゾン | OZD | ||
| 衛生と飲料水 | 安全でない飲料水 | UWD | |
| 安全でない衛生 | 米ドル | ||
| ヘビーメタル | 鉛暴露 | PBD | |
| 廃棄物管理 | 固形廃棄物 | MSW | |
| 生態系の活力 | 生物多様性と生息地 | 陸生生物群系保護–国の重み | TBN |
| 陸生生物群系保護–グローバルウェイト | TBG | ||
| 海洋保護 | MPA | ||
| 保護地域代表性指数 | パー | ||
| 種の生息地指数 | 市 | ||
| 種保護指数 | SPI | ||
| 生物多様性生息地指数–維管束植物 | BHV | ||
| 生態系サービス | 樹木被覆損失、% | TLC | |
| 草地の喪失 | GRL | ||
| 湿地の喪失 | WTL | ||
| 漁業 | 魚の在庫状況 | FSS | |
| 地域海洋栄養指数 | RMS | ||
| トロール網で釣った魚 | FGT | ||
| 気候変動 | CO 2強度のトレンド | CDA | |
| メタン強度の傾向 | CHA | ||
| Fガス強度の傾向 | FGA | ||
| N 2 Oの強度トレンド | NDA | ||
| ブラックカーボン強度トレンド | BCA | ||
| GHG排出原単位の成長率 | GIB | ||
| 一人当たりのGHG排出量 | GHP | ||
| CO 2土地被覆から、トレンド | LCB | ||
| 汚染物質の排出 | SO 2強度トレンド | SDA | |
| NO Xの強度の傾向 | NXA | ||
| 農業 | 持続可能な窒素管理指数 | SNM | |
| 水資源 | 廃水処理レベル | WWT |
本研究では、5つの要素に分解された大気欠損2020年EPIを重視している。家庭用固形燃料(HAD)は、家庭での固形燃料の使用による室内大気汚染への曝露による実際の結果を測定する。周囲粒子状物質汚染(PMD)は、各国の代表的な国民が曝露されるPM2.5の年間平均濃度を測定する。オゾン(OZD)は、各国の代表的な国民が曝露されている地上レベルのオゾンの強度を測定する。前者の3つの指標の単位は、10万人あたりの年齢標準化された障害調整寿命年の喪失数である。また、SO2排出量(SDA)とNOx排出量(NXA)は、経済全体からのSO2とNOx排出量の強度を、それぞれ現在の強度と10年の傾向をブレンドしたものとして測定する。これらの測定値の算出方法の技術的な詳細は、EPIの技術的な付録(https://epi.yale.edu/downloads/epi2020technicalappendix20200604.pdf)に掲載されている。EPIレポートのすべての変数について、我々は2020年のEPIの値を使用し、また、指標の増分または減分を含めるために10年間の変化率を使用している。2020年EPI報告書は、前年の大気質および汚染物質対策を計算している。
2.4. 都市別の大気測定値
世界大気質指標プロジェクト(https://aqicn.org/data-platform/COVID-19)から、気温、湿度、地表面オゾン、大気中の粒子状物質の粒径10μm以下(PM10)粒径2.5μm以下(PM2.5)の日平均値の情報を取得した。これらの指標は都市別のものであり、COVID-19の広がりのレベルが異なる3都市を使用した。デンバー(中程度のケース率)バルセロナとミラノ(高いケース率)である。データは2020年3月8日から5月18日までのものを使用した。
2.5. 統計的分析
2.5.1. ベイズ時空解析
COVID-19の陽性症例数と死亡者数に関連する生物多様性、気温、降水量の要因の影響を明らかにすることを目的とした。異なる国をまたいで時間的に収集したデータを利用しているため、ベイズ的時空間アプローチを用いて、結果と共変量の間の影響を把握している。特に、このアプローチでは、COVID-19の拡散と死亡例の性質から重要であることが証明されている時間的相関と時空間相互作用を考慮している。さらに、統計的に健全な推論を導くために、過分散と過剰ゼロを示す、関心のある結果のゼロインフレーション負二項分布(ZINB)を仮定した。
COVID-19の拡散と死亡率の進化を評価するための時空間モデルは、以下の交絡因子を調整して適用した:政府の介入レベル(4段階の厳しさ)COVID-19の新規症例が最後に報告されてからの日数(最後に報告されてからの日数)降水量と気温の測定、国の所得水準、各国での最初のCOVID-19の症例が報告されてからの日数(最初に報告されてからの日数)および人口密度(平方キロメートルあたり)。総人口の自然対数は、国ごとの感染率と死亡率を考慮するために(オフセットとして)線形予測関数に加えた。我々は、デビアンス情報基準(DIC)(Spiegelhalter et al 2002)と渡辺・赤池情報基準(WAIC)(Watanabe, 2010)の両方に基づいて、最適なモデルを選択した。すべてのモデルの詳細は、S1に記載されている。ベイズ時空モデルは、空間的・時間的効果を捉えるために、今回の分析の最初のステップとして使用された。すべての計算は、R バージョン 4.0.2 の R パッケージ R-INLA (Rue et al 2009) を使用して行われた。
2.5.2. 一般化線形混合モデル
