
PP2Aとアルツハイマー病
概要
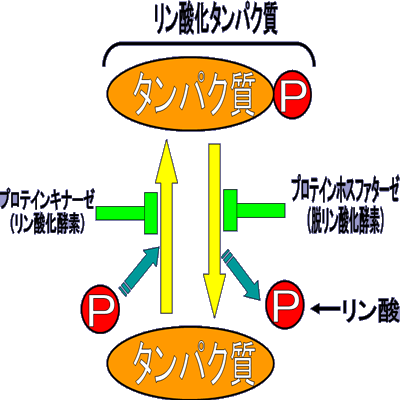
細胞は、機能を維持するために細胞内をタンパク質のリン酸化と脱リン酸化反応を繰り返している。
反対に、リン酸化タンパク質からリン酸基を脱離させる酵素をプロテインホスファターゼ(脱リン酸化酵素)と呼ぶ。
この2つの酵素の働きにより、生体内のリン酸化の活性レベルが調節されている。
www3.chubu.ac.jp/faculty/ohnishi_motoko/kenkyunaiyo/kenkyu1/
セリン/スレオニン ホスファターゼ
セリン/スレオニンを基質とするプロテインホスファターゼをSer/Thrホスファターゼと呼ぶ。
Ser/Thrホスファターゼは、その化学的性質によってPP1、PP2A、PP2B、PP4、PP5、PP6、PP7に分類される。
プロテイン ホスファターゼ2A(PP2A)
PP2AはすべてのSer/Thrホスファターゼの中で最も広い特異性を示し、真核細胞に広く一般的に存在する。
PP2Aは主に代謝調節、シグナル伝達、DNA複製・転写、細胞増殖などの広い細胞機能に関与しており、血液がんの腫瘍抑制因子としても同定されている。
PP2Aの機能低下はパーキンソン病、アルツハイマー病と関連しており、治療標的候補のひとつとして浮上している。
アルツハイマー病
タウホスファターゼPP2A
PP2Aは脳のリン酸化を調節する主要なタウホスファターゼ。(71%を占める)
アルツハイマー病の脳ではPP2Aが半減。特定のタウリン酸化酵素が重要な役割を果たす。
www.ncbi.nlm.nih.gov/pubmed/16262633
PP2A阻害因子の増加
核内I1 PP2A(PP2A阻害因子1)および細胞質I2 PP2A(PP2A阻害因子2)の活性は、アルツハイマー病脳において20%増加する。
www.ncbi.nlm.nih.gov/pmc/articles/PMC1602412/
www.ncbi.nlm.nih.gov/pubmed/18245083
PPP2A阻害因子によるタウのリン酸化
核内I1 PP2Aおよび細胞質I2 PP2A活性はタウの過剰リン酸化を誘導。
www.ncbi.nlm.nih.gov/pubmed/18245083
www.sciencedirect.com/science/article/pii/S0014579311005497
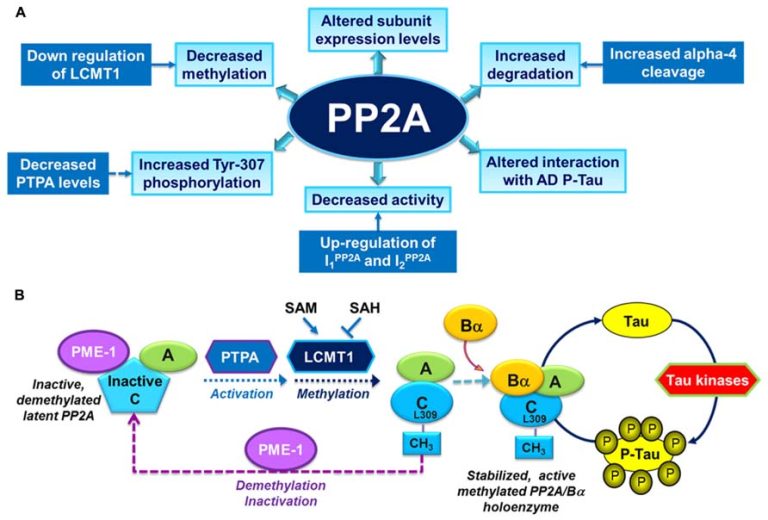
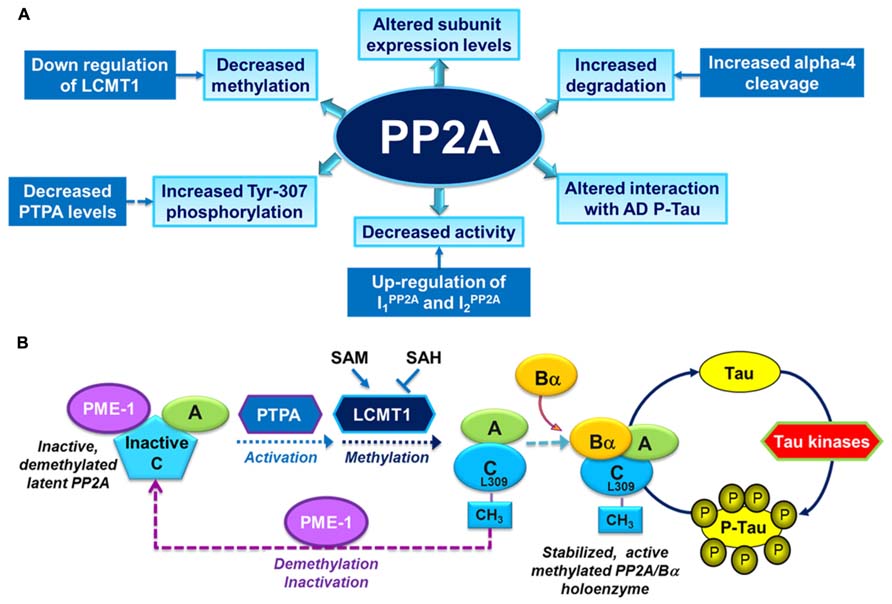
PP2Aの機能不全は、タウの過剰リン酸化、アミロイド形成およびシナプス欠損に関連している。
PP2Aの調節因子およびPP2A触媒活性の変化
サブユニットの発現
メチル化および/またはリン酸化
が、アルツハイマー病患者の脳領域で生じていることが報告されている。
www.ncbi.nlm.nih.gov/pubmed/15099019/