また、COVID-19の拡散と死亡率に関連する生態系の活力と環境衛生変数の影響を明らかにするために、Template Model Builder(glmmTMB)を用いて一般化線形混合モデルを適合させ、関心のあるアウトカムにZINB(ZINB)分布を仮定した。解析期間は1月21日から5月18日までとした。
今回の分析で使用した要因は、表1に示したものである。予備分析として、すべての変数タイプが数値であることを考慮して、多重共線性の問題を避けるために、それらの間の相関関係を確認した(使用した最終的な変数セットとの相関行列については、補足図S1を参照してほしい)。glmmTMBモデルは、以下の交絡因子を調整して適用された:政府介入レベル(低、中、高、非常に高)国の所得レベル、各国における最初のCOVID-19症例からの日数、世界銀行の地理的地域分類、および最後の2つの交絡因子間の相互作用。我々は、赤池情報基準(AIC)(Akaike, 1974)に基づいて最適なモデルを選択した。さらに、我々のデータは、国ごとに経時的に測定した縦断的研究から来ているので、国のグルーピング変数にはランダム効果を含めた。しかし、我々はまた、経時的な国内の相関は一定ではないと仮定している。したがって、我々は、国内のオブザベーション間の相関が時間の関数であることを暗示するランダム切片とランダム・スロープ・モデルを含みた。上記で述べた交絡因子は、モデルの固定効果として組み込まれた。我々がベイズ時空分析で適用したように,母集団オフセットがモデルに考慮された。同様の方法論的アプローチは、この分野の先行研究によって適用されている(Wu er al 2020; Travaglio er al 2020)。すべてのモデルの詳細は、補足付録S2に示されている。
解析的なモデルをフィッティングすることを目的として、ランダムな傾きの仮定なしで同じモデルをフィッティングした。2つのモデルの適合を比較するために尤度比検定を計算した(ランダム・スロープありとなし)。その結果、ランダムスロープを含むモデルは、単純なモデルよりも有意に良好であるという証拠を示した(p値<0.001)。最後に、我々はまた、地域間のモニタリングインフラとモニタリングシステムのウイルスの拡散と死亡率の変動を制御するために、国の所得水準によって層別化した同様の回帰モデルを適合させた。このようにして、低・中所得国(94カ国-58.7%)と高・中所得国(66カ国-41.3%)の2つのグループを作成し、それぞれの所得水準グループが医療の質を均質に反映していることを確認した。すべてのモデリングは、Rバージョン4.0.2のRパッケージglmmTMB(Brooks et al 2017)を用いて実施した。
2.5.3. 一般化線形モデルに従ったカウント時系列の分析
COVID-19の拡散と大気質因子との関連を特徴づけるために、カウント時系列をフィットさせ、自己回帰項を持つ負の二項モデルに従った。特に、大気中の粒子状物質の直径が10μm以下(PM10)2.5μm以下(PM2.5)地上レベルのオゾン(O3)がCOVID-19の症例数に関連して有意な効果を持つかどうかに関心があった。湿度と温度の交絡因子で調整したすべてのカウント時系列モデルを評価し、Rバージョン3.6.3のRパッケージtscount(Liboschik et al 2015)を用いて解析を行った。加えて、COVID-19の散布とすべての大気質因子との間のスピアマン相関を計算した。大気質因子の日付は、伝播から症例確認までの14日間を考慮して、それぞれのCOVID-19症例の2週間前の日付であることに注意した。
3. 結果
社会人口統計学、国別所得水準、政府政策介入要因に関するデータを 208 ヶ国すべてから取得した。欠損値の存在により、サンプル国は 192 カ国(90.8%)に減少した。この 192 カ国のうち、174 カ国(90.6%)と 180 カ国(93.8%)が、それぞれ生物多様性指数(National Biodiversity Index)と EPI 報告書の情報を得ていた。欠落データのパターンが異なるため、最終的なデータは、ここで報告された分析に関連するすべての変数を持つ 160 カ国のデータとなった。国の所得水準の分布は、低所得国を除くすべてのグループでほぼ同様である(HICs:29.4%、UMICs:28.8%、LMICs:25.6%、LICs:16.3%)。地域分布は降順である。欧州・中央アジア(29.5%)サハラ以南のアフリカ(27.5%)中南米・カリブ(16.4%)東アジア・太平洋(11.9%)中東・北アフリカ(9.4%)南アジア(4.4%)北米(1.3%)の順となっている。
3.1. COVID-19の蔓延と死亡率と各国の生物多様性指標との関連:世界的・時空間的分析
COVID-19の普及率(10万人単位の症例数)とNBIの生データを図1の世界地図に表示した。COVID-19の症例数とNBIについては、比較できるように色のスケールを0から 1の間に設定した。すなわち、一方の地図では大きな値(赤みがかった色調)が他方の地図では小さな値(青みがかった色調)に対応しており、その逆もまた然りである。これらのマップは、NBIとCOVID-19の症例数との間に負の相関関係があることを視覚的に示している。
図1 1月21日から5月18日までのNBI値(左の地図)とCOVID-19の分布(右の地図)の世界地図
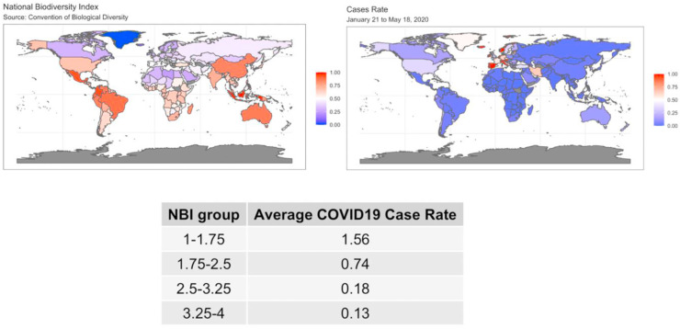
図1に示したサマリー表は、この一変量的な関連性を確認している。この表は、NBIの異なるグループ(四分位を用いて分類)のCOVID-19の症例率の平均を示している。例えば、NBIの低い値に対応するCOVID-19の症例率は1.56であり、NBIの高い値の症例率0.134から急激に増加している。
時空間回帰分析は、政府の介入、所得水準、生物多様性、環境、その他の要因と合わせて、160カ国におけるCOVID-19の普及状況を評価したものである。
表2 COVID-19の普及を評価するためのベイズ時空間回帰分析
| アイテム | 推定係数 | 95%HPDI |
|---|---|---|
| NBI | −0.606 | −0.946、-0.268 |
| 沈殿物Ca | −0.001 | −0.003、0.001 |
| 温度(最大) | −0.010 | -0.017、-0.002 |
| 人口密度(平方キロメートル) | 0.0003 | −0.0002、0.0006 |
| 最後のケースからの日数 | −0.015 | −0.041、0.010 |
| 最初のケースからの日数 | −0.009 | -0.016、-0.002 |
| HIC | 参照カテゴリ | |
| LIC | −0.334 | −0.801、0.132 |
| LMIC | −0.275 | −0.636、0.086 |
| UMIC | −0.305 | −0.612、0.000 |
| NBI:温度(最大) | −0.011 | −0.023、0.002 |
95% HPDIがゼロ値を含まない有意効果は太字で示す。HPDI: Highest Posterior Density Interval, ベイズのフレームワークにおける等価CI。LMICs. 下位中所得国; UMICs. UMICs: Upper Middle-income countries; LICs. LICs: 低所得国。NBI. precipitationCa: マイクロ波と赤外線を組み合わせたスペクトルの降水量を測定する。最後のケースからの日数:前回のCOVID-19の新規ケースからの日数。最初の症例からの日数:各国でCOVID-19の最初の症例が報告されてからの日数のカウント。
時空間モデルは、政府の政策介入のために調整された。
ある国の生物多様性の程度は、COVID-19の世界的な広がりと逆の関係にあった[NBI:-0.61,95%CI(-0.95, -0.27)]。NBIの影響はこのモデルでは統計的に最も強く、遺伝的、種的、生態系的レベルでの国の変動が低いほど、COVID-19の拡散レベルへの影響は高かった。有意性は低いが、最高気温2mの逆効果もあり(表2参照)気温の高い国ではウイルスの拡散が低いことが示唆された。COVID-19の拡散に関連した生物多様性と気温の間には有意な交互作用は見られなかった。降水量についても同様の観察がなされた。しかし、最初に報告されたCOVID-19症例からの日数はCOVID-19症例数と反比例していた[最初の症例からの日数:-0.01, 95%CI(-0.02, -0.002)]。
また、ベイズ時空間モデル、一般化線形混合モデルを用いて死亡率の進化を解析した(詳細な結果はS3の表S1を参照)。その結果、生物多様性や環境要因は死亡率に直接的な影響を与えないことが示された。
3.2. COVID-19の散布と死亡率と生態系の活力や環境衛生変数との関連性
図2は、表3に示した生態系対策のための一般化混合ZINBモデルを用いて推定したCOVID-19の散布に有意な影響を与える要因を示したものである。
図2
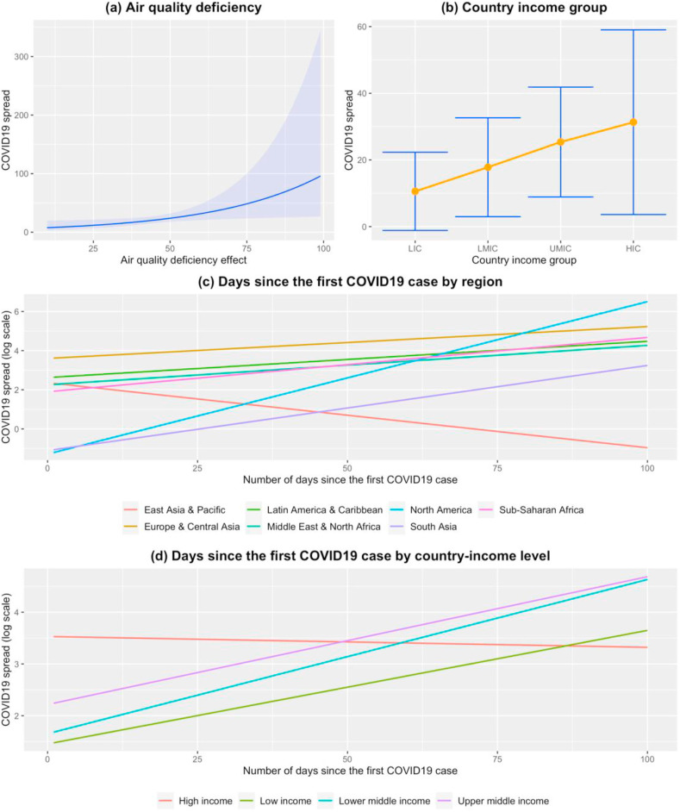
(a)大気質異常、(b)国別所得グループ、(c)地域別、(d)国別所得グループにCOVID-19の拡散に及ぼす影響をプロットしたものである。COVID-19のスプレッド単位は、関連するモデルによって生成されたオフセットを経由したレートに基づいている。
表3 COVID-19の普及を評価するための一般化混合ZINBモデル回帰分析
| アイテム | 推定係数 | 95%CI |
|---|---|---|
| HIC | 参照カテゴリ | |
| LIC | −2.080 | −3.842、-0.311 |
| LMIC | -1.880 | −3.425、-0.332 |
| UMIC | −1.310 | −2.515、-0.114 |
| 空気不足 | 0.028 | 0.004、0.053 |
| 空気不足10年の変化 | −0.020 | −0.142、0.101 |
| 衛生・飲料水10年変更 | 0.001 | −0.140、0.143 |
| 重金属10年の変更 | 0.030 | −0.116、0.177 |
| 生物多様性と生息地 | −0.001 | −0.019、0.017 |
| 生物多様性と生息地の10年間の変化 | 0.005 | −0.028、0.038 |
| 生態系サービス | 0.006 | −0.009、0.020 |
| 気候変動10年の変化 | 0.0002 | −0.021、0.021 |
| 汚染物質の排出 | −0.014 | −0.033、0.006 |