www.ncbi.nlm.nih.gov/pubmed/8395566/
www.ncbi.nlm.nih.gov/pubmed/7616230/
www.ncbi.nlm.nih.gov/pubmed/11259128/
PP2A酵素の変調は、アルツハイマー病に関与するその他の多くのセリン / スレオニンホスファターゼの活性にも影響する。
その他の疾患
PP2A関連経路は、アルツハイマー病だけではなく多くの疾患に関与している。
PP2A活性の喪失は癌、神経変性疾患の両方の病理学的なメカニズムを含みうる。
www.ncbi.nlm.nih.gov/pubmed/29358171
急性骨髄性白血病
www.ncbi.nlm.nih.gov/pubmed/27102578
慢性閉塞性肺疾患(COPD)
www.ncbi.nlm.nih.gov/pubmed/22223484/
心疾患
www.ncbi.nlm.nih.gov/pubmed/25320082/
対照的な役割を持つ糖尿病におけるPP2A
インスリンシグナル伝達が正常に機能している場合にはおいてインスリンの分泌はPP2Aの不活性化をもたらす。グルコースホメオスタシスが機能を果たすには、PP2Aのダウンレギュレーションを必要とする。
しかし、2型糖尿病患者ではインスリン分泌によるPP2Aの減少は見いだせなかった。
www.ncbi.nlm.nih.gov/pubmed/8940115/
www.ncbi.nlm.nih.gov/pubmed/12534451/
PP2Aの構造
www.sciencedirect.com/science/article/pii/S0092867409012549
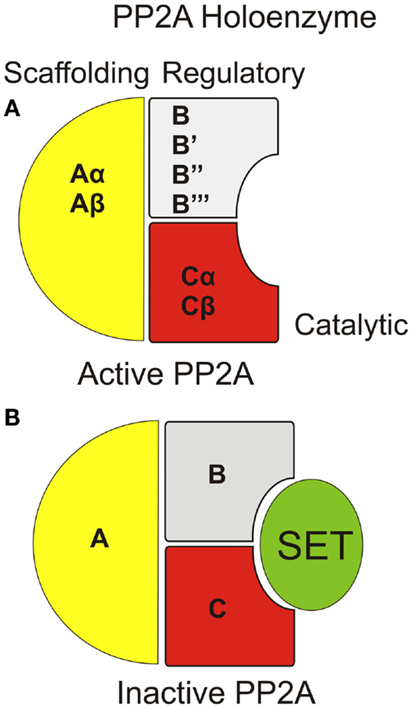
A:足場サブユニット、B:調節サブユニット C:触媒サブユニット
www.frontiersin.org/articles/10.3389/fonc.2014.00388/full
PP2Aを構成する触媒ユニットC末端のメチル化により、ホロ酵素の形成が促進されある。
ホロ酵素が形成されたメチル化PP2Aはタウタンパク質に対して高い活性をもつようになる。
PP2Aはヒトの脳のタウの脱リン酸化を71%を媒介する。

www.ncbi.nlm.nih.gov/pmc/articles/PMC4803620/
PP2Aのメチル化
ロイシンカルボキシルメチルトランスフェラーゼ1(LCMT-1)
PP2Aの脱メチル化
プロテインホスファターゼメチルエステラーゼ1(PME-1)
PP2Aの作用メカニズム
GSK-3βはPP2Aの発現を減少させる
www.ncbi.nlm.nih.gov/pubmed/21554241
www.ncbi.nlm.nih.gov/pubmed/22732552
PP2Aの調節
ハーブ
フォルスコリン
www.ncbi.nlm.nih.gov/pubmed/21098287
www.ncbi.nlm.nih.gov/pubmed/6267587/