| 汚染排出量10年の変化 | 0.009 | −0.004、0.023 |
| 農業 | −0.010 | −0.031、0.011 |
| 農業10年の変化 | −0.004 | −0.036、0.029 |
| 最初のケースからの日数:東アジアと太平洋 | 参照カテゴリ | |
| 最初のケースからの日数:ヨーロッパと中央アジア | 0.050 | 0.024、0.075 |
| 最初のケースからの日数:ラテンアメリカおよびカリブ海 | 0.052 | 0.023、0.080 |
| 最初のケースからの日数:中東および北アフリカ | 0.053 | 0.023、0.084 |
| 最初のケースからの日数:北米 | 0.111 | 0.050、0.172 |
| 最初のケースからの日数:南アジア | 0.077 | 0.036、0.117 |
| 最初のケースからの日数:サハラ以南のアフリカ | 0.061 | 0.032、0.090 |
| 最初のケースからの日数:HIC | 参照カテゴリ | |
| 最初のケースからの日数:LIC | 0.024 | −0.006、0.054 |
| 最初のケースからの日数:LMIC | 0.032 | 0.009、0.054 |
| 最初のケースからの日数:UMIC | 0.027 | 0.007、0.046 |
有意な効果を太字で示す。LMICs. 下位中所得国; UMICs. 上位中所得国;LICs. 低所得国。
一般化混合ZINBモデルは、最初のケースからの日数、世界銀行の地域、政府の政策介入についても調整された。
様々な交絡因子を調整した後 2020年のEPI変数としては、単一ではあるが関連性のあるもの、すなわち空気欠乏のレベルのみが見つかった。この変数は、家庭の大気汚染(55%)2.5μm以下の微小粒子状物質(PM2.5,40%)地上レベルのオゾン汚染(5%)への曝露の加重平均を用いて構築されている。
図2aは、大気質不良と関連づけた場合のCOVID-19の広がりに対する影響の増大を示したものである。4つの因子のうち、健康への影響が最も大きいのは大気の質であり、大気欠損の値が大きいほどCOVID-19の拡散拡大効果が大きくなっている[大気欠損:0.028,95%CI(0.004, 0.053)p値=0.021]。COVID-19の伝播に有意に関連する追加の共変量があった。ウイルスの拡散に関しては、図2に描かれているように、HICは最悪の効果を有していた。さらに、COVID-19の伝播については、HICsと比較して、HICsが最も高く、LICsが最も低いという一貫した勾配を示した[LICs:-2.080, 95%CI(-3.842, -0.311), p値=0.021]。しかし、この関係は、世界旅行の性質など、いくつかの他の基礎となる変数によって駆動される可能性があることに留意すべきであり、それはおそらく調査期間においてHICに偏っていたのであろう(Menkir et al 2020;Shellekens and Sourrouille 2020)。COVID-19の最初に報告された症例からの日数は、東アジア・太平洋地域を除くすべての世界銀行地域においてCOVID-19症例の増加と正の相関を示した(図2c参照)。
また、興味深い相互作用も観察された。COVID-19 症例報告からの日数については、LMICs と UMICs が HICs に比べてウイルスの拡散に有意な影響を与えていた(図 2d)。LMICs 0.032, 95%CI (0.009, 0.054), p値=0.005; 初発からの日数。UMICs:0.027,95%CI(0.007,0.046)p値=0.006]。
同じモデル化戦略の下で同じセットの共変量を用いた死亡率の解析(結果は補足付録の表S2および図S2に示されている)は、大気質の欠如が死亡率の進展に与える影響が小さいことを示している。大気質は、病気の広がりの解析よりも死亡率の解析の方が有意ではなかった。さらに、世銀地域全体で最初に報告された症例からの日数は、COVID-19の死亡率分析において唯一の有意な予測因子であり、COVID-19の拡散分析と同様の結論であった。
両モデルのランダム切片の分析では、我々の分析で報告された基礎となる共変量の違いをすべて考慮した後でも、国ごとのばらつきを示している[拡散モデル. 3.205, 95%CI (2.444, 4.204); 死亡率モデル。4.052,95%CI(2.939,5.586)]。これは、すべての共変量をゼロにしたときのランダム変動が有意であることを示している。
3.3. COVID-19の散布と死亡率の大気質対策との関連性
COVID-19の蔓延と死亡率に対する空気欠乏の影響が観察されていることから、この影響をさらに調査するために、前節で説明したような類似の混合効果モデルを適用した。大気欠乏の変数は、3つの具体的な大気質指標に分解されている。大気汚染(HAD)PM2.5曝露(PMD)地上レベルオゾン曝露(OZD)である。我々の分析では、表1に記載されている他の生態系の活力と環境衛生変数も使用し、それらを前述と同じ一連の交絡因子で調整した(COVID-19の拡散と死亡率のモデル推定値は、補足付録S4の表S3とS4に記載されている)。図3は、両アウトカムの有意因子の効果を示したものである:拡散(図3a参照)と死亡率(図3b参照)。
図3 COVID-19の散布(a)と死亡率(b)に対する大気汚染(HAD)の影響
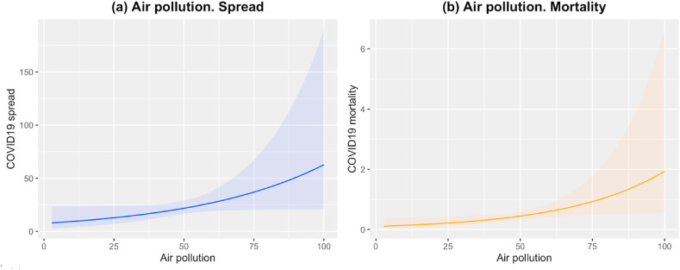
(a)の青線はCOVID-19の散布に対する大気汚染の影響を示し、上下の帯は95%CIを示す。同様に、(b)のCOVID-19の死亡率の変化についても、その影響とバンドはオレンジ色で表示されている。COVID-19の広がりと死亡率の単位は、関連するモデルによって生成されたオフセットを介した率に基づいている。(この図の凡例での色の参照の解釈については、この記事のウェブ版を参照してほしい)。
図3aおよびbは、大気汚染への曝露における同様の傾向を示しており、その影響はCOVID-19の広がりよりも死亡率に有意である[大気汚染(広がり):0.021,95%CI(0.001,0.043)p=0.049;大気汚染(死亡率):0.029,95%CI(0.005,0.054)p=0.019]。大気汚染のレベルが大きいほど、その要因がCOVID-19の蔓延と死亡率に及ぼす影響が大きいことを示している。他の分析された大気質因子は、散布や死亡率に影響を与えていないようである。
3.4. COVID-19の拡散および死亡率と大気汚染のレベルとの関連 国-所得レベルによる層別化
国によっては、COVID-19の蔓延と死亡率を他の国のように同じレベルで捉えていない可能性がある。この前提に基づき、前2節で紹介した混合モデルの分析を国-所得レベル別に層別化した。
その結果の解析結果をS4の表S5~表S8に示したが、高・中所得国においては、大気の欠乏が一貫して疾病の蔓延と死亡率に影響を与えていることが示された[大気欠乏(蔓延):0.037,95%CI(0.009,0.064)p値=0.008,大気欠乏(死亡):0.032,95%CI(0.003,0.062)p値=0.031]。補足付録S3の図S3のプロットは、空気欠乏が病気の蔓延とその死亡率に及ぼす影響を示している。ただし、低所得国、低中所得国の要因を考慮した場合には、空気欠乏の影響は認められなかった。
3.5. COVID-19と特定の大気汚染物質との感染拡大関連;3都市の分析
我々の時系列モデルは、各都市について、バルセロナ[O3: 0.04, 95%CI = (0.02, 0.08), p値<0.001]とミラノ[O3: 0.02, 95%CI = (0.01, 0.03), p値=0.003]で地上レベルオゾンに有意な効果を示し、これは、過去14日間の地上レベルオゾンの高い値がCOVID-19の拡散を増加させることを示唆している。10μm以下の大気中微粒子物質(PM10)については、デンバーでも同様のパターンが観測された。このように、PM10は分析期間中、COVID-19の日次症例と正の相関を示した[PM10: 0.02, 95%CI = (0.06, 0.26), p値 = 0.002]。モデル推定値の詳細は、補足付録S4の補足表S9に示されている。
図4は、3都市のCOVID-19時系列を14日遅れの日別PM10レベルまたはO3レベルと重ね合わせたもので、灰色の網掛け部分は、3都市のPM10レベルまたはO3レベルを表している。灰色の網掛け部分は、国レベルと同様の方法で、各都市が厳格な介入レベル(「非常に高い」)であった期間を示している。この点は、PM10とO3の時系列が14日遅れであるため、COVID-19の日別事例の時系列のみが関係している。したがって、これらの汚染物質対策の時系列は、「高い」レベルの介入が実施された期間にあり、したがって、大気汚染物質を発生させる可能性のある事業活動や産業活動がまだ存在していた期間であった。COVID-19の毎日の事例に有意な影響を与える要因のみを各グラフに示した。各都市で同様の傾向パターンが観察され、いくつかのピークが同時に発生している。例えば、デンバーのプロット(c)では、3月末から4月にかけて、PM10とCOVID-19の日別発生件数が同時にピークを迎えている。次に、COVID-19の事例と大気汚染物質測定値との間のスピアマン相関を計算した。その結果は、時系列で観察されたことを支持した[バルセロナCOVID-19散布とO3:ρ=0.38,p値=0.001;ミラノCOVID-19散布とO3:ρ=0.55,p値<0.001;デンバーCOVID-19散布とPM10:ρ=0.48,p値<0.001]]。
図4 バルセロナ(a)、ミラノ(b)、デンバー(c)のCOVID-19の地上レベルオゾン(O3)と大気中粒子状物質PM10とPM2.5の日例の時系列
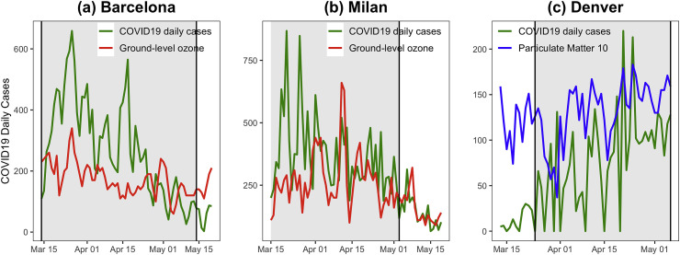
灰色の網掛け部分は、都市が厳しい介入レベルにあった期間を強調している。
4. 議論
エボラやSARSのような他の主要な既知の伝染病と同様に、COVID-19の出現は、私たちが経験している気候や生物多様性の危機と無関係ではない。人間の健康が、人間が自然界に介入することと密接に関連しているという証拠がある(Seymour, 2016; Thompson Coon er al)。 私たちの研究は、環境、生物多様性、および生態系要因と 2020年1月21日から5月18日までの間のCOVID-19の蔓延および死亡率との関連に焦点を当て、ここでは、いくつかの社会人口統計学的、社会的距離、および郡所得レベルの要因を調整した。この結果には、本稿で紹介した以上に議論に値するいくつかの側面がある。