www.ncbi.nlm.nih.gov/pubmed/21233840/
www.ncbi.nlm.nih.gov/pubmed/12065707/
www.ncbi.nlm.nih.gov/pubmed/18213449/
EGCG(緑茶カテキン)
www.ncbi.nlm.nih.gov/pubmed/9175270/
カルノシン酸(ローズマリー)
ローズマリーから単離されたカルノシン酸によるPP2Aの調節効果 in vitro
www.ncbi.nlm.nih.gov/pubmed/22453599/
栄養化合物
ビタミンE(α-トコフェロール)
www.ncbi.nlm.nih.gov/pubmed/19420015
www.ncbi.nlm.nih.gov/pubmed/24725826
ベタイン、SAM、ビタミンB
葉酸、B12の低値によるホモシステイン濃度の上昇はPP2Aのメチル化を妨害
journal.frontiersin.org/article/10.3389/fnmol.2014.00016/full
パルミチン酸
パルミチン酸が、セラミド依存性PP2A活性化を介して、内皮細胞におけるAMPKおよび内皮酸化窒素合成酵素のリン酸化を阻害する。
www.ncbi.nlm.nih.gov/pubmed/17255104/
ホルモン
メラトニン
メラトニンは、PP2A負リン酸化を部分的に逆転させ、酸化ストレスのマーカーであるマロンジアルデヒドレベルを低下させた。in vivo
www.ncbi.nlm.nih.gov/pubmed/20542922/
プロゲステロン
www.ncbi.nlm.nih.gov/pubmed/19945993/
PP2A活性 薬剤
メトホルミン
www.ncbi.nlm.nih.gov/pubmed/21098287
メマンチン
メマンチンは、ラットの海馬PP2A活性を増加させ、タウリン酸化を減少させる。
www.ncbi.nlm.nih.gov/pubmed/15147906
www.ncbi.nlm.nih.gov/pmc/articles/PMC4689401/
トログリタゾン(PPARγアゴニスト)
www.ncbi.nlm.nih.gov/pubmed/16825603
www.ncbi.nlm.nih.gov/pubmed/9271121/
1,8-ナフチリジン(コリンエステラーゼ阻害剤)
www.ncbi.nlm.nih.gov/pubmed/20575555/
ジチオレチオン
www.ncbi.nlm.nih.gov/pubmed/19701246/
タウロリジン
www.ncbi.nlm.nih.gov/pubmed/19460788/
4-ヒドロキシノネナール
www.ncbi.nlm.nih.gov/pubmed/12893001/
www.ncbi.nlm.nih.gov/pubmed/12815460/
フェノチアジン
www.ncbi.nlm.nih.gov/pubmed/24401270/