第一に、国の生物多様性のレベルが病気の蔓延に与える影響は中程度であった。
第二に、COVID-19の伝播は、気温が高い国ほど小さいように見えた。
第三に、空気不足のレベルとCOVID-19の蔓延と死亡率の進化との間には直接的な影響があった。特に、家庭での固形燃料の使用(表1のHAD変数を参照)による室内空気汚染への曝露が高いほど、COVID-19の蔓延と死亡率の両方に高い影響を与えている。
第四に、大気の質の欠如は、LICsとLMICsよりもHICsとUMICsにおける疾病の蔓延と死亡率に一貫して影響を与えていた。
第五に、都市の事例研究で示されているように、COVID-19の伝播は、地上レベルのオゾンと大気中の粒子状物質のレベルが10μm以下の14日遅れと関連している。
COVID-19の出現は、生物多様性の損失に対する戦闘の極端な重要性を強調している(WHO 2017)。この新しい病気と生物多様性の減少との関係についての証拠の取り付けは、緊急の注意を必要としている(Lorentzen er al)。 より多くの生物多様性を持つ生態系では、病原体の迅速な拡散は困難である。生物多様性の喪失は、感染症の伝播に影響を与え(Keesing et al 2006)ウイルスが動物と人との間を通過する機会を提供する(Keesing et al 2010)。研究者らは、自然生態系の乱れが野生動物と人との間の病気のジャンプを増加させることを報告しており、これが無視され、忘れ去られ、最近の未知のヒトの病気の発生の主な原因であることが示唆されている(Epstein et al 2003)。例えば、3つの研究では、米国における鳥類の多様性の低さとヒトの西ナイル脳炎のリスクまたは発生率の増加との間の強い関連性が検出されている(Allan et al 2009;Ezenwa et al 2006;Swaddle and Calos 2008)。NBIに基づく本研究の結果は、これらの評価と一致している。このように、生物多様性の保全と持続可能な管理は、来たる世代の健康と福祉を守るために、気候の混乱を緩和し、パンデミックを防ぐために必要であるかもしれない(国連事務総長 2020)。我々の調査結果は、生物多様性の要因とCOVID-19の蔓延との間に関係があることを示唆している。我々の研究は、基礎となるデータ収集条件と、1月21日から5月18日までの期間にわたって、この可能性のある関係を浮き彫りにすることを目的としている。しかし、この関係は、社会経済的発展のようないくつかの他の基礎となる変数によって駆動される可能性があることを認識しており(Travaglio et al 2020年)したがって、さらなる分析が必要である。
新しい研究では、人間に飛びつく病原体や寄生虫を宿主とすることが知られている野生動物は、人間が自然の生息地を二次的、農業的、都市的な生態系に変えてしまった場所では、その地域の動物のシェアが高くなることが示されている(Gibb et al 2020)。この研究では、げっ歯類、コウモリ、スズメ目の鳥などの宿主種との人獣共通感染症について、後者の効果が最も強いことがわかった。これらの種の中には、人間が生物多様性を減少させた後に生き残った種もあり、その結果、人間に飛躍する危険な病原体のリスクが高くなる(Tollefson, 2020) しかし、人間の発達による生物多様性の喪失と疾病の発生との間の後者の関連性は、次のパンデミックや現在のCOVID-19の蔓延を予測するものではなく、現在のパンデミックとのメカニズムを確立するための調査を続けることを促している。
WHOは、脳卒中、心臓病、肺がん、慢性閉塞性肺疾患、肺炎を含む呼吸器感染症などの疾患につながる汚染された空気中の微粒子への曝露により、毎年約700万人が死亡していると推定している(WHO 2016)。大気汚染はまた、欧州公衆衛生連合(Vettore, 2020)によると、免疫システムを弱体化させ、感染症を撃退する能力を低下させることが知られている。特に、家庭の大気汚染は380万人の死亡に貢献している(WHO 2016)。2003年の研究では、COVID-19と密接な関係にある呼吸器ウイルス疾患であるSARSの患者は、汚染レベルの高い地域に住んでいると84%の確率で死亡することが明らかになった(Cui er al)。 私たちの研究では、大気汚染とCOVID-19との間にも関連性があることが示された。しかし、HICsとUMICsでは大気汚染の影響が高く、国によって影響が異なることがわかった。所得水準の異なる国間でのこの不一致の理由として考えられるのは、HICsとUMICsの方が工業化レベルが高く、それに伴って大気汚染のレベルが高くなっていることである(Vigo er al)。
先進工業国は、COVID-19の確定症例のうち全世界で530万人、現在までに全世界で35万人の死亡者を出している(WHO, 2020b)。これらの国々では、大気中の粒子状物質(PM10とPM2.5)と地上レベルのオゾン(O3)汚染物質のレベルが高くなる傾向があり、これらは自動車や産業排出物などの人為的な発生源から発生している。PM10とPM2.5は空気中に長く留まる傾向がある微粒子で、地上レベルのオゾン(O3)は刺激性ガスである。これらの汚染物質はすべて、呼吸器系の慢性疾患の引き金となったり、悪化させたりする(Vigo er al)。 これは科学的根拠のある深刻な健康問題である。例えば、ある最近の研究では、PM2.5の1立方メートルあたりわずか1μgの増加は、COVID-19の死亡者数の15%の増加に対応することがわかった(Wu et al 2020)。また、O3レベルの増加の結果は、PM10レベルのCOVID-19拡散への影響よりも悪いことが観察されている(Cui et al 2003; Australian Governament, 2020; Staehelin et al 2001; United States Environmental Protection Agency, 2020)。我々は、3つの先進都市におけるPM10, PM2.5, O3のレベルを調査し、これらの汚染物質がCOVID-19の拡散と関連していることを確認した。ヨーロッパの2都市(バルセロナとミラノ)ではCOVID-19の発生数とO3のレベルが相関しており、デンバーではPM10のレベルとの関連が観察された。我々の知見を裏付けるように、最近の研究では、PMレベルや他の汚染物質とCOVID-19の拡散との関連性が指摘されている(Travaglio et al 2020; Zoran et al 2020)。
環境および生物多様性の要因に関連した前述の知見とは別に、他の有意な影響があった。我々の研究を世界銀行の地域で層別化した場合、COVID-19の最初の症例が特定の世界銀行の地域で報告されて以来、日数が多くなるにつれて、COVID-19の拡散への影響はより大きくなった。唯一の例外は東アジア・太平洋地域であった。1つのもっともらしい説明は、この病気が最初にアジアで発生し、次にヨーロッパ、北米などで発生したことを知っているということである。したがって、アジアの地域は、後発の地域に比べてCOVID-19の感染が減少するまでの期間が長かったと考えられる。さらに、HICsが多く存在する地域では、COVID-19の影響を受けていることがわかった。また、私たちの分析では、裕福な国ほど、最初の症例からの日数が増えるほど、病気の伝播への影響が小さくなると判断された(Clouston er al)。 考えられる議論としては、HICsはCOVID-19検査の実施数が多く、国民の保健システムや組織が整っている一方で、LICsの政府には緩和策を実施するためのリソースが不足しているということが考えられる。
最後に、大規模国における地域拡大の観点からの推論を評価するために、感度分析を行った。感度分析では、面積の大きい上位 5 カ国(ロシア、カナダ、中国、米国、ブラジル)を除外し、COVID-19 の拡散と死亡率に関するデータを用いてモデルを再適合させた。これらの大国を除外することで、推論分析に影響を与えているかどうかを確認することができた。いずれの場合も、死亡率と散布率は確定モデルの推定値と一致しただけでなく、有意な環境共変量も5カ国を含めた場合と同様の効果を持つと推定された。所得共変量の変化が観察されたのは所得共変量のみであり、これは症例数の多いUMIC3カ国(ブラジル、ロシア、中国)を除外したことによるものと考えられる。時空間分析の結果を、拡散と死亡率について、それぞれS4の表S10と表S11に示す。
4.1. 限界
私たちの知る限りでは、この研究は、公開されているCOVID-19のデータを用いて、世界160カ国における疾病の蔓延と環境要因と生物多様性レベルに関連した死亡率を、社会人口統計学、国別所得水準、政府の政策介入の交絡因子で調整して分析した初めての研究である。しかし、注意しなければならないいくつかの限界がある。
- 我々の推論は観測データを用いている。データが利用可能な範囲で、我々の推論は観測単位間の共変量の差を調整している。理想的には無作為化デザインでモデルを適合させるべきであったが、現在の環境ではそのようなデザインを追求することは不可能である。しかし、この研究の延長線上には、マッチングや重み付けに基づいたランダムサンプリング法に基づくポストランダム化技術を用いることも可能であり、潜在的に変化する背景特性を特に対象としている。残念ながら、相関関係の複雑な性質や時空間データデザインの性質に対処するための手法は、すぐに利用できるものがない。相関構造の分析結果に基づいて、研究デザインの役割はこのような考察において重要であると予想される。
- 本研究は観察研究であり、以前の類似研究で報告されたように、この種の研究が持つバイアスの潜在的なリスクを含んでいる(Wu er al)。 したがって、本研究の結果は、個人レベルの推論的結論を出すために使用すべきではない。さらに、本研究はまた、測定されていない交絡バイアスを生じやすい可能性がある。しかし、我々は人口密度、パンデミック開始からの時間、政府の介入、天候、社会経済的要因などの最も重要な交絡因子の調整を試みた。さらに、本研究は観察的な性質を持っているため、報告された結果は特定のモデル化の選択に敏感に反応する可能性がある。このようなモデル化アプローチの感度を評価するために、国の所得レベルで層別化したモデルを適用した(S4の表S5-S8を参照)。
– WHOのCOVID-19データセットの完全性や、ある特定の日に発表され、数日後に効果的に適用される政府の介入など、いくつかの要因により、COVID-19の時間的な広がりと死亡率の傾向を決定的に捉えることはできないかもしれない。 - 信頼できる報告システムを持っている国もあれば、そうでない国もある。このように、国や地域ごとのCOVID-19の症例数や死亡者数のモニターやサーベイランスにはばらつきがある。例えば、COVID-19の症例数と死亡数が過小報告される可能性がある(Piovani et al 2020;Islam et al 2020)。それほどまでに、本研究では無症候性の症例を考慮に入れることができなかった(Oran and Topol, 2020)。それでもなお、各国は一定の基準を用いて、国レベルでの普及率と死亡率をWHOに報告しなければならない。その意味で、COVID-19の蔓延と死亡率と生態系の活力や環境健康要因との関連性についても、サーベイランスやヘルスケアシステムの多様性を捉えることを目的に、国の所得水準別に層別化して分析した(Barber et al 2017)。その結果、グローバルな分析との整合性が示された。
- 国ごとの毎日のスクリーニングCOVID-19検査(Udugama et al 2020)は、その情報を報告している国が限られていたため、使用されなかった。他の研究でも、同期間の国別COVID-19検査率は困難であると指摘されている(Islam et al 2020)。また、環境変数とNBI変数を持つ国の数が減っていたため、国のサンプルが少ないためにパラメータ推定に変化が生じる可能性があるため、COVID-19検査データを使用しないことを好んだ。
- 降水量と気温に関連する共変量を時空間モデルで使用した。これらの共変量は各国のセントロイドの降水量と気温のレベルを要約したものに過ぎず(例えば、スペインのRivas-Vaciamadrid(マドリッド近郊)の値しかない)国全体を代表するものではない。これは代理として使用されているだけであり、関連する誤差は、ゼロインフレーションされた負の二項確率分布に組み込まれている。しかし、この研究を拡張することで、一連の国のサブナショナルデータに焦点を当てた、より正確な地理的プロキシを使用することができる。
- 我々の分析は、国別の生物多様性指数(NBI)に基づいている。国の生物多様性の指標は、国の地域(都市部とそうでない地域)によって大きなばらつきがあることを理解しているが、COVID-19の感染は主に都市部で発生していたと考えられる。我々の交絡モデルと都市レベルのデータとの相関分析は興味深いものである。我々の知る限りでは、生物多様性の指標となる都市レベルのデータは公開されていない。さらに、最大面積を持つ上位 5 カ国(ロシア、カナダ、中国、米国、ブラジル)を除外した時空間モデルに感度分析を適用した。結果は同じ方向に推移した。
- 2020年環境パフォーマンス指数は、国レベルで最も更新されたデータセットである。ただし、大気質と汚染物質対策は前年のEPI報告書から算出されたものである。そのため、それらはCOVID-19の毎日の事例の時系列と同じ時期には測定されていない。そこで、本研究では、4ヶ月間の環境トレンドとCOVID-19の感染と死亡率との関連を評価することを試みた。
- 我々の死亡率モデルの限界は、人口の年齢密度や国ごとの平均寿命の間接的な指標を考慮していないことである。さらに、個人の微小環境(職場、学校など)や行動(屋外活動、喫煙習慣など)に関連する因子は、大気汚染への曝露と大きく関連しており(Travaglio et al 2020年)本研究では評価できなかった。
- 我々の分析の限界は、ローカルスケールでの分析を行うために、各国でより高い空間分解能を使用しなかったことである。同じデータセットを持っていても、ローカルスケールでの分析を行うことはほぼ不可能である。仮にそのようなデータがあったとしても、有意なパラメータの対応や信頼性の問題により、統計的推論の品質が標準以下になる可能性がある。さらに、これらの問題を補正するための統計的介入が必要となるが、残念ながら本研究の範囲を超えている。
我々の調査と分析は 2020年1月21日から5月18日までのCOVID-19の蔓延と死亡率のデータの変動に焦点を当てた。国のサブセットであるLIC、LMIC、UMIC、HICについて、症例数の中央値の5月18日までの傾向と6月30日までの傾向を比較するグランジャー検定を用いた感度分析を行った。その結果、HIC国とLIC国の6月30日までの傾向は、5月18日までの傾向とほぼ同じであった。しかし、LMIC国とUMIC国では5月18日までの傾向と5月18日以降の傾向が大きく異なっていた。したがって、本論文の結果は、あくまでも基礎となるデータ収集条件と期間に関連するものであるため、慎重に解釈すべきである。COVID-19は動的な伝播を伴う感染症であり、我々が使用したすべての共変量が変化する可能性があるため、さらなるデータと分析が必要となるため、5月18日以降の結論を出すことは適切ではないと考えている。
5. 結論
COVID-19パンデミックは、世界的な生物多様性の喪失と環境衛生の危機に関連した結果であると考えられる。本研究では、生物多様性の指標である大気質変数とCOVID-19の感染と死亡率との関連性の程度と有意性を測定した。大気汚染の削減と生物多様性の保全は、政府の行動と財源に依存する複雑な問題である。本研究で得られた知見は、COVID-19の新たな発生に対応し、将来の危機を防ぐための代替戦略として、政府の環境・健康政策の立案に役立つ可能性がある。